基于特示關系的生態學信息及其認識論意蘊*
2014-12-21 01:40:38范念念
華東師范大學學報(教育科學版) 2014年3期
王 義 范念念
(1.東北大學文法學院科技與社會研究中心,沈陽110819;2.四川大學水力學與山區河流開發保護國家重點實驗室、水利水電學院,成都610065)
美國生態心理學家Gibson 的生態學方法是在實在論的背景中提出來的。實在論和反實在論爭論問題的核心是,被感知的環境是否獨立于感知者,如果獨立,如何來證明它的獨立?顯然,這種哲學爭論實際上已經超出科學實證的界限。因為實證有效的前提是理論只有指向的是客觀事實,實證方法才能有效,而這里由于涉及到客觀事實和對客觀事實的感知,因而付諸于實證來證明實在論是無效的。對于實在論者來說,如果堅持被感知的世界是實在的這個前提,那么就需要說明,實在的世界和被感知的世界如何是同一個世界。然而對于Gibson 來說,實在論的證明是可能的,它就是能量運動構型和實在之間的特示關系。
一、引言:心理學的生態學方法
特示關系蘊含在心理學的生態學方法之中。在論述特示之前,必須首先闡述生態學方法的實際內涵。到目前為止對生態心理學的理解中,國內學者仍然延循著Lombardo 的觀點,即,Gibson 生態心理學的總原理是相互作用(Lombardo,1987),它蘊含著一種整體主義視角。然而,整體主義對還原主義、機械論的批判并不能足夠精確地表達出生態學方法的實質內涵。因為符合“方法”的必要條件是,它必須首先達到定量性的標準,才能具有可操作性。
首先需要知道,即使Gibson 本人也沒有對生態學方法進行單獨論述,它只是貫穿在其對心理學的實際應用中。在The ecological approach to visual perception 一書開篇之處,Gibson 提到“值得牢記卻經常被忽略的事實是:動物和環境這兩個詞是不可分割的一對,一個詞暗示著另一個詞的存在,沒有周圍環境動物無法生存……動物-物體被以一種特殊方式環繞著,一個環境圍繞一個生命體的方式與一堆物體圍繞著一個物理物是不同的”(Gibson,1986,p.8),這可以看作Gibson 對生態學方法最為素樸的表達。
生態學方法分為三個有機研究的維度。一是環境與動物之間的關聯或互補,Gibson 將之命名為可供性(affordance)(Gibson,1986,p.127),在生態學語境中,它就是生態學實體,它所作用的對象是動物的行為而不是身體。二是這種關聯和互補可以特異化(be specific to)為能量的運動構型、形態、結構等等,也就是說,這里的構型由于是關聯和互補關系的特異化(specificity),在生態心理學的語境中是特示(specification),因而能量運動構型對于被能量(而非運動構型)所作用的動物來說就是潛在的可使用、可獲得的(available)信息。第一維度和第二維度合在一起意味著環境中存在著一些獨立于動物身體(生理)特異性的信息,這些信息是一種客觀形成的信息,它在心理學中屬于Gibson 生態心理學的獨創。三,環境中存在著客觀信息,并且它們標識著動物身體和環境之間交互的可能——行為,因而,對它們的拾取(pickup)就是心理學研究或感知覺研究的主要內容(Gibson,1986,p.239)。補充一句,這里信息拾取當然也可以通過認知科學中的“具身認知”來理解。
在Gibson 之后,生態學方法逐漸地融入自然科學之中,相互作用原理也獲得了精確的內涵。Warren 的登臺階知覺判斷實驗最為經典而簡潔地將可供性知覺應用于定量研究中(Warren,1984)。Warren 探討了生態學方法與傳統方法之間的差別,即生態學方法使用內在單位進行測量(intrinsic measurement),并提出了身體尺度(body-scaled)和行為尺度(action-scaled)(Warren,1984,p.686),兩者之間的差別不在本文內容之內,因而不作進一步論述。Warren 的實驗揭示出,當臺階高度除以被試的腿長之后,高和矮兩組實驗者的知覺判斷值出現一致,這說明環境的身體比性質與知覺判斷之間存在內在的關聯,即知覺判斷范疇與行為的臨界值即最高值、最優值存在一致關系。
身體尺度或行為尺度是對Gibson 交互作用原理的定量性的表達。它們說明了,動物直接生存的環境是在尺度交互意義上的物理環境。身體比是環境和身體交互作用而形成的本征或內在(intrinsic)尺度,它可以應用到心理學的研究中,是生態學方法的實質內涵。更詳細地說,可供性是由環境的動力(重量、應力、摩擦力等)性質和動物的動力(身體的重量、身體的結構、四肢)聯結而成,這種交互性質限定了動物行為的界限(行為閾限),展示了動物行為所發生的本征環境——或本征時空。“每個動物——至少在某些程度上——都是感知者和行為者,用老式詞匯說,它是有感知能力和有生命力的。它是環境的感知者,并且在環境中行動。但這并不是說它感知物理世界并在物理的空間和時間中行動。”(Gibson,1986,p.8)
