杭州灣海域春秋季蝦蟹類群落結構及生物多樣性分析
2015-01-09 02:32:28周青松俞存根宋偉華李德偉
浙江海洋大學學報(自然科學版) 2015年1期
龐 敏,周青松,俞存根,宋偉華,李 新,李德偉
(1.浙江海洋學院水產學院,浙江舟山 316022;2.國家海洋局第二海洋研究所,浙江杭州 310012)
杭州灣海域春秋季蝦蟹類群落結構及生物多樣性分析
龐 敏1,周青松2,俞存根1,宋偉華1,李 新1,李德偉1
(1.浙江海洋學院水產學院,浙江舟山 316022;2.國家海洋局第二海洋研究所,浙江杭州 310012)
根據2012年5月(春季)、10月(秋季)2個季度月在杭州灣海域開展拖網漁業資源調查所獲得的蝦蟹類資料,用漁獲率作為蝦蟹類資源數量分布的指標,對杭州灣海域蝦蟹類的種類組成、數量分布、優勢種、生物多樣性等群落結構特征進行了分析。結果表明:杭州灣海域春、秋季共捕獲并鑒定出蝦蟹類12種,隸屬于3目6科9屬,其中,蝦類7種,蟹類4種,蝦蛄1種;調查海域春、秋季的優勢種均為安氏白蝦Exopalaemon annandalei,根據蝦蟹類的生活與生態習性分析得知,分布在本次調查海域的蝦蟹類可分為2種生態類群,以廣溫低鹽生態類群為主,廣溫廣鹽生態類群為輔;生物多樣性分析表明,調查海域的豐富度指數(D)、均勻性指數(J′)和多樣性指數(H′)均是秋季高于春季,這表明秋季蝦蟹類群落的豐富度、均勻性與多樣性都較春季高。另外,群落分析表明杭州灣蝦蟹類群落結構單一;蝦蟹類主要受溫度、鹽度、水深等環境因子影響,故其數量分布規律是春季在調查海域的內側或者說中西部蟹類分布多且集中,而灣口處蝦類分布較多,相反,秋季在灣口處蟹類分布較多,而蝦類在內側或者說中西部分布較多。
蝦蟹類;群落結構;多樣性;春秋季;杭州灣
杭州灣位于中國浙江省東北部、上海市南部,東臨舟山群島和著名的舟山漁場,北側與長江入海口相鄰,是錢塘江的入海通道。水文環境受江浙沿岸流及正規半日潮的影響,杭州灣海域環境條件優良,是舟山漁場的重要組成部分,歷史上盛產海蟄Rhopilma esculenta、鳳鱭Coilia mystus等,是許多經濟生物種類的繁殖、索餌和生長的棲息地。另外,杭州灣也是鰻苗溯河生長的通道。
過去,對東海、浙江近海及舟山漁場等較大空間尺度范圍的蝦蟹類曾開展過許多研究調查。如宋海棠等[1]研究報道了東海北部海域蝦類不同生態類群分布;宋海棠[2]分析了東海蝦類的生態群落與區系特征;俞存根等[3-5]分別研究報道了東海大陸架經濟蟹類種類組成及數量分布、群落結構特征以及春季東海蝦類群落結構特征;薛利建等[6]研究報道了東海中部海域蟹類群落結構及其多樣性;陳小慶等[7-9]報道了東海中南部外海蝦類、蟹類群落機構特征以及東海中北部海域蝦類群聚結構特征及空間分布,鄭獻之等[10]研究報道了舟山漁場及鄰近海域蟹類組成和時空分布;王甲剛[11]研究報道了舟山沿岸張網作業區蝦蟹類群落結構和多樣性;何賢保等[12]研究報道了舟山漁場及鄰近海域蝦類群落結構;而關于河口區或者沿岸海域的甲殼類群落結構研究較少有報道,主要有徐兆禮[13]分析了甌江口海域夏秋季蝦類資源分布特征;趙蒙蒙[14]對三門灣海域冬夏季蝦類的種類組成、時空分布及多樣性進行了分析研究;朱增軍[15]報道了夏秋季浙江岱衢洋蝦蟹類群落多樣性分析;瞿瀟南等[16]對舟山黃大洋海域春秋季蝦類種類組成和數量分布進行過報道,近年,國外對沿岸近海小空間蝦蟹類的研究也比較活躍,如MéLANIE BéGUER等[17]對法國Gironde河口葛氏長臂蝦種群動態的長期變化進行了研究;COSTA等[18]對巴西東北部的熱帶河口蝦類多樣性的季節變化進行了報道。