不同激素配比對半支蓮細胞生長與甜菜紅素積累及抗氧化酶活性的影響
2015-01-08 07:57:38王玉嬌管恒潔
天然產物研究與開發 2015年12期
趙 胡,王玉嬌,管恒潔,劉 雪
阜陽師范學院生物與食品工程學院,阜陽 236037
甜菜素是一種含氮類吡啶衍生物水溶性色素,因最早在甜菜根中發現而得名,包括甜菜紅素(betacyanin)和甜菜黃素(betaxanthin)2 種形式[1]。尤其是甜菜紅素不僅作為傳統意義上食品添加劑的成分,而且作為重要的植物次生代謝物質在離體條件下具有很強的抗氧化和清除氧自由基能力,可以用來治療與過氧化脅迫有關的人體機能失調和抑制癌細胞增殖的功效[2,3]。因此,甜菜素的醫用價值最近幾年引起了人們的廣泛關注。
半支蓮(Portulaca grandifloraHook)系馬齒莧科馬齒莧屬一年生草本植物,因含豐富的甜菜紅素化合物,全草均可入藥,有清熱解毒、消腫止痛、活血化瘀的功效[4]。鑒于甜菜紅素藥用成分的特殊功效和在人們日常生活中的廣泛應用,發展甜菜紅素特有野生植物種植與開發,社會及經濟效益均極其顯著。近年來,隨著對甜菜紅素研究的不斷深入,國內外需求量顯著增加。但由于從野生植物資源中提取甜菜紅素,不僅受植物生長條件和季節的限制,而且提取量并不能滿足市場的供應需求。植物細胞培養技術生產次生代謝物克服植物受地理、季節等各種環境因素的影響,保證產物在一個限定系統中連續、均勻生產,有效地提高了人們獲取植物次生代謝產物范圍和生產規模。利用植物細胞生產有益人類需求的植物次生代謝物已受到人們廣泛的關注并已成功應用到多種藥用植物天然活性成分的生產[5]。國內外有關對半支蓮的研究主要集中在組織培養及再生體系建立,關于半支蓮細胞培養及甜菜紅素產物含量的報道還比較少見[4,6]。本試驗就不同植物激素配比對半支蓮細胞生長和甜菜紅素含量的影響進行了研究,并從細胞抗氧化酶系統方面對不同激素配比對甜菜紅素形成差異的成因進行初步分析和探討,為進一步用植物細胞培養技術生產甜菜紅素及研究甜菜紅素在植物細胞內的代謝調節機制提供基礎數據資料。
1 材料與方法
1.1 材料
半支蓮種子購自阜陽市花卉市場,在實驗室盆栽一個月后,以幼嫩的帶紫紅色莖段為外植體經0.1%升汞消毒、無菌水清洗后接種到MS +2,4-D(2.0 mg/L)培養基中,在25 ± 1 ℃光照下培養,光照強度為25 μmol/m2/s,光周期(白天/黑暗)為16 h/8h,培養15 d,誘導半支蓮愈傷細胞備用。
1.2 實驗設計
