水母消亡對海洋生態(tài)環(huán)境的影響
2015-01-19 02:57:22曲長鳳宋金明
生態(tài)學(xué)報 2015年18期
關(guān)鍵詞:影響
曲長鳳, 宋金明, 李 寧
1 中國科學(xué)院海洋研究所, 青島 266071 2 中國科學(xué)院大學(xué), 北京 100049
水母消亡對海洋生態(tài)環(huán)境的影響
曲長鳳1,2, 宋金明1,*, 李 寧1
1 中國科學(xué)院海洋研究所, 青島 266071 2 中國科學(xué)院大學(xué), 北京 100049
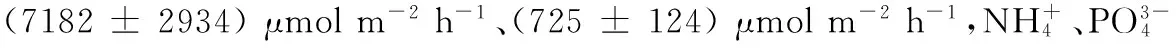
水母消亡; 生態(tài)環(huán)境影響; 生源要素
作為海洋大型浮游動物的水母,其種類多、數(shù)量大、分布廣,在浮游動物群落中占有相當(dāng)重要的地位。近年來不同種類水母在全球許多海域里頻繁出現(xiàn),在部分海域出現(xiàn)大量暴發(fā)[1-2],例如:地中海沿岸(西班牙、以色列沿岸)[3-4]、阿拉伯海沿岸(阿曼灣、波斯灣)[5]、亞洲東部海域(中國渤海、黃海、東海、日本海)[6-10]以及北大西洋與北太平洋的某些海域(愛爾蘭海、蘇格蘭沿岸、白令海)[11-12]等均出現(xiàn)過不同程度的水母暴發(fā)。大量水母聚集可導(dǎo)致浮游動物以及魚類大量減少,間接導(dǎo)致浮游植物增多,引發(fā)赤潮、綠潮等次生災(zāi)害,造成生物地球化學(xué)途徑和食物網(wǎng)的改變。水母暴發(fā)也可影響社會經(jīng)濟(jì)發(fā)展,對海洋漁業(yè)、沿海工業(yè)和旅游業(yè)帶來重大損害,甚至對人身安全和生命造成損失。不僅損壞漁具、拖網(wǎng),影響打撈作業(yè),減少漁獲量,影響船舶的正常航行,阻塞核電廠與發(fā)電廠的水循環(huán)系統(tǒng),而且還蜇傷游人,致使濱海旅游設(shè)施關(guān)閉,水母腐爛散發(fā)惡臭,影響濱海水質(zhì)安全。水母的暴發(fā)已經(jīng)成為近年來新型的近海生態(tài)災(zāi)害,嚴(yán)重影響海洋生態(tài)系統(tǒng)的結(jié)構(gòu)和功能及人類生產(chǎn),對海洋生態(tài)環(huán)境和人類活動造成較大威脅與損害。
水母的暴發(fā)及消亡機(jī)制研究已成為了當(dāng)前海洋生態(tài)環(huán)境科學(xué)的研究熱點之一,水母暴發(fā)后可在較短的時間段內(nèi)死亡,出現(xiàn)大量水母尸體聚集現(xiàn)象,對海洋生態(tài)系統(tǒng)特別是對生源物質(zhì)循環(huán)及生物群落有著重要影響,因此水母暴發(fā)-消亡機(jī)制及影響效應(yīng)研究至關(guān)重要。本文系統(tǒng)總結(jié)了水母消亡的觀測及方法、水母沉降速率及影響因素,重點探討了水母消亡對海水中生源要素、溶解氧、酸堿度及海洋生物群落的影響,對水母生物量的增加與海洋系統(tǒng)的耦合關(guān)系進(jìn)行了初步評估,以期為深入揭示水母消亡對海洋生態(tài)系統(tǒng)的影響以及系統(tǒng)的研究水母暴發(fā)消亡機(jī)制提供基礎(chǔ)。
1 海洋水母消亡的過程
1.1 水母消亡觀測
水母暴發(fā)通常可維持?jǐn)?shù)天或幾周,暴發(fā)后,由于水母老化死亡、食物缺乏以及水體環(huán)境中溫度改變等因素,水母開始下沉并伴隨死亡,垂死或死亡水母在下沉到海底過程中,水母體進(jìn)行分解。水母自然下沉消亡現(xiàn)象已在日本海、阿拉伯海、挪威海、紅海等許多海域被觀察到[13-17],研究者將海洋中這種膠質(zhì)動物下沉分解的現(xiàn)象命名為Jelly-falls[18],即水母下沉消亡。