但是針對杭州灣這一典型河口漁場的蝦蟹類資源狀況研究報道極少,相關的報道主要有吳常文等[16]報道了杭州灣海域安氏白蝦生物學與生態分析研究,至今還未見關于杭州灣蝦蟹類群落結構及生物多樣性的具體研究報道。
筆者主要根據2012年5月、10月在杭州灣海域開展漁業資源調查所獲得的蝦蟹類資料,分析研究了春、秋季杭州灣蝦蟹類的群落結構特征,以期為該海域的蝦蟹類資源可持續利用提供科學依據。
1 材料與方法
1.1 數據來源
本文所用的數據采自2012年5月(春季)、2013年10月(秋季)租用“浙普漁32267號”單拖網作業船在杭州灣口附近海域開展漁業資源調查所獲得的蝦蟹類資料。調查范圍為30°15′08.90″N-30°34′58.22″N,120°47′18.01″E-121°22′19.93″E,共設置13個調查站位。具體調查站位設置如圖1所示。

圖1 漁業資源調查站位圖Fig.1 The crustaceans resources survey stations of Hangzhou bay
調查船長為29 m,寬3.3 m,主機功率257 kW,單拖網網具規格為1 060目×80 mm。船上配備有探魚儀(測深儀)、對講機、衛導等輔助導航、定位、以及其他助漁助航機械設備。調查時每站的拖曳時長約1 h,拖速約為2.7 kn。調查采樣及生物學測定按照《海洋調查規范第6部分:海洋生物調查》(中華人民共和國國家質量監督檢驗檢疫總局,2007)[20]和《建設項目對海洋生物資源影響評價技術規程》(中華人民共和國農業部,2007)[21]進行。將每個站位拖網調查所獲得的漁獲物,全部裝入樣品袋,并在漁撈記錄表中進行記錄后,放入魚艙冰鮮保存,種類鑒定,尾數計算、稱重及測量體長等在實驗室內進行。稱重使用電子天平,精確度為0.01 g。漁獲數量統一標準化為每小時的漁獲量。調查海域的水溫、鹽度等測定采用CTD儀器與拖網調查同步進行。
1.2 數據分析與計算方法
1.2.1 相對重要性指數(IRI)計算
甲殼類優勢種主要根據PINKAS等的相對重要性指數(IRI)來確定,其計算公式如下:
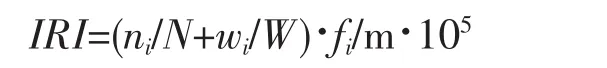
式中,ni、wi分別為第i種蝦蟹類的個體數和生物量(g);N、W分別為蝦蟹類總個體數和總生物量(g);fi為第i種蝦蟹類在m次取樣中出現的頻率;m為取樣次數。
1.2.2 生物多樣性指數計算公式
蝦蟹類生物多樣性的計算,主要采用Margalef豐富度指數(D)、Shannon-Weaver多樣性指數(H′)和Pielou均勻度指數(J′)計算公式,具體如下:
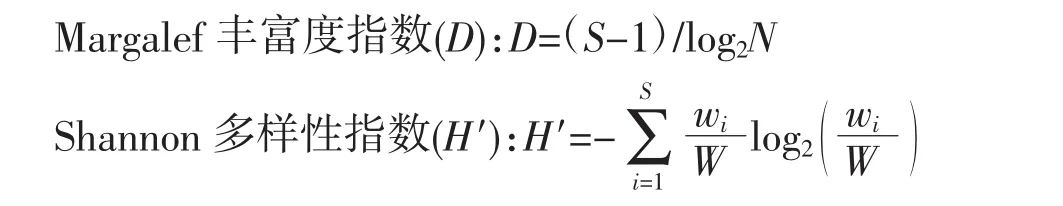
Pielou均勻度指數(J′):J′=H′蛐log2S
式中,S為蝦蟹類的總種類數;N為蝦蟹類的總個體數,wi為第i種生物的生物量(g),W為蝦蟹類的總生物量(g)。
1.2.3 群落的季節更替指數(AI)和遷移指數(MI)計算公式

