小鼠性激素結合球蛋白條件基因打靶載體的構建
2015-01-02 06:36:32
中國醫科大學學報 2015年3期
關鍵詞:小鼠
(1.中國醫科大學附屬盛京醫院婦產科,沈陽 110004;2.上海南方模式生物研究中心,上海 201203)
小鼠性激素結合球蛋白條件基因打靶載體的構建
王越1,金鎮1,孫磊1,鄭晉華2
(1.中國醫科大學附屬盛京醫院婦產科,沈陽 110004;2.上海南方模式生物研究中心,上海 201203)
目的構建小鼠性激素結合球蛋白(Shbg)條件基因打靶載體。方法運用ET克隆的方法構建Shbg基因第4-7號外顯子兩側插入loxp位點的條件性剔除載體。首先以BAC DNA為模板克隆出Shbg及其上、下游基因并打靶質粒載體(pBR322-MKAB),利用該質粒和BAC克隆,通過同源重組方式,獲得套取質粒(pBR322-Shbg-Re);再分別以PL452及PL451為模板擴增出含Neo片段,經過再一輪的Red/ET重組成功實現條件性基因敲除打靶載體(pBR322-MK-SHBG-cko)的構建。結果經多個限制性內切酶酶切鑒定和測序證實,構建的pBR322-MK-AB、pBR322-Shbg-Re、SHBG-Ln重組質粒和Shbg條件基因打靶載體(pBR322-MK-SHBG-cko)結構正確,與設計相符。結論成功構建了小鼠Shbg條件基因打靶載體,為后續Shbg基因條件敲除小鼠模型的構建奠定了基礎。
性激素結合球蛋白;條件性基因敲除;載體;Red/ET重組
性激素結合球蛋白(sex hormone-binding globulin,SHBG)是一種能結合性激素甾體的球蛋白,主要由肝臟組織細胞合成和分泌,在其他組織中如睪丸、腦、卵巢、胎盤、子宮內膜和乳腺癌組織也有SHBG基因表達[1]。妊娠期糖尿病的發病機制與2型糖尿病相似,妊娠期胰島素抵抗的增加和胰島β細胞分泌的減少也是其發病的病理生理基礎。目前認為SHBG的低水平是2型糖尿病發生的獨立危險因素,是高胰島素血癥性胰島素抵抗的一個標志[2]。最近研究發現,SHBG基因啟動子(TAAAA)_n重復多態與糖代謝異常有關,這種重復多態性可以影響血液循環中的SHBG水平,對易發生糖尿病等并發癥的高危個體的早期識別具有重要的價值[3,4]。我們通過RNA干擾與基因沉默技術研究了胎盤滋養層合體細胞中SHBG的自分泌作用機制;通過胎盤滋養層合體細胞胰島素抵抗模型的建立,研究了SHBG對其胰島素信號的轉導和胰島素抵抗的影響[5~7]。
目前利用基因敲除技術觀察候選基因在產生胰島素抵抗和胰島素分泌缺陷中的作用已成為該領域的研究熱點[8],國際上尚無SHBG基因敲除動物模型的研究。Red/ET重組(Red/ET recombination)技術因不涉及酶切、連接反應,具有高效率,且不會引入突變,為構建打靶載體提供了一種新的可靠的方法[9]。為此我們采用優化的Red/ET同源重組技術,構建Shbg條件性基因敲除載體,為進一步獲得正確同源重組的胚胎干細胞(embryonic stem cells,ESC)和建立Shbg條件基因敲除小鼠模型奠定實驗基礎。
1 材料與方法
1.1 材料