測量尺度和本征尺度之間的差別和內在關聯,是生態學的重要問題,也是Gibson 生態心理學中生態學方法的本質內涵,它關系到人類認識自然的工具、手段和方法,以及因此而決定的認識界限和認識發展的規律,同時也是解決認識論問題的重中之重,但是這一問題在認識論中還不是特別凸顯。“總體來講,尺度的存在根源于地球表層自然界的等級組織和復雜性。尺度本質上是自然界所固有的特征或規律,而為有機體所感知。因而尺度又可分為測量尺度和本征(Intrinsic)尺度。測量尺度是用來測量過程和格局的,是人類的一種感知尺度,隨感知能力的發展而不斷發展。本征尺度是自然現象固有而獨立于人類控制之外的。測量尺度相當于研究手段,隸屬方法論范疇,而本征尺度則是研究的對象。尺度研究的根本目的在于通過適宜的測量尺度來揭示和把握本征尺度中的規律性。”(呂一河,傅伯杰,2001,第2097 頁)
二、特示關系與生態信息
生態學第二個研究維度是信息理論,它的核心概念是特示(specify)。生態系統中充斥著多種形式的能量,如光能、機械能、內能、化學能、原子能、生物能等,它們是物質的固有屬性,由于物質本身的屬性而具有一些確定的量度,是引發物質運動變化的原因。但另一方面,能量所引起的運動變化必然展現為特定的樣式、構型或結構,它們反過來特示(specify 或be specific to,下文簡稱為特示)物質所具有的屬性(Gibson,1960,p.701)。也就是說,能量具有兩種意義,第一是在它們的轉換中遵循量度上的守恒,是物質之間相互作用所具有的量度,第二則作為信息的載體,在這個意義上,它并不關系到物理運動形式上的變化和轉換。按Chemero 所說,“信息不是環境中物體或物質以外某種可測量、可計量的東西,相反,信息是環境的某種關系特征。……光中的信息就是光和環境之間的關系。”(Chemero,2009,p.108)
因而,從對特示關系的分析中,可以發現,能量運動構型和物質屬性之間的關聯,從屬于動力學研究范疇,也就是說,信息是動力學的,它的展示是動態的,它與實在之間的關聯是無法通過測量尺度還原的,關于這一點,Gibson 以后,Runeson 首先明確指出生態學信息的動力學機制(Runeson,1983,p.587-588)。信息載體的運動既然是動力學的,那么就意味著,信息是物理的、客觀的、實在的,“如果某機體接受器中能量流的不變量存在,如果這些不變量對應于環境的不變特性,并且如果它們是機體對環境知覺的基礎,而不是感覺信號的基礎,那么,認識論中的實在論就有新的證據,同樣,心理學中也會出現新的知覺理論”(Gibson,1982,p.374);同時也說明,信息的存在獨立于知覺者,或者說它是非感受器和神經特異性的,“信息這個詞語在本書中的意義(不是在其他的書中)是觀察者的環境的呈現,而不是觀察者的接收器或感覺器官的呈現。物體的性質通過信息展現出來;接收器和神經的性質通過感覺展現出來。關于世界的信息與感覺的性質沒有什么關系。”(Gibson,1986,p.242)
特異性(specificity)是一個神經生理學概念,它說的是,感覺、神經興奮和能量刺激作用之間的關系。“在將外部世界的信息傳給個體的過程中,感覺系統執行一系列普遍的功能。其中最基礎的是,每個系統都與某個刺激的特異性(specificity)發生反應,它們使用特異化的細胞——外周接收器(peripheral receptors)——把刺激轉換為所有神經元都能夠使用的信號。”(Squire,Berg,& Bloom,etc.,2009,第535 頁)在認知科學中,神經特異性概念穩固地樹立了外部世界的“表征”對于認知研究的基礎性作用。比如說,認知科學家們將認知劃分為感受和解釋這兩個機制。例如他們認為,“感覺是對刺激的探測,并識別出某件事情的發生;而知覺則是解釋和理解。”(Squire,Berg,& Bloom,etc.,2009,第536頁)