式中,A為2個季節調查所得的實際物種數;C為某一個季節調查新遷入物種數;B為某一個季節調查遷出物種數;R為2個季節調查中均出現的物種數。AI反映物種更替導致群落穩定性降低的節律,該值越大則群落穩定性越小。MI為研究系統外遷入與遷出種的兩種成分在群落中的相對比例,當C〉B時,MI為正,表示遷入種多于遷出種;當C=B時,MI為0,即表示群落動態平衡。
1.3 數據處理
數據統計分析在Microsoft Excel上進行,站位圖和漁獲率的分布圖均采用Surfer8.0軟件進行繪制。
2 結果
2.1 種類組成
根據2012年春(5月)、秋(10月)在杭州灣海域所設的13個站位的拖網調查所獲得漁獲物樣品,共鑒定出蝦蟹類種類12種,其中蝦類7種,隸屬于3科、4屬;蟹類4種,隸屬2科、4屬;蝦蛄類1種。在蝦類種類組成中以長臂蝦科Palaemonidae種類最多,共有5種,藻蝦科Phippolytidae和鼓蝦科Alpheidae各為1種;蟹類則以梭子蟹科Protunidae種類為主,共有3種,方蟹科Grapsidae有1種。具體種類名錄以及出現季節情況如下:

?
本次調查共漁獲甲殼類7 111.1g,在不同種類的漁獲重量組成中,所占百分比最大的是安氏白蝦Exopalaemon annandalei(81.78%),其次是狹顎絨螯蟹Eriochier leptognathus(5.11%),細指長臂蝦Palaemon tenuidactylus(4.55%)脊尾白蝦Exopalaemon carinicauda(4.20%),其余種類所占漁獲物重量百分比則較小。同時,不同季節的漁獲物重量組成略有差異,如春季以安氏白蝦(83.80%)所占比重最高、狹顎絨螯蟹(7.79%)次之,秋季也以安氏白蝦(79.24%)所占比重最高,但具體比例要比春季低,其次是細指長臂蝦(9.78%)和脊尾白蝦(8.09%)。
2.2 優勢種
為確定不同蝦蟹類在群落中的重要性,講相對重要性指數(IRI)大于1 000的定義為優勢種,IRI值在100~1 000的定義為常見種。由此可知,春季的優勢種有安氏白蝦和狹顎絨螯蟹2種(表1),合占漁獲物總生物量的91.59%,合占漁獲物尾數的97.85%。秋季的優勢種則為安氏白蝦、脊尾白蝦和細指長臂蝦3種,合占漁獲物總生物量的97.10%,合占漁獲物尾數的98.25%。而常見種春季僅有鋸齒長臂蝦1種,秋季也僅有狹顎絨螯蟹1種。以上5種蝦蟹類為杭州灣調查海域的重要種類。

表1 杭州灣海域春秋季蝦蟹類相對重要性指數值(IRI)Tab.1 The index of relative importance(IRI)of The crustaceans in spring and autumn in survey area of Hangzhou bay
總體來說,調查海域春、秋季蝦蟹類相對重要性指數除了安氏白蝦IRI值變化不大外,其余種類的變化都較大。如春季的優勢種狹顎絨螯蟹春季的IRI值為1 086,到了秋季則急劇下降,僅為296。而春季IRI值較低的脊尾白蝦(48)和細指長臂蝦(0),到了秋季,其IRI值則上升為1 106和1 897,成為該調查海域的優勢種。
2.3 數量時空分布
2.3.1 季節變化