Jelly-falls被認(rèn)為是點源有機(jī)物質(zhì)的輸入,水母尸體在水體中下沉期間重新礦化為溶解有機(jī)無機(jī)組分,并最終引起海底水母類顆粒有機(jī)物質(zhì)的積聚。水母的季節(jié)性下沉往往發(fā)生在強(qiáng)烈海流上涌之后或溫帶/近極地地區(qū)春季水母暴發(fā)之后及晚春或早夏之后,其中水母暴發(fā)后消亡是水母下沉的最主要來源,所占比例大于75%[18]。水母下沉消亡現(xiàn)象可通過多種技術(shù)觀測到,主要包括大規(guī)模地區(qū)性措施,如建立深海天文臺網(wǎng)和近海科學(xué)平臺及與產(chǎn)業(yè)合作;中規(guī)模地方性措施,如遠(yuǎn)程控制潛水器,船只拖曳攝影、靜止攝影、定時攝影,拖網(wǎng),聲學(xué)/電學(xué)標(biāo)記研究;小規(guī)模措施,如沉積物捕獲裝置,自由漂浮剖析工具及沉積物基因工具等[18],目前應(yīng)用最為廣泛的為錄像技術(shù)和攝影技術(shù)。
水母下沉(Jelly-falls)始于垂死或死亡水母的垂直下沉,在水母下沉過程中,水母體或水母碎屑可被某些動物吞食,對于高能量的陸架邊緣海來說,這種情況極少發(fā)生,但對于食物短缺的深海,死亡水母也是一種有效的養(yǎng)分輸入;水母釋放的溶解有機(jī)物質(zhì)既可被浮游細(xì)菌和其他微生物當(dāng)作養(yǎng)料,也可被浮游動物吸收[19-22]。到達(dá)海水-沉積物界面的水母體去向主要包括:(1)水母體是某些海底食腐動物、無脊椎動物的食物來源,可能被食腐動物消耗掉進(jìn)而重新回到動物食物鏈中。(2)水母體釋放的有機(jī)物質(zhì)既可作為浮游細(xì)菌和其他浮游微生物的原料,供給微生物能量,對微生物群落有重要貢獻(xiàn);也可被小型和微型浮游動物吸收,將能量返還到浮游食物網(wǎng)中,其余未被消耗的部分有機(jī)無機(jī)物質(zhì)是營養(yǎng)鹽的重要來源,該過程是水母對海洋貢獻(xiàn)的主要過程。(3)水母剩余殘屑及水母體釋放的未被利用的顆粒態(tài)物質(zhì)會下沉到海底沉積物中,進(jìn)入生物地球化學(xué)循環(huán),除為海底底棲生物提供營養(yǎng)外,可增加深海層的沉積碳(C)、氮(N)、磷(P)量。
水母下沉到海水-沉積物界面后,開始迅速腐爛,一般在4—7 d內(nèi)一只完整的水母即可完全分解。腐爛初期,水母塊體迅速分解,水體變?yōu)樽丶t色,并有惡臭,水體表面開始出現(xiàn)一層白色薄膜;隨著消亡分解的進(jìn)行,水母塊體逐漸變小,水體顏色逐漸變淡變?yōu)樽攸S色,惡臭加劇,水體有粘性;腐爛后期,水體中水母已基本消亡完全,殘留白色絮狀物,并形成棕色沉淀物,水體變?yōu)榈咨杂袗撼簦恢了阁w完全消亡時,水體變?yōu)榈咨緹o粘性,水體底部有棕色沉淀[23-24]。水母消亡后的養(yǎng)分釋放及其與微生物的關(guān)系以及溶解氧的研究是水母消亡研究的主要內(nèi)容,也有學(xué)者研究水母消亡對海洋環(huán)境其它參數(shù)的影響,但是目前來說,水母消亡的研究仍相對較少。
1.2 水母的沉降速率
不同種類水母有不同的下沉深度,以至于一些水母種類可以運移至海底,成為大陸架邊緣和深海底棲生物群落的重要資源,而另一些在水體中被完全礦質(zhì)化。水母下沉到海底之后可能留在原地也可能被運移到其他地方[14],被當(dāng)?shù)氐膭游锖臀⑸锶郝湎牡簟K傅南鲁辽疃仁撬缸鳛樯锉眠M(jìn)行物質(zhì)與能量運移的重要影響因素。水母下沉深度受到很多因素的影響,既取決于水母本身的大小及暴發(fā)密度,又受沉降速率、腐爛速率、海水溫度、深度、海涌大小、海水流速等環(huán)境因素影響,還受外界食腐動物的清除速率及微生物的礦化速率等生物影響,這些影響可改變水母下沉?xí)r物質(zhì)與能量的分配。表1為不同種類水母的沉降方式及速率,在不考慮其他外界影響的情況下,水母的下沉速率非常快,沉降時大都成細(xì)長形、卵形或者球形,平均沉降速率在850—1500 m/d,大于浮游植物顆粒物和其它浮游動物的尸體,如水騷為140 m/d,片腳類為900 m/d,毛顎類為400 m/d,橈腳類為30—700 m/d,緣葉水母(Periphyllaperiphylla)的沉降速率更可達(dá)到3892 m/d[25]。