從這兩個特異性關系的比較,可以發現,兩者在測量方法上存在差異。生態學方法強調身體尺度或行為尺度對于動物行為研究的基礎性作用,因而不變量并不是非量化的東西。在Gibson 以后,生態信息理論開始走向量化的發展方向。以生態光學為例,Gibson 認為生態光學的核心假設是光列陣的變量能夠承載關于作為光來源的環境的信息。這意味著,光列陣不是復制實在,而是特示實在。Gibson的學生Lee 致力于將基于生態光學的視覺理論數學化,將其應用在位移行為的視覺控制研究上。在《光流域:視覺的基礎》一文中,Lee 提出光流域中包含有光學變量τ(t),它能夠表示為在某個時刻t,距離與速度之間的比率,即,Z(t)/V(t)。在這里,光學變量τ(t)是一種光學變化率(即擴張率),它能夠作為動物為保持與表面特定關系的光學信息量(Lee,1980)。以駕駛行為為例,在時刻t,某輛瞬時速度為V(t)的汽車駛向一個與其距離為Z(t)的表面,為了防止沖撞到這個表面上,駕駛員必須減速(D)剎車,使得該輛汽車在無限接近表面的時候停止,因而,根據位移公式,V(t)2/2D≤Z(t)。通過對Z(t)/V(t)=τ(t)公式進行微分,得到Z(t)D/ V(t)2 =1-dτ(t)/dt,最后得到公式:
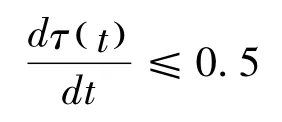
按照這個公式,在與表面接近的過程中,動物與表面之間的接觸時間(time-to-contact)就包含在光的運動變量中,它提供了如何與表面保持特定關系的信息,例如,如若避免被碰撞,就需要將τ(t)的時間導數保持為小于等于0.5。對于這個公式的驗證,請參看Lee 對蜂鳥飛行的研究(Lee,1991)。光學τ(t)理論提出之后,它被拓展到接觸時間的聲音變量研究中(Shaw,McGowan,& Turvey,1991)。
三、特示的形式化及生態信息理論的發展
如果能量運動構型是物理事實的特示或特異化,那么就意味著,環繞著動物的能量環境就必然特示動物身體尺度或行為尺度的相關信息,這種信息本質上是行為暗示的,而且它們是準確的清晰的。但另一方面,Gibson 也提出了能量樣式對事實特示的局限,這種局限產生了模糊信息。例如光不能特示透明物的結構,前庭系統中耳石的運動樣式能夠特示啟動、停止、推拉、主動和被動加速,即能夠特示不穩定狀態,但不能夠特示靜止不動和直線勻速運動狀態之間的區別,而半規管的運動樣式也不能夠特示靜止狀態和勻速旋轉之間的區別,等等。更為明確地說,所謂特示,即能量列陣的樣式和事實之間的專屬指向關系,但在某些臨界條件下會出現一對多,或多對一的關系,這時就產生了模糊信息。模糊信息源于任何能量列陣的運動構型并沒有表現出足夠的異質性,從而導致動物在拾取信息的時候無法判斷事實。
Stoffregen 等人在姿態平衡的知覺研究領域,對特示關系理論進行了完善,在生態信息理論的發展中作用巨大。就身體姿態的平衡來說,首先需要考慮到身體所處環境的性質,即使在靜止的狀態下身體也存在振蕩性的擺動,因而它是一個動力系統,這意味著動物在感知身體方向的時候,面對的是一個動態的信息環境;其次,對于動態信息環境的感知涉入到自身的動力學性質,因而,這里就涉及非線性動力感知,具體地說,在判斷方向的時候,不同的感知系統、感知系統與運動系統、身體系統與環境之間的耦合,方向知覺與這種嵌套系統之間的耦合關系是一種復雜的非線性知覺。
因而,Stoffregen 和Riccio 發現了姿態平衡系統所存在的知覺復雜性問題,對基于前庭系統掌管重力感知的定向理論進行了批評(Stoffregen & Riccio,1988)。根據經典的定向理論,身體的方位信息主要基于重力慣性向量,即公式為,Fs=Fg +Fa。也就是說,當身體方向與重力方向發生偏離的時候,身體會跌落,這時候重力的方向就為身體方向提供了基準。然而,有一些情況會引發重力錯覺,即,當視覺、聲音、觸覺等慣性參照系與重力方向產生沖突的時候,會引發重力錯覺;當身體主動運動產生加速度的時候,其慣性力向量則與重力產生偏離,前庭系統也不能區分慣性力和重力。這時候的方向判斷就需要在前庭系統感知基礎上,進行歸納推理,即與其他方向信息對比處理,完成正確的方向判斷。
在Stoffregen 和Riccio 看來,根據生態學法,方向知覺必須考慮到支撐面的性質,因為它影響到支撐面對重力的抵抗以及身體為了保持某種姿態所必須對該支撐面所主動施加的作用。在水中,水的浮力分布全身而抵消了質量中心和支點之間的力臂,身體不會因為和重力方向的偏離而產生加速度,因而前庭系統無法判斷垂直方向;在徹底失重情況下,就沒有重力慣性向量,因而,也就不需要保持特定的姿勢了;在有支撐面的情況下,如果支撐面和重力方向產生偏斜角度,質量中心和身體和支撐面之間的支點(即腳)就會產生旋轉運動,而為了控制身姿保持平衡,動物必須對支撐面主動施以驅力來抵抗這種重力慣性向量,這時候,就會產生出一種不斷向前的加速度,即身體穩定的方向開始與重力方向發生偏移。