杭州灣春、秋兩季蝦蟹類數量群落結構組成變化不大(圖2)。均以蝦類占主導地位,其中春季,蝦類的生物量和密度分別占蝦蟹類生物量及總密度的 88.84%和96.88%,秋季分別占97.61%和98.98%;其次是蟹類,春季分別占蝦蟹類生物量及總密度的11.04%和3.10%,秋季分別占2.29%和1.01%;蝦蛄類資源較少,春季分別占蝦蟹類動物生物量及總密度的0.12%和0.02%,秋季分別占0.10%和0.01%,幾乎可以忽略不計。以上數據表明,春、秋季蝦蟹類生物量及總密度略有變化,蝦類秋季多于春季,而蟹類則相反,春季多于秋季。蝦蛄類春季和秋季變化不大。

圖2 杭州灣甲殼類群落結構Fig.2 Community structure of the crustacea in Hangzhou bay
2.3.2 數量分布
春季的蝦類漁獲率分布如圖3、4所示,從圖中可以看出,蝦類平均尾數漁獲率為464 ind/h,最高的是19號站,為1 299 ind/h,最低的是21號站,為57 ind/h。蝦類平均重量漁獲率為260 g/h,最高漁獲率出現在19號站,為609.3 g/h,其中漁獲數量最多的是安氏白蝦,約占該站位漁獲物重量組成的99.43%。漁獲率較高的站位依次11號站、15號站和10號站,漁獲率分別為384.5、376.4和373.7 g/h。漁獲率最低的出現在21號站,為36.9 g/h。
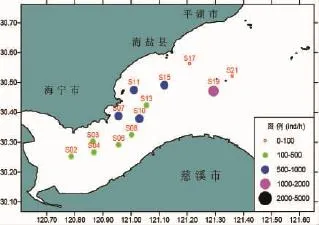
圖3 春季蝦類尾數漁獲率平面分布Fig.3 The distribution of the shrimp's mantissa CPUE in spring

圖4 春季蝦類重量漁獲率平面分布Fig.4 The distribution of the shrimp's weight CPUE in spring
秋季的蝦類漁獲率分布如圖5、6所示,從圖中可以看出,蝦類平均尾數漁獲率為715 ind/h,最高的是21號站,為2 136 ind/h,最低的是13號站,為163 ind/h;蝦類平均重量漁獲率為248.3 g/h,最高漁獲率出現在21號站,為759.5 g/h,其中漁獲數量最多的仍為安氏白蝦,約占該站位漁獲物重量組成的87.26%。漁獲率較高的站位依次17號站、2號站和3號站,漁獲率分別為470.0、309.1和305.7 g/h。漁獲率最低的出現在13號站,為68.8 g/h。

圖5 秋季蝦類尾數漁獲率平面分布Fig.5 The distribution of the shrimp's mantissa CPUE in autumn

圖6 秋季蝦類重量漁獲率平面分布Fig.6 The distribution of the shrimp's weight CPUE in autumn
春季的蟹類漁獲率分布如圖7、8所示,從圖中可以看出,蟹類平均尾數漁獲率為15 ind/h,最高的是2號站,為80 ind/h,最低的是1號站和19號站,僅為1 ind/h;蟹類平均重量漁獲率為32.3 g/h,最高漁獲率出現在2號站,為139.4 g/h,其中漁獲數量最多的為狹顎絨螯蟹,約占該站位漁獲物重量組成的35.17%。漁獲率較高的站位依次3號站和13號站,漁獲率分別為102.2和45.1g/h。漁獲率最低的出現在10號站,僅為0.7 g/h。

圖7 春季蟹類尾數漁獲率平面分布Fig.7 The distribution of the crab’s mantissa CPUE in spring