1.1.1 質粒與菌株:含Shbg的BAC宿主菌購自BAC PAC資源中心(Children′s Hospital akland Research Institute)。質粒PL-452、菌株EL-350由Liu Pengtao(英國劍橋Wellcome Trust S-anger Institute)惠贈。含Red/ET重組酶基因的pSC101-BAD-γβα-A -tet質粒由張友明(德國Gene Bridges GmbH)惠贈。大腸桿菌D-H5α株為博大泰克產品。質粒pBR322-MK為上海南方模式生物研究中心改建。
1.1.2 酶及主要試劑:各種限制性內切酶、T4 DNA連接酶及PCR相關試劑購自TaKaRa公司。L-阿拉伯糖、氨芐青霉素、氯霉素、卡那霉素、四環素等常規化學試劑主要購自Sigma和上海化學試劑公司。DNA片段回收試劑盒購自華舜生物公司。BAC質粒DNA提取試劑盒購于Qiagen公司。其他試劑均為國產分析純。所有PCR引物合成和測序均由上海生工生物技術服務有限公司完成。
1.1.3 引物:由上海生工生物技術公司合成,打靶載體構建用引物:SHBG-A1CCCAAGCTTGTGGGTTGCTATGTGGGTTCCG;SHBG-A2 GCAAATGGTAC CCTGGAGAGAGGGCTCAGTGGTT;SHBG-B1 TAA ACAGGTACCAGGAGTGAGTGGACCGCTGT;SHBGB2 CGCGGATCCGTGGGAGGCAATTCGAGGTCTT;SHBG-L1 CTGAGGGGAACGAGATCACTGGGAGGA AAGAGAGGGGTGACTTGTGGGGTGTTGCTCAAGC ACTGGTCGACGAATTCCTGCAGCCCAATTC;SHBG -L2 ACAAGCCTGCCCCCTTTCTATTCCACTTTTATT TTCTAGTTATAAAACCACAGGCCTCAGGCACTGGA TCCCCTCGAGGGACCTA;SHBG-F1 TGGGAAGTG GCTATTCTTGTCCTAGTTTCATCAGATGGGAATC
CCAAGGCTGAGTCAGTATGTCAGAATTCCGAAGTT CCTATTC;SHBG-F2 AGCCGAGTACAGCTGAAA GAAGATGCTCTGGAACATGGGTGCTTGGCTCACTC AGACTGTGTCACGGATCCACCTAATAACTTCG。
1.2 設計
1.2.1 Shbg基因的生物信息學分析及同源臂的設計:通過分析小鼠Shbg基因的全長外顯子起始密碼子以及全基因的長度及編碼蛋白情況,綜合考慮決定要進行敲除的基因片段和設計同源臂。
1.2.2 同源臂套取擴增:
1.2.2.1 上下游同源臂A、B的擴增 在PCR引物末端設計好酶切位點,以含Shbg的BAC DNA為模板,PCR擴增Shbg基因外上游基因片段A同源臂、Shbg基因外下游基因片段B同源臂。PCR引物為:A臂:SHBG-A1和SHBG-A2,長度為372 bp;B臂:SHBGB1和SHBG-B2,長度為398 bp。常規PCR擴增,經1.25%瓊脂糖凝膠電泳后,回收獲得372 bp/398 bp的目的基因片段。