采用由不同激素配比類型的MS 固體培養基對半支蓮愈傷細胞進行培養以期篩選細胞生長量最大和甜菜紅素產量最高的激素配比類型培養基。半支蓮細胞培養基激素配比設計是以固定細胞分裂素的濃度為1.0 mg/L,生長素濃度分別為0.5 mg/L 和1.0 mg/L 為基礎,篩選兩種類型細胞分裂素(6-BA,KT)和三種類型生長素(NAA、IAA 和2,4-D)配比對細胞生長量和細胞色素表型的影響。確定生長素類型對細胞色素表型影響以后,進一步地細化濃度分別為0.1、0.5 mg/L 和1.0 mg/L 的不同細胞分裂素(6-BA,KT)對半支蓮細胞生長量和甜菜紅素含量的影響。
1.3 試驗方法與測試指標
在無菌操作室里,精確稱取色澤鮮嫩一致的半支蓮愈傷細胞1.00 ±0.01 g,分別接種于不同激素配比的MS 固體培養基中,重復三瓶,培養溫度與光照條件同上,培養期30 d。
1.3.1 愈傷細胞干重測定
取不同激素配比MS 固體培養基培養30 d 的愈傷細胞和原初接種半支蓮愈傷細胞1.00 ±0.01 g各3 份,濾紙吸去細胞中過多的水分后用鋁箔紙包裹細胞置于80 ℃干燥箱中烘干至恒重,稱其重量為干重。細胞增長量為:培養30 d 后細胞干重-原初接種半支蓮愈傷細胞干重。
1.3.2 甜菜紅素提取及含量測定
甜菜紅素提取及含量測定參照文獻方法并做修改[7]。具體做法:稱取1 g 新鮮半支蓮細胞加入20 mL 甲醇研磨,破碎細胞釋放出色素。10000 g 離心10 min,棄上清,沉淀細胞碎片用20 mL 去離子水浸提30 min,10000 g 離心10 min,沉淀再次用去離子水浸提,離心,直到細胞色素抽提殆盡,定容后,在538 nm 處分光光度計測定甜菜紅素的光吸收值,以上操作均在0~4 ℃下進行。根據甜菜紅素的摩爾消光系數56,600 L/cm/mol 計算甜菜紅素濃度及細胞中甜菜紅素含量。
細胞中甜菜紅素的含量為:
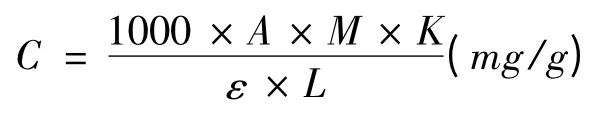
式中,A為吸光值;M為甜菜紅素平均分子量,K為稀釋倍數,ε 為甜菜紅素的摩爾消光系數;L為比色皿的口徑。
1.3.3 酶液提取
分別取不同激素配比培養下的半支蓮細胞1.00 ±0.01 g,按W∶V=1∶5 加入50 mmol/L pH 為7.8 的含有4 g/L 不溶性聚乙烯吡咯烷酮PVP 的磷酸緩沖液(PBS),加少許石英砂于冰浴中研磨成漿,用10000 g 于4 ℃離心10 min,棄沉淀,上清液即為酶提取液,待測。蛋白含量測定,按照Bradford 的方法測定[8],采用考馬斯亮藍G250 法于595 nm 波長測定光吸收值,根據繪制的標準曲線換算可溶性蛋白含量。