圖8 春季蟹類重量漁獲率平面分布Fig.8 The distribution of the crab’s weight CPUE in spring
秋季的蟹類漁獲率分布如圖9、10所示,從圖中可以看出,蟹類平均尾數漁獲率為7 ind/h,最高的是21號站,為25 ind/h,最低的是4號站和13號站,僅為1 ind/h;蟹類平均重量漁獲率為5.8 g/h,最高漁獲率出現在21號站,為23.5 g/h,漁獲率較高的站位依次11號站和17號站,漁獲率分別為102.2和45.1g/h。漁獲率最低的出現在4號站,為0.6 g/h。

圖9 秋季蟹類尾數漁獲率平面分布Fig.9 The distribution of the crab’s mantissa CPUE in autumn
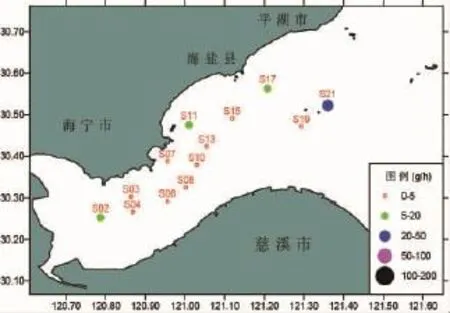
圖10 秋季蟹類重量漁獲率平面分布Fig.10 The distribution of the crab’s weight CPUE in autumn
綜上所述,杭州灣調查海域的不同季節蝦類尾數漁獲率變化是秋季〉春季,但從平均重量漁獲率來看,卻是春季略大于秋季。蟹類尾數漁獲率變化則是春季〉秋季,春季蟹類尾數漁獲率為秋季的2.14倍,而從平均重量漁獲率來看相差更為懸殊,春季(32.3 g/h)是秋季(5.8 g/h)的5.57倍。從地理分布趨勢上看,春季杭州灣調查海域的內側或者說中西部蟹類較多,而灣口處蝦類較多,相反,秋季在杭州灣的灣口處以蟹類較多,而在灣口和中西部均以蝦類較多。
2.4 生物多樣性
春季,Shannon-Wiener生物多樣性指數(H′)分布在0.06~1.40之間,平均值為0.79。Shannon-Wiener生物多樣性指數(H′)較大的站位分別為2、7、13、21號站位,最高值出現在21號站;秋季,Shannon-Wiener生物多樣性指數(H′)較春季有所上升,分布在0.32~1.66之間,平均值為1.07,Shannon-Wiener生物多樣性指數(H′)較大的站位為別為2、8、10、13、19號站,最高值出現在19號站。春秋兩季Shannon-Wiener生物多樣性指數(H′)最高值都位于杭州灣灣口,可能是由于錢塘江沖淡水帶來大量的營養物質,為此海域的蝦蟹類帶來了豐富的餌料。
春季,豐富度指數(D)分布在0.23~1.02之間,平均值為0.44。較大的站位分別為13、17、21號站位,最高值出現在21號站位(1.02),出現7種蝦蟹類。在3號站和6號站出現最低值(0.23),僅出現3種蝦蟹類。秋季,豐富度指數(D)分布在0.31~0.72之間,平均值為0.49。較大的站位分別為7、8、10、13、21號站位,最高值出現在21號站位(0.72),出現9種蝦蟹類。在3號站出現最低值(0.31),僅出現4種蝦蟹類。秋季較春季豐富度指數(D)變化不大。
春季,均勻性指數(J′)分布在0.03~0.71之間,平均值為0.37,站位間相關不大;秋季,均勻性指數(J′)分布在0.20~0.72之間,平均值為0.45,站位間相關也不大。各站位春秋季均勻性指數(J′)差別不大。

表2 春秋季杭州灣甲殼類生物多樣性Tab.2 The diversity indexes of Hangzhou bay in spring and autumn
2.5 群落季節更替與遷移