1.2.2.2 含neo同源臂片段的擴增 同樣方法,以PL452質粒為模板DNA,SHBG-L1,SHBG-L2為引物,PCR直接擴增出含Neo的L片段(即L1-neo-L2片段)。以PL451質粒為模板DNA,SHBG-F1,SHBGF2為引物,PCR直接擴增出含Neo的F片段(即F1-neo-F2片段)。
1.2.3 感受態細胞的制備:
1.2.3.1 大腸桿菌DH5α感受態細胞制備 將菌液轉入50 mL離心管中,冰上放置10 min。4℃,4 000 r/min離心10 min。棄去上清,將管倒置1 min使培養液流盡,用冰上預冷的0.1 mol/L的CaCl2溶液10 mL輕輕懸浮細胞,冰上放置30 min。0~4℃4 000 r/min離心10 min,棄去上清,加入2 mL預冷的0.1 mol/L的CaCl2溶液,輕輕懸浮細胞,冰上放置,即可獲得DH5α感受態細胞。
1.2.3.2 EL350感受態細胞制備 挑選一EL350克隆,32℃培養過夜,次日轉接到3 mL的新鮮LB培養液,32℃培養,加10%阿拉伯糖后32℃繼續培養,制備感受態細胞。
1.2.3.3 BAC感受態細胞 制備誘導重組酶表達的大腸桿菌電轉感受態細胞,將BAC單克隆常規轉化至其中,通過篩選獲得轉入成功的BAC細胞。將獲得的細胞在含氨芐青霉素(50 μg/mL)LB板中32℃培養過夜,次日轉接到3 mL的新鮮LB培養液,32℃培養后迅速移至42℃熱激,迅速冰上冷卻,制備感受態細胞。
1.3 方法
1.3.1 同源臂的擴增:以BAC DNA為模板擴增出所需的同源臂A、B,PCR體系為:模板DNA(BAC質粒/菌液)2 μL,10×PCR緩沖液5 μL,dNTP 4 μL,上下游引物各2 μL,rTaq酶1U,DDH2O 36 μL。PCR循環條件:95℃預變性4 min,94℃變性30 s、58℃退火30 s、72℃延伸30 s(共30個循環)、72℃延伸5 min。同樣方法,以PL452質粒為模板DNA擴增出含Neo的L片段(即L1-neo-L2片段)。以PL451質粒為模板DNA擴增出含Neo的F片段(即F1-neo-F2片段)。PCR體系為:模板DNA(PL452/PL451質粒)2 μL,10×PCR緩沖液5 μL,dNTP 8 μL,上下游引物各2 μL,rTaq酶:1 U,DDH2O 32 μL。PCR循環條件:95℃預變性4 min,94℃變性30 s、58℃退火30 s、72℃延伸2 min(共35個循環)、72℃延伸5 min。
1.3.2 套取質粒的構建:A臂(372 kb)經HindⅢ、KpnⅠ雙酶切后連入經HindⅢ、KpnⅠ雙酶切后的線性化的pBR322-MK載體中,經1%瓊脂糖電泳后,膠回收,在16℃連接過夜,連接物常規轉化大腸桿菌DH5α感受態細胞,陽性克隆采用HindⅢ、KpnⅠ限制性內切酶雙酶切鑒定,獲得的質粒命名為pBR322-MK-A。按照如上操作,再次將B臂(398 kb)經KpnⅠ、BamHⅠ雙酶切連入KpnⅠ、BamHⅠ雙酶切后的線性化的pBR322-MK-A載體中,常規轉化至大腸桿菌DH5α感受態細胞,通過篩選獲得的質粒命名為pBR322-MK-AB。