除受溫度、深度、流速、海涌以及外界生物等不可控的因素影響外,水母的沉降速度主要受自身性狀影響,水母的元素組成影響有機(jī)體的材料密度,沉降速度會隨水母生物量的增加而增加;另一個重要影響因素是水母體的形狀和沉降方向,水體的阻力系數(shù)會隨水母體形狀而改變,因此,呈球形或卵形的水母較細(xì)長形的水母沉降速度快,同時細(xì)長形水母往往有黏液尾巴,會增加阻力從而減少整體的沉降速率;此外礦質(zhì)化元素的變化也會影響沉降速率,例如消亡水母C/N比先迅速增加后降低[22],導(dǎo)致出現(xiàn)隨沉降深度增加而沉降速率降低的現(xiàn)象。水母的腐爛速率也是可變的,腐爛動態(tài)除受溫度、鹽度等因素影響外,與水母的自身重量有重要關(guān)系,Titelman等[22]將水母在消亡過程中的濕重與時間進(jìn)行擬合,曲線呈指數(shù)變化:y=ae-bx,式中y為濕重(g),x表示消亡時間(d),系數(shù)a隨著水母濕重的增加而增加,不同濕重的擬合曲線與系數(shù)差異顯著,系數(shù)b可作為腐爛系數(shù),其范圍在0.67—1.12 d-1之間,由于水母從200 m下沉到3000 m時,腐爛程度較小[18],因此可認(rèn)為在此距離范圍內(nèi)腐爛速率對沉降深度的影響較小。目前為止,由于缺少充足的水母沉降深度及沉降速率等的數(shù)據(jù)記錄,因此水母作為營養(yǎng)泵的運輸效率、運輸量等仍然未知。

表1 不同種類水母的沉降方式及沉降速率[25]
2 水母消亡對海洋環(huán)境的影響
水母消亡對海洋生態(tài)系統(tǒng)的影響概括起來主要包括兩方面,對海洋環(huán)境的影響以及對海洋生物的影響,具體可用圖1來描述。水母大量聚集死亡使水母攜帶的生源要素發(fā)生形態(tài)的變化和轉(zhuǎn)移[21,26-27]; 水母消亡時除自身消耗氧氣外,其不滲透性組分可能會沉到海底,導(dǎo)致海底沉積物中的氧氣輸入減少[14,24,26],形成低氧區(qū),給其它海洋生物的氧氣消耗帶來影響;水母消亡使周圍海水出現(xiàn)酸化現(xiàn)象,影響水體酸堿環(huán)境;海洋中以死亡水母為食的生物,例如食腐動物[14-17]可很好的消化水母碎屑,水母聚集消亡增加了浮游動物與微生物的豐度與生物量[13,28-29]。水母通過對海洋環(huán)境及海洋生物的影響改變了海洋生態(tài)系統(tǒng),還可引起其它海洋災(zāi)害的發(fā)生,例如富營養(yǎng)化、赤潮、綠潮及其它物種的暴發(fā)或者死亡等,增加的水母生物量可增加水母中物質(zhì)向海洋底層物質(zhì)的轉(zhuǎn)移,從而加強(qiáng)與水母有關(guān)的生物地球化學(xué)進(jìn)程和生態(tài)學(xué)進(jìn)程。

圖1 水母消亡過程及其與海洋生態(tài)系統(tǒng)的關(guān)系[18] Fig.1 Processes of jellyfish decomposition beginning with sinking to deposition at the seabed and its relationship with marine ecosystem[18] DOM:溶解有機(jī)物dissolved organic matter,DIM:溶解無機(jī)物dissolved inorganic matter,POM:顆粒有機(jī)物particulate organic matter, DOC:溶解有機(jī)碳dissolved organic carbon,DON:溶解有機(jī)氮dissolved organic nitrogen,DOP:溶解有機(jī)磷dissolved organic phosphorus,DIC:溶解無機(jī)碳dissolved inorganic carbon,DIN:溶解無機(jī)氮dissolved inorganic nitrogen,DIP:溶解無機(jī)磷 dissolved inorganic phosphorus