根據兩個季節對杭州灣海域調查所獲得的種類,杭州灣蝦蟹類生物種類數(A)、移入種數(C)和移出種數(B)和各航次均出現的種類數(R),計算出杭州灣海域春、秋兩季的群落更替指數AI=100和遷移指數MI=0。數據表明杭州灣蝦蟹類群落結構單一,穩定性一般,春、秋季的遷入與遷出種類數相等。
3 討論
3.1 種類組成及優勢種
春、秋兩季調查中,共鑒定出蝦蟹類種類12種,從春、秋兩季共同出現的蝦蟹類種類數(8種)及單一季節出現的種類(4種)來看,分布在杭州灣調查海域的季節性分布蝦蟹類較少,說明該海域蝦蟹類多數是長年在此生棲的種類,只有少數會進行季節性洄游;其次從春、秋兩季共同出現的8種蝦蟹類的生態習性來看,該海域的蝦蟹類種類以廣溫低鹽性種類為主,如安氏白蝦、脊尾白蝦、鋸齒長臂蝦,廣溫廣鹽性種類為輔,如細指長臂蝦、三疣梭子蟹、口蝦蛄。這與鄭獻之等、何賢保等在2006-2007年研究調查舟山漁場及鄰近海域蝦類、蟹類優勢種及生物多樣性有較大差異,據資料顯示,舟山漁場及鄰近海域優勢種大多為沿岸廣溫廣鹽性種,如葛氏長臂蝦,鷹爪蝦、細點圓趾蟹、三疣梭子蟹、口蝦蛄等,而在本次調查的海域,春、秋兩季都占絕對優勢的種類為沿岸廣溫低鹽性種類的安氏白蝦,除調查網具不同造成的漁獲組成差異外,更大的可能是因為本次調查海域地處河口區,與舟山漁場及鄰近海域相比,這里受錢塘江徑流等影響更為明顯,根據同步調查所獲得的溫度和鹽度數據表明,杭州灣海域春季表層溫度范圍為18.6~21.2℃,底層溫度范圍為19.6~21.0℃,表層鹽度范圍為5.3~7.5,底層鹽度范圍為5.3~7.6。秋季水體表層溫度為13.0~14.8℃,底層溫度范圍為13.0~14.7℃,表層鹽度范圍為9.7~12.1,底層鹽度范圍為9.1~12.1,該調查海域春、秋兩季水溫變化幅度較大,而鹽度低且變化較小。因此更適合廣溫低鹽性的安氏白蝦、脊尾白蝦、鋸齒長臂蝦的生長、繁殖。據吳常文等研究報道,安氏白蝦在20世紀80-90年代在杭州灣海域有廣泛分布,內側張網漁獲物中平均占11.41%,產量顯著高于脊尾白蝦,全年累計無論是出現尾數或是重量均居各種類之首位。安氏白蝦為廣溫低鹽性小型游泳蝦類,生長快,世代更新迅速,對環境適應能力強。一般4月中旬進入繁殖期,以抱卵親蝦為主,而9月份則以當年生的安氏白蝦占絕對優勢,親體基本消失。這也正好說明了本次調查中秋季安氏白蝦尾數漁獲率雖比春季多,但平均重量漁獲率卻還不及春季的原因。
3.2 數量時空分布特征
根據本次調查結果,春季,蟹類以調查海域的內側或者說中西部分布較多且集中,而蝦類以杭州灣灣口處分布數量較多。相反,秋季在杭州灣灣口處蟹類較多,而蝦類在內側或者說中西部較多。通過與相關的環境因子綜合分析,對安氏白蝦、脊尾白蝦、狹顎絨螯蟹等蝦蟹類而言,鹽度是決定其數量分布的主導因子。據同步調查所得的鹽度資料顯示,杭州灣海域春、秋季鹽度分布范圍為5.3~12.1,主要受錢塘江沖淡水和江浙沿岸水影響,鹽度分布呈現東南高,西北低。春季蝦類多分布于灣口進行產卵生殖活動,而秋季多分布于調查海域的內側或者說中西部。此外,杭州灣海域屬于溫度季節變化明顯的海域,春、秋季海水溫度的變化也會影響蝦蟹類的分布。如狹顎絨螯蟹的繁殖期一般在3-5月,適宜水溫在18~20℃,春季杭州灣海域內側水溫略比灣口高,故狹顎絨螯蟹集中分布在海域內側(2號站和3號站)。
3.3 生物多樣性分析