1.3.3 轉化pSC101-BAD-γβα-A-tet:挑一BAC單克隆,37℃培養過夜,次日轉接到3 mL的新鮮LB培養液,37℃培養,制備感受態,并將pSC101-BAD-γβα-A-tet(ZET-1)轉化該感受態細胞,涂至含氯霉素(20 μg/mL)+四環素(30 μg/mL)LB板,30℃培養過夜。
1.3.4 DNA套取、鑒定:挑取已轉ZET-1成功的細胞,30℃培養過夜,次日以1∶50轉接到3 mL的新鮮LB培養液,30℃培養(OD600<0.2),加L-阿拉伯糖后改至37℃培養,此期間redγ/redβ/redα被誘導表達,經誘導后制備感受態細胞,方法同上。轉化50~100 ng的線性化的pBR322-2S-AB,涂布氨芐板,37℃培養過夜。抽提轉化的質粒,獲得pBR322-SHBG-Re質粒初步酶切鑒定后,進一步用PCR方法測序鑒定重組質粒。
1.3.5 neo基因第1次靶向插入及篩選:將上述獲得的pBR322-SHBG-Re質粒轉化到已制備成功的EL350感受態細胞,涂至含氨芐青霉素(50 μg/mL)LB板,32℃培養過夜。將L1-neo-L2片段(200~300 ng)轉化到BAC感受態細胞,涂至含氨芐青霉素(50 μg/mL)+卡那霉素(15 μg/mL)LB板,32℃過夜。利用介于兩端同源序列之間的線性化位點進行線性化的套取載體在EL350中與BAC發生同源重組,得到SHBG-Ln質粒,并用限制性內切酶及測序鑒定。轉化陽性SHBG-Ln質粒到已制備成功的EL350感受態細胞,涂至含氨芐青霉素(50 μg/mL)LB板[卡那霉素(15 μg/mL)LB板做對照],32℃培養過夜,獲得SHBG-Ln-Cre質粒。
1.3.6 neo基因第2次插入:將SHBG-Cre質粒轉化到已制備成功的EL350感受態細胞,同理轉化F1-neo-F2片段(200~300 ng),涂至含氨芐青霉素(50 μg/mL)+卡那霉素(15 μg/mL)LB板,32℃過夜。即可將上述SHBG-Ln-Cre質粒與含Neo的F片段共同轉入EL350菌株,經過同源重組完成第2次插入neo基因,構建載體pBR322-MK-SHBG-cko。
1.3.7 純化及打靶載體的鑒定:此時得到的質粒往往混有2種質粒(pBR322-MK-SHBG-cko+SHBG-Ln-Cre質粒),將酶切鑒定正確的neo插入質粒重新轉化DH5α感受態細胞,涂至含氨芐青霉素(50 μg/mL)LB板或卡那霉素(15 μg/mL)LB板,此過程套取質粒被去除,最后獲得pBR322-MK-SHBG-cko載體質粒,酶切鑒定并送測序。
2 結果
2.1 Shbg條件性基因敲除打靶載體同源臂的設計