2.1 水母消亡對海水中生源要素的影響
水母消亡對海洋生態(tài)系統(tǒng)中生源要素的動態(tài)變化具有重大影響,水母是生源要素從浮游區(qū)域轉(zhuǎn)移到海底沉積物的媒介,也是有機(jī)物質(zhì)從深海環(huán)境進(jìn)入浮游食物鏈和其它海洋生態(tài)循環(huán)的營養(yǎng)途徑。水母暴發(fā)后消亡可引起大量養(yǎng)分的快速釋放,顯著改變周圍環(huán)境中養(yǎng)分狀況。由于水母下沉速度很快,水母的消亡多發(fā)生在沉積物-海水界面。水母消亡時,水母群變成一個大的養(yǎng)分儲存庫,釋放的碳、氮、磷既有可能溶解于海水中,也有可能降落到沉積物表面,成為沉積物的一部分,因此死亡水母是生源要素的重要來源,對海水及沉積物中生源要素的含量變化及形態(tài)分布有重大影響。
水母體水分含量非常高,超過97%[26],有機(jī)物質(zhì)的含量不足3%,屬低能量密度種群,能量組成僅為0.5—6 kJ/g干物重[27],水母體內(nèi)C、N含量分別約占水母干重的1.40%—42.75%與0.54%—13.64%,C/N僅為3.5%[26],高水分、低有機(jī)物質(zhì)含量及較低的C/N使水母極易腐爛。Larson等[26]將水母體的C、N含量與水母的無灰干重進(jìn)行了換算:C% DW=-5.20 + 0.50 AFDW% DW;N% DW=-0.95 + 0.13 AFDW% DW (AFDW:無灰干重;DW:干重),由于水母體C、N總量占到水母無灰干重的90%左右,因此認(rèn)為水母的無灰干重可很好的表征水母的C、N含量。雖然水母個體的能量密度較低,但是水母暴發(fā)后聚集的水母群體則會形成一個大的碳氮儲備庫,例如中國河北秦皇島2008年發(fā)生過海月水母的暴發(fā),清除水母達(dá)到了4000 t[10],按照海月水母干重占濕重的3.07%—3.91%,無灰干重占干重的25%—46%計算[28],此次水母暴發(fā)可提供的C和N量分別約為8.96—27.84 t和2.82—7.87 t,由此可見聚集的大量水母在海洋中可作為養(yǎng)分泵,增加海水與沉積物中的生源要素量,改變生源要素的資源分布。
大量水母消亡時,水母體是水體的主要碳源[14-17],可改變海水中碳的循環(huán)路徑。水母體中的碳既可以溶解有機(jī)碳的形式釋放到水體中,經(jīng)浮游細(xì)菌利用返還到浮游系統(tǒng)中,又可以顆粒碳的形式沉降到海底,增加沉積物中的碳沉積量。研究表明,水母體的碳含量與水母的濕重成正比,水母的濕重可通過以下公式來傳遞水母體的碳量:mg C/ind=0.867×g濕重/ind+ 20.85[29],由此可見水母聚集后死亡可形成大型碳庫,向外界輸送大量的DOC和POC,水母的數(shù)量越多,個體越大,輸送的碳量越多。水母死亡后向水體釋放DOC速率約為0.36 mg C g-1濕重 d-1[22],而直徑為9.5—18 cm的活體水母DOC的釋放率為0.007—0.016 mg C g-1濕重 d-1,平均值為0.012 mg C g-1濕重 d-1[21],可見死亡水母中DOC的釋放速率是活體水母30倍。以中國東海水母豐度較高的2003年為例,9月份水母的最高生物量約為30000 kg/h,即約為 1350 g/m2[30],當(dāng)水母消亡時,1 m-2的水母體每天可釋放的DOC約為480 mg C,雖然水母消亡僅僅持續(xù)幾天或幾周,但其影響是巨大的,東海中DOC的濃度范圍僅為0.44—12.03 mg/L[31]。