1.3.4 超氧化物歧化酶(SOD)活性測定
采用氯化硝基四氮唑藍(NBT)光化學還原法測定,參照文獻方法[9]。當光照時體系中產生氧自由基能還原NBT 形成藍色物質,在波長為560 nm 處測定消光值,以抑制光還原NBT 50%為1 個酶活性單位,酶活性以每毫克蛋白含酶量計算。
1.3.5 過氧化物酶(POD)和過氧化氫酶(CAT)活性測定
參照文獻方法進行[10]。當反應體系存在H2O2,POD 能使愈創木酚氧化,生成茶褐色物質,該物質在470 nm 處有最大光吸收值,以每分鐘內吸光值變化為0.01 為1 個酶活性單位。H2O2在240 nm處有最大光吸收值,過氧化氫酶能分解H2O2,使反應溶液吸光值隨反應時間而降低,根據測量吸光值的變化速率即可測出H2O2的活性。
1.3.6 數據處理
采用Microsoft Excel 2007 和SPSS19.0 相結合,對不同激素配比處理數據進行LSR-單因素最小顯著性差異分析(ANOVA),比較各處理之間差異顯著性,數據以平均值 ± 標準誤表示。
2 結果與分析
2.1 不同激素配比對半支蓮細胞生長量的影響
對在不同激素配比的MS 固體培養基上生長30 d 的半支蓮細胞生長量進行比較如圖1,結果表明:不同種類、不同劑量激素配比對半支蓮細胞生長量差異顯著。在相同細胞分裂素濃度下添加2,4-D 對細胞的增長量顯著高于NAA 和IAA。其中當向培養基中添加濃度均為1.0 mg/L 的2,4-D 和6-BA時,細胞的生長量最大,細胞干重達332.68 mg 分別是相同濃度下NAA 和IAA 處理下細胞干重的21.0倍和4.3 倍。當培養基中細胞分裂素為6-BA 時,NAA 和IAA 2 種生長素低濃度組(0.5 mg/L)處理對細胞生長量高于高濃度組(1.0 mg/L)處理;而1.0 mg/L 2,4-D 處理對細胞生長量高于0.5 mg/L濃度組處理,如在6-BA 或KT 為1.0 mg/L 時,1.0 mg/L 2,4-D 處理對細胞生長量分別比0.5 mg/L 濃度組處理高44.1%或47.1%。總體來看,2,4-D 處理下半支蓮細胞生長量高于NAA 和IAA 處理;而在相同濃度2,4-D 存在條件下,向培養基中添加6-BA 對半支蓮細胞生長效果要好于KT。
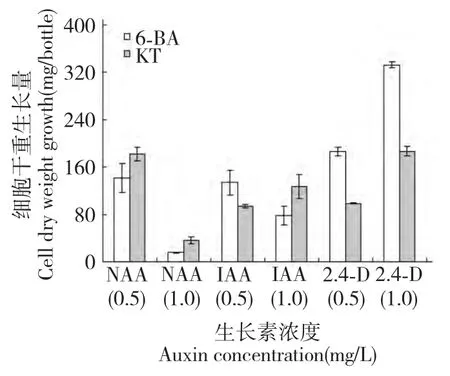
圖1 生長素對培養30 d 半支蓮細胞生長的影響Fig.1 Effect of auxins on 30 days-cultured P.grandiflora cell growth