從Ensembl數據庫中(http://www.ensembl.org/ index.html)獲得Shbg基因組序列,根據基因組序列設計打靶載體。小鼠Shbg位于染色體11:69,614,804-69,617,905(-),產生兩個轉錄本Shbg-001和Shbg-002。Shbg-001由8個外顯子組成,蛋白質翻譯起始于第1外顯子,結束于最后外顯子,編碼蛋白大小為403aa。Shbg-002由4個外顯子組成,該轉錄本不編碼蛋白。根據上述信息和基因條件剔除的一般原則,需要選擇剔除核苷酸數不為3整除的外顯子,剔除外顯子4~7后,此段外顯子缺失后使后繼序列發生移碼突變,提早出現終止密碼子,使其蛋白翻譯提前終止,使Shbg重要功能域缺失。運用ET克隆的方法構建Shbg基因第4~7號外顯子兩側插入loxp位點的條件性剔除載體,步驟如圖1。
2.2 同源臂套取擴增及套取
2.2.1 Shbg同源臂擴增:設計好酶切位點,以含SHBG的BAC DNA為模板,擴增其上下游基因,經PCR擴增后獲得長約372 bp/398 bp的DNA片段,分別命名為同源臂A、同源臂B(圖2)。
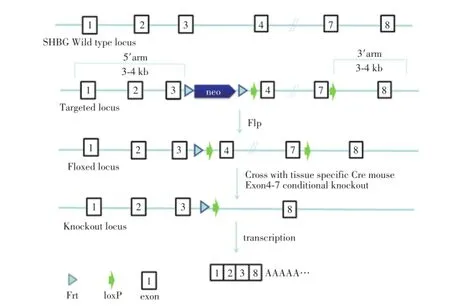
圖1 Shbg基因敲除打靶載體設計Fig.1 Diagram of the strategy to generate Shbg targeting vector
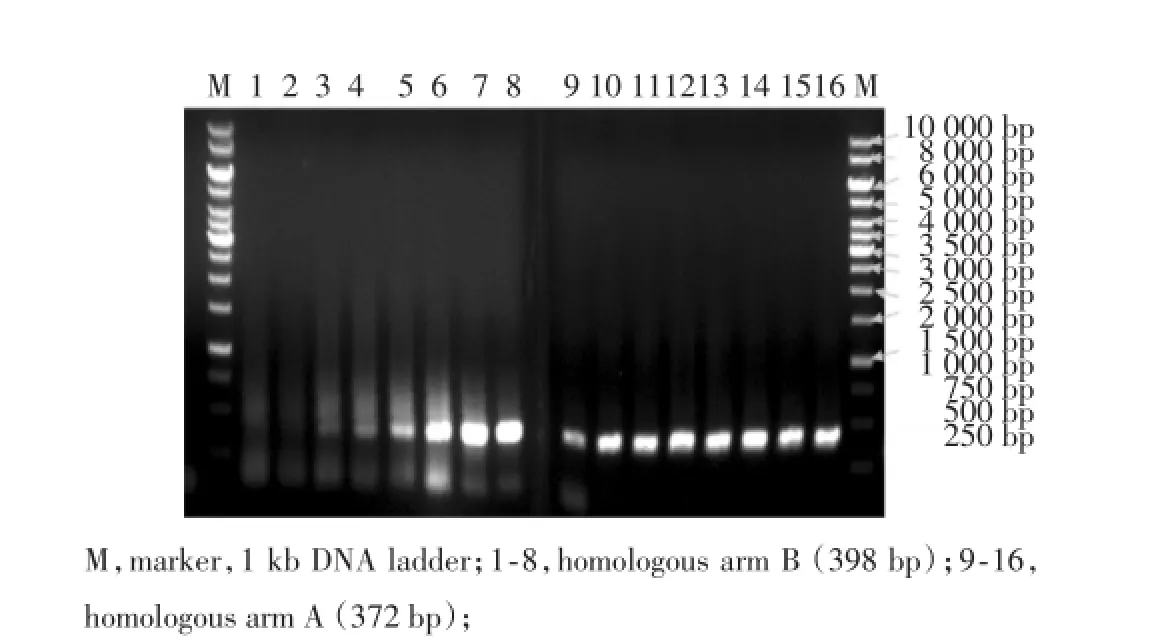
圖2 同源臂A、同源臂B的PCR擴增產物電泳結果Fig.2 Electrophoresis of PCR products of homologous arm A and homologous arm B
2.2.2 含neo同源臂片段的擴增:設計好酶切位點,以含PL452為模板,以SHBG-L1,SHBG-L2為引物,擴增其上下游基因,經PCR擴增后獲得長約2 kb的L1-neo-L2片段,并含neo、LoxP片段和SalⅠ酶切位點(圖3)。同上述方法,以含PL451為模板,擴增出F1-neo-F2片段(圖4)。
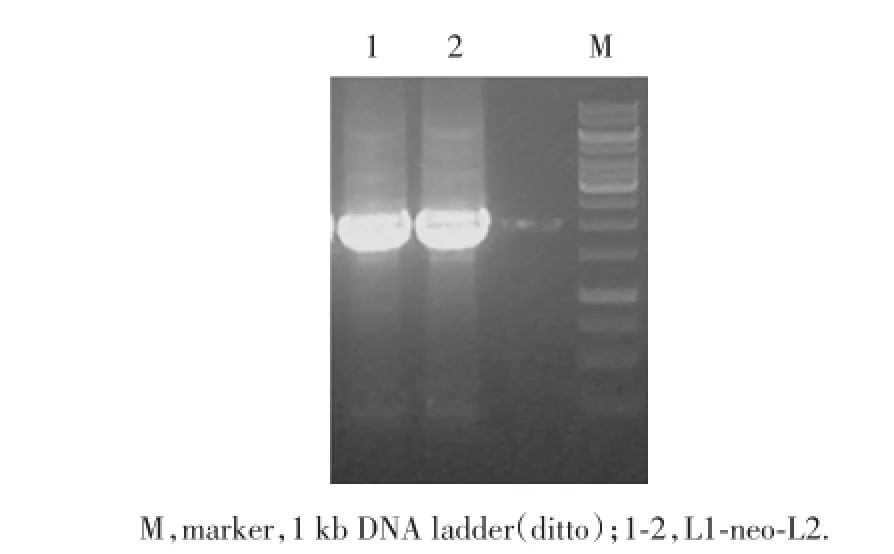
圖3 L1-neo-L2的PCR擴增產物電泳結果Fig.3 Electrophoresis of PCR products of L1-neo-L2
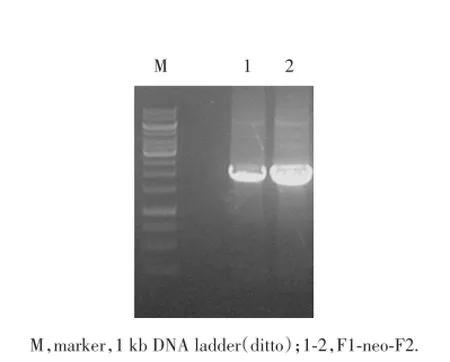
圖4 F1-neo-F2的PCR擴增產物電泳結果Fig.4 Electrophoresis of PCR products of F1-neo-F2
2.2.3 套取質粒的構建:在感受態細胞中,利用HindⅢ&KpnⅠ雙酶切限制性內切酶將A臂克隆至pB-R322-2S質粒中,構成pBR322-MK-A;同種方法,通過KpnⅠ&BamHⅠ雙酶切將下游基因片段B臂連接到pBR322-MK-A,構成pBR322-MK-AB,通過HindⅢ&BamHⅠ雙酶切電泳鑒定(圖5),得到770 bp片段。
2.3 DNA套取、鑒定
挑取已轉ZET-1的BAC單克隆制備感受態細胞,轉化50~100 ng的線性化的pBR322-MK-AB,涂布氨芐板,37℃培養過夜。抽提轉化的質粒,獲得pBR322-SHBG-Re質粒經SalⅠ酶切,得到兩個片段,分別為5 668 bp和7 553 bp(質粒抽提可能會存在RNA沒有被完全降解的情況,不影響質粒酶切),見圖6。