Stoffregen 和Riccio 認為,這種不穩定的平衡狀態才是運動中身體垂直方向的信息來源。“定向行為涉及到對一種特定運動學狀態的獲得,即,不穩定的平衡(unstable equilibrium)。因此我們相信,這種不穩定的平衡方向才是相關于姿勢控制的垂直狀態”(Stoffregen & Riccio,1988,p.8)。也就是說,身體方位的信息既包含重力慣性向量,也需要考慮到身體的運動狀態以及考慮到支撐面和身體之間的關系。這種信息就是“模間不變量(intermodal invariants)”信息,它需要知覺系統的統籌才能獲得,即,它不是專屬地指向特定模態的感受器的信息(例如前庭系統對垂直方向的感知),然后經神經系統對其他模態感受器接受的信息進行整合處理的產物,它的形成完全是一個身體與環境相互作用的事件,即一個生態學事件。
繼提出“模間不變量”概念之后,Stoffregen 和Bardy(2001,p.199)又對特示類型進行了邏輯性地分類。他們認為,從能量列陣和物理事實之間的對應關系上來看,包含有3 種類型的特示:1.“無特示(No Specification)”;2.“單模態特示(Modal Specification)”;3.“獨立特示(Independent Specification)”,包括“多模態特示(Multiple Specification)”和“變模態特示(Amodal Specification)”。
“無特示”,即能量列陣和實在之間的對應關系是多對多,即,對于某個確定的能量列陣,也可以對應多個實在;對于確定的實在,可以對應某能量列陣的不同形式、結構。在無特示關系中,感覺是模糊的,感覺本身不能作為信息,只能作為信息處理的材料;“單模態特示”關系則主要是Gibson 生態學方法中的特示觀點,即單個的能量列陣和實在之間是一一對應關系,例如光環境和光列陣之間的確定對應關系,振動事件和聲波之間的對應關系,等等;“獨立特示”主張,實在和能量列陣之間的對應關系是一對多,即實在引發相互獨立的多種模態列陣以及能量列陣的不同方面。其中的“多樣特示”主張,實在某方面影響某特定列陣的多重結構,這些多重結構之間互不相關地特示實在的變化。
Stoffregen 和Bardy 認為,在動物-環境的相互關系中,只有多種模態能量列陣之間生成的高階列陣即“全模態列陣”才能夠清晰地指向、特示一個確定的事件(Stoffregen & Bardy,2001,p.206)。“全模態列陣”揭示出生態系統中能量之間的相互關系,以及這種相互關系對生態事件的特示。任何一事件的變化,都會影響到多種列陣的樣式變化,這種惟一的指向關系要求在實在和運動構型之間只能存在惟一的運動構型,因而,無論是單模態特示,還是獨立特示,都沒有考慮到實在特示的唯一性,因而存在邏輯上的欠缺。全模態列陣不是專屬于某特定模態感知系統的信息,而是專屬于知覺系統的信息,它的提出是為了規避某特定知覺系統得到信息以后再與其他系統信息進行處理的內在機制問題;它也能夠更好地說明為什么有機體要主動行為來探索信息,為什么行為能夠更好地支持知覺的實施。例如,被試在判斷物體是否在接觸范圍內的時候,在實驗室控制頭部靜止條件下判斷的準確性,和在頭部自主運動條件下判斷的準確性之間存在差異,后者的準確性具有統計學效度(Mantel,Stoffregen,&Bardy,2008)。
四、特示關系的認識論意蘊
Gibson 的生態心理學被視為具身認知的一個理論來源。本文的研究對象則是為具身認知奠定基礎的生態學信息,它是具身認知研究的前奏。理由如下:具身認知的理論前提是實在論的,而不能是表征主義的,即,認識對象必須是實在的、客觀的,或者說,認識論的前提必須是“物理主義”的,原因是,如果認識的對象是“心理的”,那么認識本身就是無意義的語義循環。然而,物理主義認識論的危險性在于,在認識一種具有“主觀的”、“價值的”屬性的研究對象的時候,測量尺度是無法派上用場的,它必須取消“心理的”所標識的屬性。最后像行為主義一樣,將所有的行為都視為是機械的過程。
因而,在這個意義上,Gibson 的生態學方法給認識論指出了一條有價值的方向。其中的特示關系說明,環境中存在著準確的信息,如果將它應用到動物和環境的交互系統研究中,就會發現,環境中存在著的信息并不是刺激式的,也不是解釋的,而是身體尺度或行為尺度的。這說明,信息對于認識的作用是行為引導性的,感覺系統與運動系統的特異性是自然演化的,起源于適應環境選擇的功能性要求,而不是結構性的或機制性的。更進一步說,特示關系將知覺、經驗等內在意向性的認識論研究方法轉變為外在的信息探索問題,在本征尺度的時空中來研究知覺,因而具有將心理學引入動力學研究的可能。但Gibson 沒有進一步將其形式化和數學化。因而在他之后,這些問題成為生態心理學的奠基性工作。由于特示理論是生態信息理論的根基,因而Stoffregen 等人“全模態列陣”的研究進一步豐富了特示關系的內涵和外延。
Squire,L.,Berg D.,& Bloom,F.,etc.(2009).基礎神經學系列3:感覺和運動系統.北京:科學出版社.
呂一河,傅伯杰.(2001).生態學中的尺度及尺度轉換方法.生態學報,21(12).