水母下沉腐爛對下沉附近的沉積物中的碳貢獻(xiàn)可達(dá)(568±84) mg/m2[32],可見大規(guī)模的水母腐爛對水體中的碳循環(huán)影響巨大,可為海水-沉積物界面提供碳源。水母釋放的DOC經(jīng)浮游細(xì)菌消耗,增加細(xì)菌的碳代謝,使碳流入細(xì)菌群落,最終使水母體中的有機(jī)碳重新回到食物鏈中,將大量水母聚集的能量返還到高級營養(yǎng)級中,大量POC流入沉積物中增加了沉積物中的碳儲備,導(dǎo)致碳的生物地球化學(xué)功能的轉(zhuǎn)變。

綜上所述,水母在海洋生源要素的循環(huán)中扮演重要角色,顯著影響海洋生態(tài)系統(tǒng)中碳、氮、磷的循環(huán)。消亡水母作為一個大的養(yǎng)分泵,將水母體通過捕食而攝取的營養(yǎng)物質(zhì)經(jīng)腐爛分解向水體中釋放有機(jī)無機(jī)物質(zhì),使碳、氮、磷重生,改變海水中生源要素的分布形態(tài)與含量;消亡產(chǎn)生的含碳、氮、磷殘骸匯入海洋底部,一部分直接成為沉積物富集,一部分經(jīng)沉積物中微生物礦化進(jìn)入水體再循環(huán)。水母重生的無機(jī)物質(zhì)為浮游植物的初級生產(chǎn)提供了少量但重要的養(yǎng)分,溶解有機(jī)物質(zhì)可支持浮游細(xì)菌及微生物的生產(chǎn),可見,水母消亡釋放的生源要素不僅在海水及沉積物碳、氮、磷循環(huán)中起著重要作用,而且影響其他海洋生物的生存繁衍,對整個生物地球化學(xué)循環(huán)都有重要意義。此外,大量氮、磷的釋放導(dǎo)致海水局部養(yǎng)分含量的劇烈增加,可引起富營養(yǎng)化、赤潮、綠潮等次生生態(tài)災(zāi)害。
2.2 水母消亡引發(fā)的海水溶解氧的變化
水母較其他海洋魚類更能耐受和適應(yīng)低氧環(huán)境,低氧環(huán)境的出現(xiàn)為水母暴發(fā)提供了有利條件[37],反過來水母暴發(fā)后消亡對低氧環(huán)境的出現(xiàn)有著正反饋作用。首先大規(guī)模的水母聚集本身耗氧加劇,造成水母聚集區(qū)溶解氧的減少,水母死亡后,水母迅速下沉并自溶分解,水母組織細(xì)胞中的養(yǎng)分釋放到水體,使微生物活動加劇,耗氧量增加,同時不透水膠體碎屑將導(dǎo)致流入沉積物中氧氣的減少[14],可導(dǎo)致海底局部低氧或無氧環(huán)境。
水母消亡可引起海水中溶解氧濃度的顯著降低,并隨消亡的進(jìn)行保持持續(xù)的降低。West等[33]研究表明,馬賽克水母消亡過程中的最大沉積耗氧量可達(dá)(6161±3422) μmol m-2h-1,是無水母水體的9倍,整個消亡過程中平均沉積耗氧量相比無水母水體增加209%;宋金明等[23]研究發(fā)現(xiàn)沙海蜇(Nemopilemanomurai)消亡過程中平均耗氧量可達(dá)65.5 μmol kg-1h-1,水體飽和度僅為8.3%—13.3%,海水溫度和酸堿度的變化也可增加水母消亡耗氧量,由海水溫度和pH值變化形成的低氧區(qū)更為嚴(yán)重。水母尸體腐爛所釋放的大量養(yǎng)分導(dǎo)致細(xì)菌生物量和生長速率的增加[38],顯著改變周圍環(huán)境中的氧氣含量,這可能是水母消亡導(dǎo)致溶解氧消耗的主要原因。水體溶解氧的減少可引起其他不耐低氧生物的死亡,形成一系列連鎖反應(yīng),加重溶解氧的消耗,形成大范圍的低氧無氧區(qū)域;低氧可影響海洋生物行為、生長、生殖及其生存方式,導(dǎo)致某些海洋生物的豐度下降,改變海洋生態(tài)系統(tǒng)的種群組成及食物鏈的營養(yǎng)途徑,不僅危害海洋生物的多樣性,而且影響海洋生態(tài)系統(tǒng)功能的穩(wěn)定性;再者水體溶解氧的減少會增加海水-大氣界面溶解氧的梯度,使大氣中氧氣進(jìn)入海水,間接影響空氣中的氧氣含量,對陸地生態(tài)系統(tǒng)產(chǎn)生一定的影響。