2.2 不同激素配比對半支蓮細胞表型的影響
不同激素配比下半支蓮細胞色素表型差異見圖2。由圖2 可知,不同生長素類型及劑量對半支蓮細胞色素表型的影響差異顯著。在6-BA 為1.0 mg/L,向培養基中添加NAA,細胞沒有甜菜紅素產生,細胞生長緩慢,表型暗褐色,生長質量較差(圖2A)。從圖2B 和2C 可以看出如用2,4-D 代替NAA,細胞不僅生長質量大大改善,而且細胞也產生甜菜紅素,并隨著2,4-D 濃度的增加(圖2B 和2C 2,4-D 濃度分別為0.5 mg/L 和1.0 mg/L)細胞色素加深,表明2,4-D 顯著促進細胞產生甜菜紅素。
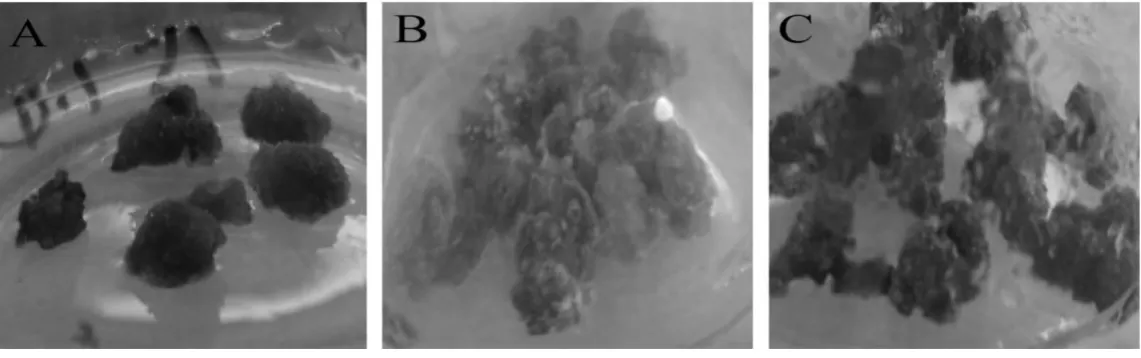
圖2 不同生長素對半支蓮細胞表型的影響Fig.2 Effect of different auxins on P.grandiflora cell phenotype
2.3 2,4-D 與不同濃度細胞分裂素配比對半支蓮細胞生長量的影響
由于2,4-D 對半支蓮細胞甜菜紅素的產生具有顯著地促進作用,進一步地實驗需要通過細化不同濃度細胞分裂素與2,4-D 結合篩選半支蓮細胞生長量最大的激素濃度組合,以期在細胞生產甜菜紅素的前提下進一步地篩選細胞生長量最大激素配比濃度。對在2 種2,4-D 濃度下3 種不同濃度的6-BA和KT 對培養30 d 半支蓮細胞生長量的影響見圖3。由圖可知,在2,4-D 濃度為1.0 mg/L、6-BA 濃度為0.5 mg/L 細胞生長量最大為465.50 mg,在此濃度下細胞生長量分別比6-BA 濃度為0.1 mg/L 和1.0 mg/L 高22.4%和28.5%,而KT 在各濃度組間對細胞生長量差異不顯著且低于6-BA 處理的細胞生長量。這一結果表明,在半支蓮細胞產生甜菜紅素的前體下,細胞生長量最大的激素配比為2,4-D濃度為1.0 mg/L 和6-BA 濃度為0.5 mg/L 激素組合。
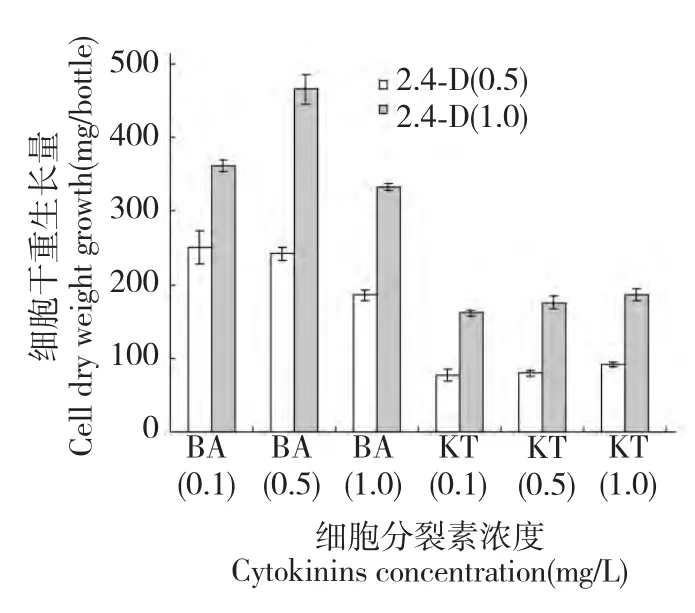
圖3 細胞分裂素對培養30 天半支蓮細胞生長的影響Fig.3 Effect of cytokinins on 30 days-cultured P.grandiflora cell growth
2.4 2,4-D 與不同濃度細胞分裂素配比對半支蓮細胞甜菜紅素含量的影響