Chemero,A.(2009).Radical embodied cognitive science.Cambridge,MA:the Massachusetts Institute of Technology Press.
Gibson,J.J.(1960).The Concetp of The Stimulus in Psychology.The American Psychologist.
Gibson,J.J.(1982).New Reasons for Realism.in E.Reed & R.Jones(eds),Reasons for Realism:Selected Essays of James J.Gibson.Hillsdale,New Jersey:Lawrence Erlbaum Associates,Publishers.
Gibson,J.J.(1986).The ecological approach to visual perception.Hillsdale,New Jersey:Lawrence Erlbaum Associates.
Lee,D.N.(1980).The Optic Flow Field:The Foundation of Vision.Phil.Trans.R.Soc.Lond.B290(1038).
Lee,D.N.,Reddish,P.E.,& Rand,D.T.(1991).Aerial Docking by Hummingbirds.Naturwissenschaften 78.
Lombardo,T.J.(1987).The reciprocity of perceiver and the environment.Mahwah,NJ:Lawrence Erlbaum Associates,Inc.P3.
Mantel,B.,Stoffregen,Thomas A.,& Bardy,B.G.(2008).Multisensory perception of whether an object is within reach.Journal of Vision 5.
Runeson,S.,Frykholm,G.(1983).Kinematic Specification of Dynamics as an Informational Basis for Person-and-Action Perception:Expectation,Gender Recognition,and Deceptive Intention.Journal of Experimental Psychology:General,112(5).
Shaw,B.K.,McGowan,R.S.,& Turvey,M.T.(1991).An Acoustic Variable Specifying Time-to-Contact.Ecological Psychology,3(3).
Stoffregen,T.A.,Riccio,G.E.(1988).An Ecological Theory of Orientation and the Vestibular System.Psychological Reviewo,95(1).
Stoffregen,T.A.,Bardy,B.G.(2001).On specification and the senses.Behavioral and Brain Sciences,24(2).
Warren,W.H.(1984).Perceiving affordances:visual guidance of stair climbing.Journal of experimental psychology:Human perception and performance,10(5).
猜你喜歡
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
中國生殖健康(2020年6期)2020-02-01 06:28:50
中國生殖健康(2019年11期)2019-01-07 01:28:02
中華手工(2017年2期)2017-06-06 23:00:31
湖北經濟學院學報·人文社科版(2015年8期)2015-12-29 05:53:07
上海電機學院學報(2015年4期)2015-02-28 14:30:00
中外會展(2014年4期)2014-11-27 07:46:46
計算物理(2014年2期)2014-03-11 17:01:39
外語學刊(2010年2期)2010-01-22 03:31:03