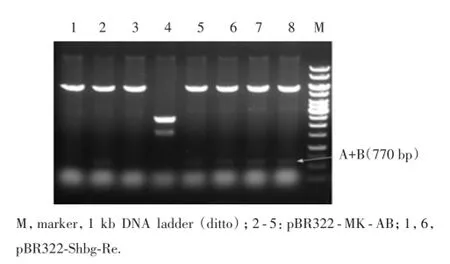
圖5 pBR322-SHBG-Re酶切的PCR擴增產物電泳結果Fig.5 Electrophoresis of PCR products of enzyme digestion of pBR322-SHBG-Re
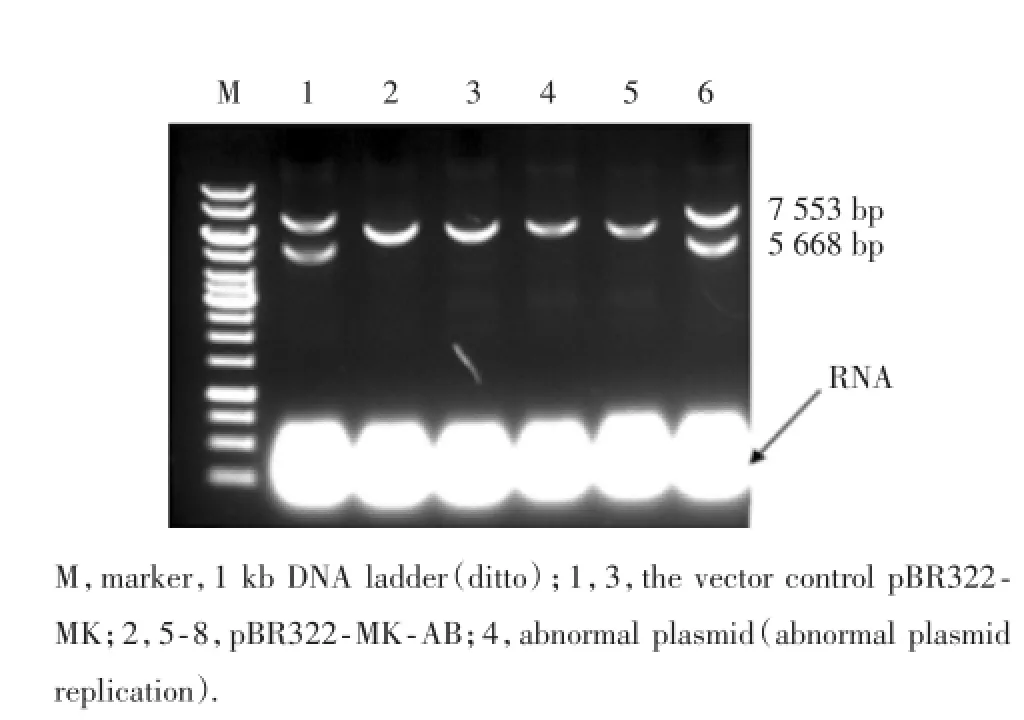
圖6 pBR322-MK-AB酶切的PCR擴增產物電泳結果Fig.6 Electrophoresis of PCR products of enzyme digestion of pBR322-MK-AB
2.4 neo基因第1次靶向插入及篩選
將pBR322-SHBG-Re質粒轉化到已制備成功的EL350感受態細胞,與BAC經同源重組后,得到SHBG-Ln質粒,用SalⅠ酶切,得到4個片段(圖7),分別為7 553 bp、5 668 bp、4 866 bp和2 703 bp。
2.5 pBR322-MK-SHBG-cko測序結果
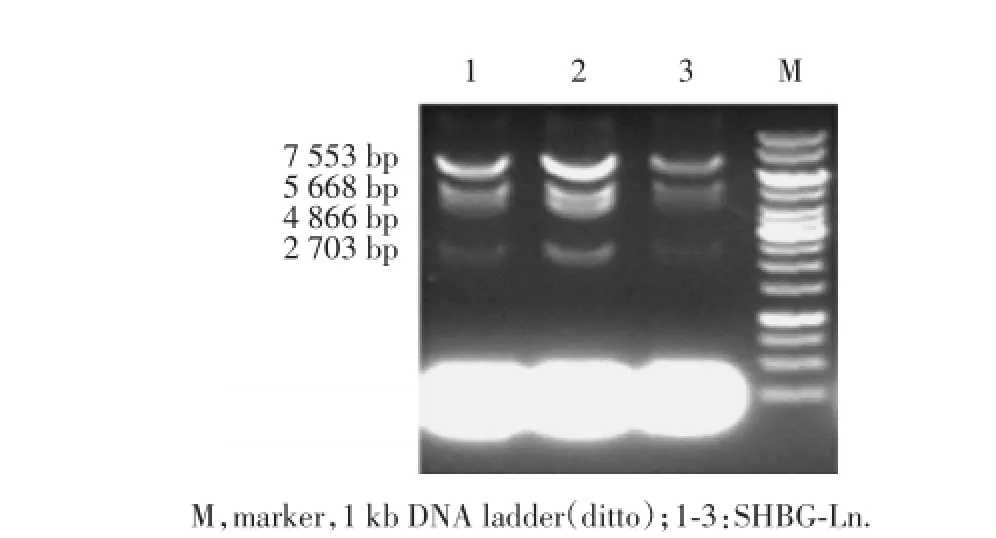
Fig.7 SHBG-Ln酶切的PCR擴增產物電泳結果Fig.7 Electrophoresis of PCR products of enzyme digestion of SHBG-Ln
SHBG-Ln-Cre質粒與含Neo的F片段共同轉入EL350菌株感受態細胞中,經過同源重組完成第2 ?次插入neo基因,構建載體pBR322-MK-SHBG-cko。pBR322-MK-SHBG-cko質粒經SalⅠ酶切,得到3個片段,分別為7 553 bp、6 764 bp和906 bp;SHBG-Ln-Cre質粒SalⅠ酶切,得到3個片段(圖8),分別為7 553 bp、4 866 bp和906 bp;酶切結果為2種質粒的疊加,應為4個片段(圖8),分別為7 553 bp、6 764 bp、4 866 bp和906 bp。
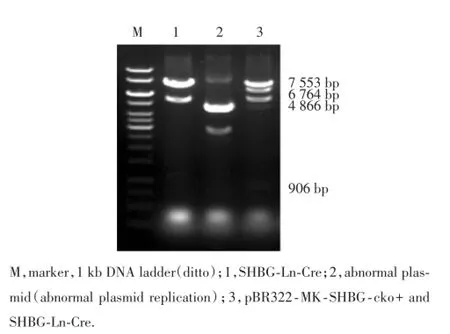
圖8 pBR322-MK-SHBG-cko酶切的PCR擴增產物電泳結果Fig.8 Electrophoresis of PCR products of enzyme digestion of pBR322-MK-SHBG-cko
2.6 純化及打靶載體的鑒定
考慮pBR322-MK-SHBG-cko耐藥性,將得到的質粒轉化DH5α感受態細胞,涂至含氨芐青霉素(50 μg/mL)LB板或卡那霉素(15 μg/mL)LB板,套取質粒將被去除,最后得到的只是pBR322-MK-SHBG-cko載體質粒,經SalⅠ酶切,得到3個片段(圖9),分別為7 553 bp、6 764 bp和906 bp。測序結果見圖10。
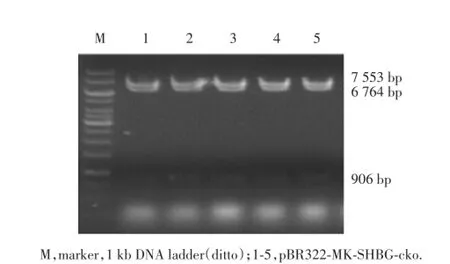
圖9 pBR322-MK-SHBG-cko酶切的PCR擴增產物電泳結果Fig.9 Electrophoresis of PCR products of enzyme digestion of pBR322-MK-SHBG-cko