2,4-D 與不同濃度細胞分裂素配比對培養30 d半支蓮細胞甜菜紅素含量的差異見圖4。由圖可知,與細胞生長量對2,4-D 與不同濃度細胞分裂素配比作用效應類似,在2,4-D 濃度為1.0 mg/L、6-BA 濃度為0.5 mg/L 細胞甜菜紅素含量最高達75.12 mg/g,在此濃度下細胞甜菜紅素含量比6-BA濃度1.0 mg/L 高24.8%,而與6-BA 濃度0.1 mg/L細胞甜菜紅素含量差異不顯著,暗示較高濃度的6-BA 不僅抑制細胞生長而且也降低細胞中甜菜紅素含量。而KT 在各濃度組間對細胞甜菜紅素含量差異不顯著且低于6-BA 處理下的細胞甜菜紅素含量。這一結果表明,半支蓮細胞甜菜紅素含量與細胞生長量對最適激素配比相吻合,細胞甜菜紅素最高的激素配比亦為2,4-D 濃度為1.0 mg/L 和6-BA濃度為0.5 mg/L 激素組合。
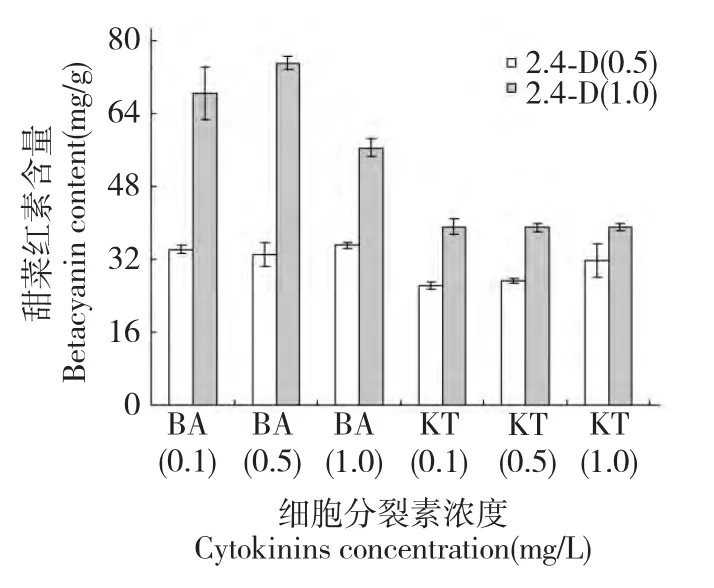
圖4 細胞分裂素對培養30 d 半支蓮細胞甜菜紅素含量的影響Fig.4 Effect of cytokinins on betacyanin content of 30 dayscultured P.grandiflora cell
2.5 不同激素配比對半支蓮細胞抗氧化酶活性的影響
根據上述的研究結果發現向培養基中添加生長素NAA 或IAA,半支蓮細胞不產生甜菜紅素,而添加生長素2,4-D 細胞開始變紅,且隨著2,4-D 濃度增加,細胞色澤越紅,表明2,4-D 對半支蓮細胞甜菜紅素合成的促進效應。為進一步地探討不同激素配比對半支蓮甜菜紅素合成差異的成因,本實驗測定了不同激素配比對培養30 d 半支蓮細胞參與活性氧代謝的抗氧化酶活性影響,以期分析活性氧代謝是否介導了半支蓮細胞甜菜紅素合成調控作用。由表1 可知,不同激素配比對半支蓮細胞抗氧化酶活性的影響差異顯著,當向培養基中添加抑制甜菜紅素合成生長素NAA 或IAA 時,SOD、POD 和CAT 的活性最低,而向培養基中添加濃度為1.0 mg/L 2,4-D時SOD、POD 和CAT 的活性最高,向培養基中添加濃度為0.5 mg/L 2,4-D 時SOD、POD 和CAT 的活性介于兩者之間。如在MS 培養基中2,4-D 為1.0 mg/L 和6-BA 為0.5 mg/L 激素組合培養的半支蓮細胞SOD、POD 和CAT 的活性分別是NAA 為0.5 mg/L 和6-BA 為1.0 mg/L 激素組合培養的半支蓮細胞相應酶活性的2.7 倍、3.1 倍和2.4 倍;比2,4-D 為0.5 mg/L 和6-BA 為0.5 mg/L 激素組合培養的半支蓮細胞相應酶活性高20.6%、30.8%和34.7%。這一結果表明,甜菜紅素激素誘導培養基上生長半支蓮細胞抗氧化酶活性顯著地高于不產色素的激素配比培養基上生長的半支蓮細胞。