生物多樣性分析表明,杭州灣調查海域蝦蟹類的豐富度指數(D)、均勻性指數(J′)和多樣性指數(H′)整體偏低,且秋季均略高于春季。這表明該海域生物多樣性水平較低,群落結構單一,處于不穩定狀態,且秋季蝦蟹類群落的豐富度、均勻性與生物多樣性都較春季高。與陳小慶等[7-9]研究報道的東海中北部海域蝦蟹類、東海中南部外海蝦蟹類的生物多樣性分析比較,東海中北部海域蝦蟹類生物多樣性(春:H′=2.65,秋:H′=3.51)、東海中南部外海海域蝦蟹類生物多樣性 (春:H′=1.74,秋H′=1.95)均比杭州灣海域蝦蟹類生物多樣性 (春:H′= 0.79,秋H′=1.07)高得多。這可能與東海生態環境以及蝦蟹類的群落結構有關,東海蝦類主要以廣溫廣鹽生態類群為主,同時東海海域因受長江徑流、臺灣暖流等影響,營養物質以及餌料生物豐富,因此蝦蟹類種類豐富,生物多樣性高。而杭州灣海域為河口區,水文、底質等環境條件比較惡劣、多變。且受沖淡水的影響,鹽度較低,溫差較大,故在此條件下適合生存的蝦蟹類種類極少,且多為廣溫低鹽性種,故生物多樣性指數偏低。當然,生物多樣性指數偏低也有可能與杭州灣海域水體受到人為污染有關。根據2004年國家海洋環境監測中心提供的近岸生態系統健康狀況報告來看,杭州灣海域生態監控區從2004年起就處于不健康狀態,春季海域無機氮、活性磷酸鹽超過四類海水水質標準,水體嚴重富營養化導致浮游植物密度增加,產卵場退化。目前來看造成該海域污染的原因可能與生活工業污染、灘涂圍墾、海岸工程導致濕地面積減少、海水養殖業造成的海水富營養化、過度捕撈、航道影響及長江水入海量減少有關。杭州灣海域蝦蟹類生物多樣性偏低的具體原因尚無定論,有待進一步研究調查。建議繼續進行相關的調查和監測,以掌握杭州灣的水域環境及生物資源變動情況和影響機制。同時還應牢固樹立海洋漁業資源可持續發展的意識,加強漁業管理,設定休漁期、限制蝦蟹類捕撈規格,以實現該漁場蝦蟹類資源可持續利用之目的。
[1]宋海棠,丁天明.東海北部海域蝦類不同生態類群分布及其漁業[J].臺灣海峽,1995,14(1):67-72.
[2]宋海棠.東海蝦類的生態群落與區系特征[J].海洋科學集刊,2002(44):124-133.
[3]俞存根,宋海棠,姚光展,等.東海大陸架海域經濟蟹類種類組成和數量分布[J].海洋與湖沼,2006,37(1):53-60.
[4]俞存根,宋海棠,姚光展,等.東海蟹類群落結構特征的研究[J].海洋與湖沼,2005,36(3):213-220.
[5]俞存根,陳小慶,宋海棠,等.春季東海海域蝦類群落結構及其多樣性[J].生態學報,2009,29(7):3 593-3 604.
[6]薛利建,盧占暉.東海中部海域蟹類群落結構及其多樣性研究[J].南方水產科學,2011,7(6):66-71,
[7]陳小慶,俞存根,虞聰達,等.東海中南部外海蝦類群落機構特征分析[J].水生生物學報,2009,33(4):664-673.
[8]陳小慶,俞存根,虞聰達,等.東海中南部外海蟹類群落結構特征[J].應用生態學報學報,2009,20(10):2 527-2 534.
[9]陳小慶,俞存根,宋海棠,等.東海中北部海域蝦類群聚結構特征及空間分布[J].海洋學研究,2010,28(4):50-57.
[10]鄭獻之,俞存根,陳小慶,等.舟山漁場及鄰近海域蟹類組成和時空分布[J].海洋與湖沼,2012,43(1):147-153.