圖10 pBR322-MK-SHBG-cko的測序圖Fig.10 Sequencing analysis of pBR322-MK-SHBG-cko
3 討論
基因敲除現主要有全基因敲除和條件性基因敲除2種方法。全基因敲除是通過將一段外源DNA序列(多數情況下是用于藥物篩選的新霉素抗性基因)來替換目標基因,進而使特定靶基因失活而使其所有組織器官中缺失的技術。條件性基因敲除主要是通過染色體位點特異性重組酶系統,例如Cre-LoxP系統和FLP-frt系統插入和重組來實現的,在待敲除的目標DNA序列兩端各插入一個LoxP(或Frt)序列,得到flox(Flanked by loxP)小鼠,將獲得的flox小鼠與帶有細胞特異性表達的Cre(或Flp)的小鼠交配繁殖,即可獲得在特定細胞里把目標基因不表達的小鼠,即條件性基因敲除小鼠[10]。其可以實現時間和組織特異性的特定基因的缺失表達,從而使對小鼠基因組的修飾的范圍和時間處于一種可控狀態[11]。目前對Shbg的研究多集中在細胞和分子水平及臨床試驗上,未曾建立相關小鼠模型,不除外Shbg具有致死性,故選擇條件性基因敲除。
基因載體的構建(即目的基因與運載體結合)是構建基因敲除小鼠的第一步,目前比較常用的基因打靶載體構建方法主要有兩種:一種是先通過PCR擴增各組分利用分子克隆來構建基因打靶載體,然后再分別進行酶切、連接和轉化篩選,這種方法工作周期相對長,至少3次酶切、連接和轉化反應;一種是通過合理借助Rac噬菌體的RecE/RecT或λ噬菌體Red操縱子(Redα/Redβ/Redγ)系統來介導實施同源重組反應,將從BAC載體中克隆出的同源重組基因(同源序列不必很長,50 bp即可)再次重組和改造進而獲得打靶載體。因其所需同源臂短,重組效率高而被廣泛應用[12,13]。本實驗中,我們2次利用Red/ET重組體系,一個是依托質粒pSC101-BAD-γβα-A-tet的重組系統,通過轉化重組質粒到含有BAC的細胞來實現DNA套取,避免了對BAC等DNA大分子的轉移操作,提高轉化效率,缺點是不除外最終所得修飾后的DNA中混有pSC101-BAD-γβα-A-tet。另一個是整合了重組蛋白基因的菌株EL-350,與pSC101-BAD-γβα-A-tet重組系統相似的是都可實現不再依賴限制性內切酶、連接酶的DNA重組,為構建打靶載體提供了新的選擇。EL-350使用相對簡單,不僅可反復轉化,重組效率高,并且避開了重組質粒對受體質粒的影響。我們在構建Shbg基因打靶載體的過程中,合理結合兩系統,使打靶載體的效率獲得提高。
通過以上步驟,2次利用Red-Cre重組酶系統成功構建Shbg的條件性敲除載體,為后續的制作基因敲除小鼠,進一步研究其生物學功能奠定了基礎。
[1]Hammond GL,Wu TS,Simard M.Evolving utility of sex hormonebinding globulin measurements in clinical medicine[J].Curr Opin Endocrinol Diabetes Obes,2012,19(3):183-189.