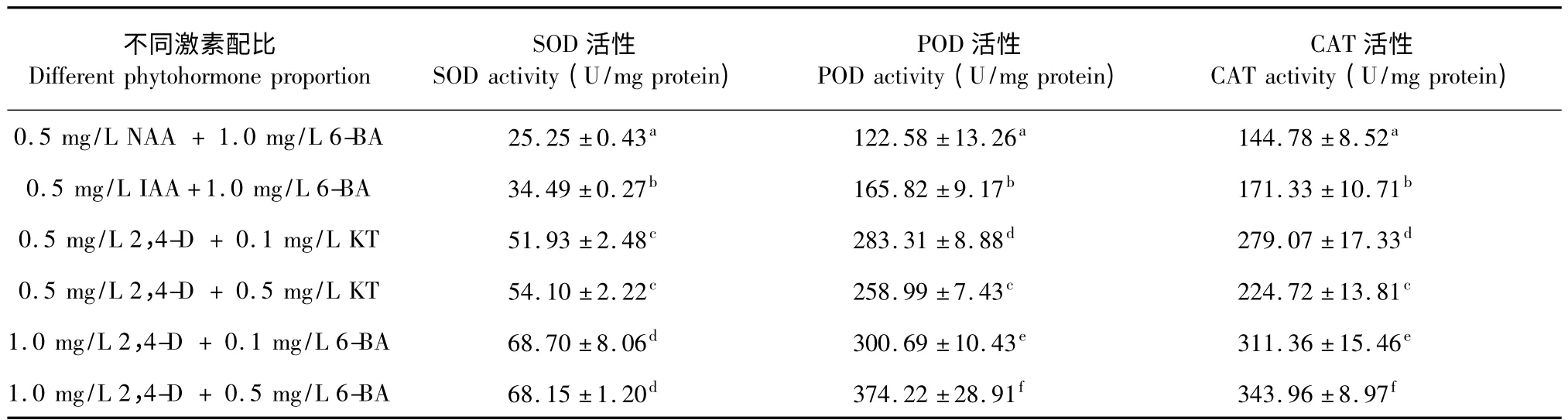
表1 不同激素配比對培養30 d 半支蓮細胞抗氧化酶活性的影響Table 1 Effect of different phytohormone proportion on antioxidant enzyme activities of 30 days-cultured P.grandiflora cell
3 討論
激素配比是影響植物細胞生長量或生長速率的一個重要因素。在植物細胞培養中,一般生長素在低濃度時能夠誘導細胞分裂,促進細胞伸長生長,濃度較高則會抑制生長,高濃度甚至會使植物受傷;細胞分裂素主要作用是促進細胞分裂。另外,細胞的生長狀況不僅受激素濃度的影響,還與激素種類有關。從本文的研究結果來看,2,4-D 促進半支蓮細胞的生長效果比NAA 和IAA 好,不僅表現在細胞的生長量高,而且細胞生長狀態也優于后兩種激素。生長素2,4-D 與細胞分裂素6-BA 搭配對促進半支蓮細胞的分裂效果要優于2,4-D 與KT 結合。許金光等[11](2007)發現在MS 培養基上生長素NAA 和細胞分裂素KT 各為3.0 mg/L 和0.1 mg/L 激素組合對三尖杉細胞生長量高于其他激素組合處理;而阮氏釧等[12](2015)研究不同激素配方對青錢柳細胞干物質量增加的影響,結果表明,在MS 培養基上NAA 1.0 mg/L 和KT 0.5 mg/L 激素組合最有利于細胞干重的增加。由此可見,細胞生長量積累所依賴的激素配比與物種緊密相關。
本文研究結果還發現,2,4-D 不僅具有促進半支蓮細胞生長的效果,而且作為誘導激素能明顯刺激細胞合成甜菜紅素,調控半支蓮細胞合成色素的最佳激素配比與細胞生長量的最佳激素組合相同,亦為2,4-D 1.0 mg/L 和6-BA 0.5 mg/L 激素組合,反應了半支蓮細胞生長與甜菜紅素的合成具有同步一致性,可能屬于細胞生長偶聯型的次生代謝模型。植物激素的種類與濃度及其配比方式不僅影響細胞生長,而且對細胞中次生代謝物積累也具有重要的調節作用。2,4-D 對瑞香狼毒細胞黃酮的積累是有利的[13],而對紫草細胞中紫草寧的合成具有嚴重的抑制作用[14]。由此可看出,不同植物激素對不同植物細胞次生代謝產物的積累有明顯的差異。在本試驗中發現,2,4-D 是最主要的影響因素,較高濃度的2,4-D 對半支蓮細胞的生長和甜菜紅素的積累都是有利的;而NAA 和IAA 對細胞生長和色素的積累不利。