[11]王甲剛.舟山沿岸張網作業區蝦蟹類群落結構和多樣性的研究[D].舟山:浙江海洋學院,2012:20-43.
[12]何賢保,俞存根,鄭獻之,等.舟山漁場及鄰近海域蝦類群落結構分析[J].水生生物學報,2013,37(3):553-559.
[13]徐兆禮,沈昂綠,李新正.甌江口海域夏、秋季口足目和十足目蝦類分布特征[J].中國水產科學,2009,16(1):104-112.
[14]趙蒙蒙,徐兆禮.三門灣海域冬夏季口足目和十足目蝦類的種類組成、時空分布及多樣性分析[J].動物學雜志,2011,46 (3):11-18.
[15]朱增軍,張洪亮,虞寶存,等.夏秋季浙江岱衢洋蝦蟹類群落多樣性分析[J].浙江海洋學院學報:自然科學版,2012,31(3): 206-210.
[16]瞿瀟南,舒振宇,謝 旭,等.舟山黃大洋海域春秋季蝦類種類組成和數量分布[J].浙江海洋學院學報:自然科學版,2013, 32(4):285-291.
[17]COSTA M F,CAMARA M R.Diversity,abundance and seasonal variation of the ichthyofauna associated with shrimp farming in a tropical estuary in northeastern Brazil[J].Marine Biodiversity,2012,42(4):471-479.
[18]BéGUER M,BERGé J,GARDIA-PARèGE C et al.Long-Term Changes in Population Dynamics of the ShrimpPalaemon longirostrisin the Gironde Estuary[J].Estuaries and Coasts,2012,35(4):1 082-1 099.
[19]吳常文,王偉洪.杭州灣海域安氏白蝦生物學與生態分布的研究[J].浙江水產學院學報,1993,12(1):21-31.
[20]中華人民共和國國家質量監督檢驗檢疫總局.中國國家標準化管理委員會.GB 12763.6-2007海洋調查規范第6部分:海洋生物調查[S].北京:中國標準出版社,2007.
[21]中華人民共和國農業部.SC/T 9110-2007建設項目對海洋生物資源影響評價技術規程[S].北京:中國農業出版社,2008.
Community Structure and Diversity of Crustacea in Hangzhou Bay during Spring and Autumn
PANG Min1,ZHOU Qing-song2,YU Cun-gen1,et al
(1.Fishery School of Zhejiang Ocean University,Zhoushan 316022; 2.Second Institute of Oceanography,State Oceanic Administration,Hangzhou 310012,China)
Adopted in May(spring),October(autumn)2012 in the Hangzhou bay to carry out single trawl fishery resources survey data obtained the crustacean,the crustacean with catch rates as indicators of the distribution of the number of resources,species composition,dominant species,community structure and diversity.The results show that:12 crustacea species were found and identified in spring and autumn,which belonged to 3 orders,6 families and 9 genera.Including 7 species of shrimp,4 species of crab and 1 species of squilid.Exopalaemon annandaleiis the most important dominant species in the Hangzhou bay.The eurythermal and hypohaline community wasmain,and a eurythermal and eurysaline was minor in the survey area.In different seasons,the biodiversity analysis shows that the autumn’s species diversity(H′),species richness(D)and evenness(J′)higher than spring’s.Plus,the analysis of community showed that Hangzhou bay crustacean community species is single;Crustacea is mainly influenced by physical factors(such as temperature,salinity,water depth,etc.).So distribution of the catch rates showed that more crabs were caught at top of bay,i.e.mid-west of the survey area while more shrimps were caught at the mouth of the bay in spring.The opposite was happened in autumn.
crustacea;community structure;diversity;spring and autumn;Hangzhou bay
S932.5
A
1008-830X(2015)01-0001-08
2014-08-25
國家自然科學基金項目(31270527);企業委托項目(杭州灣區域建設用海項目海域生態環境現狀調查2012.03-2013.03)
龐敏(1992-),女,浙江金華人,研究方向:漁業資源.E-mail:471738360@qq.com
俞存根,教授.E-mail:cgyu@zjou.edu.cn