[2]Wallace IR,McKinley MC,Bell PM,et al.Sex hormone binding globulin and insulin resistance[J].Clin Endocrinol(Oxf),2013,78(3):321-329.
[3]Bartha JL,Comino-Delgado R,Romero-Carmona R,et al.Sex hormone-binding globulin in gestational diabetes[J].Acta Obstet Gynecol Scanda,2000,79(10):839-845.
[4]Mehrabian F,Rezae M.Sex hormone binding globulin measurement before conception as a predictor of gestational diabetes in women with polycystic ovarian syndrome[J].J Res Med Sci,2013,18(8):637-640.
[5]Sun L,Jin Z,Teng W,et al.Expression changes of sex hormone binding globulin in GDM placental tissues[J].J Perinat Med,2012,40(2):129-135.
[6]Sun L,Jin Z,Teng W,et al.SHBG in GDM maternal serum,placental tissues and umbilical cord serum expression changes and its significance[J].Diabetes Res Clin Pract,2013,99(2):168-173.
[7]Zhang Y,Buchhlz F,Muyrers JP,et al.A new logic for DNA engineering using recombination in Escherichia coli[J].Nat Genet,1998,20(2):123-128.
[8]Zhang Y,Muyrers JP,Testa G,et al.DNA cloning by homologous recombination in Escherichia coli[J].Nat Biotech,2000,18(12):1314-1317.
[9]Zhang Y,Muyrers JP,Rientjes J,et al.Phagean-nealing proteins promote oligonudeotide-directed mutagenesis in Escherichia coli and mouse ES cells[J].BMC Mol Biol,2003,16(1):1.
[10]Ray MK,Fagan SP,Brunicardi FC.The Cre-loxP system:a versatile tool for targeting genes in a cell-and stage-specific manner[J].Cell Transplant,2000,9(6):805-815.
[11]Nebert DW,Duffy JJ.How knockout mouse lines will be used to study the role of drug-metabolizing enzymes and their receptors during reproduction and development,and in environmental toxicity,cancer,and oxidative stress[J].Biochem Pharmacol,1997,53(3):249-254.
[12]王軍平,張友明.Red/ET重組及其在生物醫學中的應用[J].生物工程學報,2005,21(3):502-506.
[13]Testa G,Zhang Y,Vintersten K,et al.Engineering the mouse genome with bacterial artificial chromosomes to create multipurpose alleles[J].Nat Biotechnol,2003,21(4):443-447.(編輯武玉欣)
Construction ofTargeting Vector for ConditionalKnockoutofMurine Shbg
WANGYue1,JINZhen1,SUNLei1,ZHENG Jin-Hua2
(1.Department of Obstetrics and Gynecology,Shengjing Hospital,China Medical University,Shenyang 110004,China;2.Shanghai Research Center for Biomodel Organism,Shanghai201203,China)
Objective To construct the targeting vector for conditional gene knockout of sex hormone-binding globulin(Shbg)in mice.MethodsBased on Red/ET,two LoxP were inserted into both sides of extron 4 and extron 7 for conditional gene knockout of murine Shbg.Firstly,the Shbg gene and its upstream,downstream genes obtained from BAC DNAby PCRwere cloned into plasmid pBR322-MK,which was named pBR322-MKAB.The retrieve plasmid(pBR322-Shbg-Re)was obtained by homologous recombination between the plasmid and BAC.Then a great quantity of Neo fragments obtained from PL452 and PL451 were inserted into the targeting vector after another round of Red/ET and then the final targeting vector(pBR322-MK-SHBG-cko)was achieved.ResultsThe correctstructures ofthe targeting vectorssuch as pBR322-MK-AB,pBR322-Shbg-Re,SHBG-Ln and pBR322-MK-Shbg-cko were confirmed by restriction enzyme digestion and sequencing analysis.ConclusionThe targeting vector for conditional knockout of murine SHBG was successfully constructed.The construction of targeting vector paved the way for conditional knockout mouse strain generated by targeted mutation of Shbg.
sex hormone-binding globulin;conditional knockout;vector;Red/ET recombination
Q782;R394-33;R394.2
A
0258-4646(2015)03-0203-06
國家自然科學基金(81170591;81300511);沈陽市科學技術計劃項目(F11-262-9-46)
王越(1990-),女,碩士研究生.
金鎮,E-mail:jinzhen66@yahoo.com.cn
2014-09-11
網絡出版時間:
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