為進一步地分析不同激素配比對半支蓮細胞甜菜紅素合成差異成因,本試驗初步分析不同激素配比對培養30 d 半支蓮細胞參與活性氧代謝的3 種抗氧化酶活性的差異,研究結果表明SOD、POD 和CAT 3 種酶活性與相應激素配比下的細胞中甜菜紅素的含量高度相關。2,4-D 為半支蓮細胞培養中甜菜紅素合成的誘導激素,向培養基中添加2,4-D 后,SOD、POD 和CAT 3 種酶活性顯著高于向培養基添加NAA 的3 種酶活性。SOD、POD 和CAT 活性提高表明細胞中和H2O2等活性氧代謝活躍,暗示活性氧可能參與了半支蓮細胞中甜菜紅素生物合成的調控。大量文獻報道和H2O2等活性氧分子參與植物的次生代謝調控,如參與黑曲霉誘導子對長春花細胞吲哚生物堿合成的促進作用[15]。Wang 等[16](2007)發現鹽地堿蓬根系在受到氧化脅迫時H2O2含量升高,H2O2作為信號分子轉運到葉片,導致葉片甜菜紅素含量增加。Gandía-Herrero 和Francisco García-Carmona[17](2013)認為多巴雙加氧酶是甜菜紅素生物合成途徑中關鍵限速酶,該酶催化將酪氨酸在酪氨酸酶羥化作用下形成的環狀多巴轉化為開環多巴,隨后自發反應生成甜菜紅素的基本生色團—甜菜醛氨酸。Casique-Arroyo 等[18](2014)研究發現各種生物或非生物脅迫引起多巴雙加氧酶基因表達上調進而導致千穗谷(Amaranthus hypochondriacusL)中甜菜紅素大量合成,推測可能的機制是各種環境脅迫引起H2O2含量上升,H2O2作為信號分子促進該酶基因表達引起甜菜紅素的積累。Wu 等[19](2004)研究經紫外線照射后的紅豆杉細胞后細胞中紫杉醇和H2O2含量增加,并進一步地發現H2O2通過提高細胞內源另一種植物激素茉莉酸(JA)合成途徑中關鍵酶基因脂氧合酶(LOX)基因的表達提高JA 的含量,JA 作為植物次生代謝產物合成正調控激素從而促進了細胞中紫杉醇的合成。從本試驗的研究結果來看,在含2,4-D 培養基中半支蓮細胞活性氧代謝活躍,甜菜紅素大量合成,表明細胞內產生的H2O2可能參與了半支蓮細胞甜菜紅素的合成,相關的H2O2調控半支蓮甜菜紅素合成途徑的研究目前正在進行。
1 Wang CQ (王長泉),Liu T (劉濤),Wang BS (王寶山).Advances in betalain research in higher plants.Chin Bull Bot(植物學通報),2006,23:302-311.
2 Taira J,Tsuchida E,Katoh MC,et al.Antioxidant capacity of betacyanins as radical scavengers for peroxyl radical and nitric oxide.Food Chem,2015,166:531-536.
3 Khan MI,Sri Harsha PSC,Giridhar P,et al.Pigment identification,nutritional composition,bioactivity,and in vitro cancer cell cytotoxicity ofRivina humilisL.berries,potential source of betalains.LWT -Food Sci Tech,2012,47:315-323.
4 Zhang HQ (張洪清).Regeneration and genetic transformation ofPortulaca grandiflora.Harbin:Harbin Normal University (哈爾濱師范大學),MSc.2009.
5 Qi FH (齊鳳慧),Zhan YG (詹亞光),Jing TZ (景天忠).A review on elicitors and their regulation on secondary metabolites in plant cell culture.Nat Prot Res Dev(天然產物研究與開發),2008,20:568-573.
6 Jain AK,Bashir M.In-vitropropagation of a medicinal plantPortulaca grandiflora.Hook.World J Agric Sci,2010,6:327-330.
7 Nakashima T,Araki T,Ueno O.Photoprotective function of betacyanin in leaves ofAmaranthus cruentusL.under water stress.Photosynthetica,2011,49:497-506.
8 Bradford MM.A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding.Anal Biochem,1976,72:248-254.
9 Beaucham C,Fridovic I.Superoxide dismutase:improved assays and an assay applicable to acrylamide gels.Anal Biochem,1971,44:276-287.
10 Chance B,Machly AC.Assay of catalase and peroxidase.Colowick SP,Kaplan NO.Methods in Enzymology.NewYork:Academic Press,1955,764-775.
11 Xu JG (許金光),Liu CJ (劉長江).Effect of different explants and medium composition on callus growth ofCephalotaxus fortuneiHook.f.Nat Prot Res Dev(天然產物研究與開發),2007,19:461-464.
12 Ruan SC (阮氏釧),Yang WX (楊萬霞),Fang SZ (方升佐),et al.Influence of types of basic medium,hormone formulations on accumulation of secondary metabolites inCyclocarya paliurussuspension cells.J Nanjing Forest Univ(南京林業大學學報),2015,39(2):47-51.
13 Wang WX (王文星),An Q (安琪),Wang Y (汪瑩),et al.A study on cell suspension culture and flavonoids accumulation ofStellera chamaejasme.Acta Prataculturae Sin(草業學報),2010,19:132-139.
14 Yazaki K,Matsuoka H,Ujihara T,et al.Shikonin biosynthesis inLithospermum erythrorhizon:light-induced negative regulation of secondary metabolism.Plant Biotech,1999,16:335-342.
15 Zhang XF (張向飛),Zhang RT (張榮濤),Wang NN (王寧寧),et al.Effects of fungal elicitors on accumulation of indole alkaloids inCatharanthus roseuscalli.Chin Tradit Herb Drugs(中草藥),2004,35:201-204.
16 Wang CH,Chen M,Wang BS.Betacyanin accumulation in the leaves of C3halophyteSuaeda salsaL.is induced by watering roots with H2O2.Plant Sci,2007,172:1-7.
17 Gandía-Herrero F,García-Carmona F.Biosynthesis of betalains:yellow and violet plant pigments.Cell,2013,18:334-343.
18 Casique-Arroyo G,Martínez-Gallardo N,de la Vara LG,et al.Betacyanin biosynthetic genes and enzymes are differentially induced by (a)biotic stress inAmaranthus hypochondriacus.Plos One,2014,9:99012.
19 Wu J,Ge X.Oxidative burst,jasmonic acid biosynthesis,and taxol production induced by low-energy ultrasound inTaxus chinensiscell suspension cultures.Biotech Bioeng,2004,85:714-721.