2.3 水母消亡對海水酸堿度的影響

3 水母消亡對海洋生物群落的影響
通過對水母胃的內(nèi)含物組成分析[40]與穩(wěn)定同位素及脂肪酸組成分析[41]可判斷水母的飲食來源以及在食物鏈中的營養(yǎng)關(guān)系,浮游動物(如撓足動物和端足動物)、魚類幼蟲和魚卵都可作為水母的食物,浮游動物是水母的主要食物來源,水母的營養(yǎng)級在浮游動物和高級食肉魚類之間。水母作為浮游動物與魚類幼蟲及魚卵的捕食者以及其他以浮游動物為生的海洋動物的競爭者,大量水母聚集可影響浮游動物與魚類的數(shù)量,間接影響浮游植物的數(shù)量,改變浮游生物的聚集分布以及海洋系統(tǒng)的食物鏈,因此大量水母消亡可以緩解海洋浮游動物與魚類的捕食與競爭壓力。
水母消亡對海洋生物的數(shù)量變化有一定的影響,對海洋初級生產(chǎn)也有著重要的作用。水母消亡對海洋生物影響最大最直接的有兩類,首先是微型和小型浮游動物[38],水母體或者水母碎屑是某些海底食腐動物、無脊椎動物的食物來源,如珊瑚蟲[16]、甲殼類動物[13]、棘皮動物[16]等,腐爛的水母可作為海底碎屑的來源,由于深海中食物的限制,下沉水母可作為營養(yǎng)輸入[27],相比其他食物的分散,水母尸體大規(guī)模聚集可能導(dǎo)致食腐動物的聚集,水母釋放的營養(yǎng)物質(zhì)也可被浮游動物消耗掉[22,33];其次死亡水母的命運與為微生物群落組成有著重要關(guān)系[21,33],水母腐爛時水體周圍細(xì)菌顯著增多,水母釋放的溶解有機(jī)物質(zhì)(主要是DOC)是浮游細(xì)菌的主要溶解有機(jī)碳來源[19-20],細(xì)菌通過細(xì)胞外酶可分解利用水母釋放的大部分有機(jī)物質(zhì),水母消亡釋放大量養(yǎng)分也可導(dǎo)致細(xì)菌附著基質(zhì)數(shù)量和質(zhì)量的改變,影響細(xì)菌界的組成和活性。另外,水母下沉可為微型動物和微生物界提供生殖環(huán)境,固著生物體同樣可消耗水母釋放的有機(jī)營養(yǎng)物質(zhì)[16,42],同時水母消亡所引起的低氧/無氧以酸性環(huán)境可導(dǎo)致其他海洋生物的死亡。水母暴發(fā)后消亡對浮游動植物、微生物及其他海洋生物的影響可改變海洋生物資源的生物量、群落結(jié)構(gòu)及正常的生長代謝過程,導(dǎo)致海洋生物營養(yǎng)級及能量流動的變化,從而影響海洋生態(tài)系統(tǒng)的生物平衡;水母暴發(fā)后消亡可間接影響高營養(yǎng)級海洋漁業(yè)資源,使?jié)O業(yè)資源量及種群結(jié)構(gòu)發(fā)生改變,給人類的漁業(yè)資源帶來影響。
4 總結(jié)與展望
近年來,水母的異常暴發(fā)已成為世界許多海域頻繁發(fā)生的生態(tài)災(zāi)害,引起國際上廣泛關(guān)注,水母的大量繁殖對海洋漁業(yè)、沿海工業(yè)、旅游業(yè)以及海洋環(huán)境和海洋生態(tài)系統(tǒng)等都可能造成嚴(yán)重威脅,但水母暴發(fā)持續(xù)時間較短,暴發(fā)后的水母消亡對海洋生態(tài)系統(tǒng)的結(jié)構(gòu)和功能有重要影響。水母暴發(fā)后的消亡使水體產(chǎn)生不利于生物生存的惡劣環(huán)境,例如低氧、酸化等,但另一方面,其消亡可支持海洋生態(tài)系統(tǒng)的養(yǎng)分循環(huán),水母消亡釋放出的生源物質(zhì)又可直接參與海洋生物食物鏈的傳遞,維持海洋生物種群的繁衍,釋放的碳、氮、磷對海洋生源要素循環(huán)影響巨大,直接參與全球氣候變化和生態(tài)環(huán)境變化。但是由于水母本身非常脆弱,易破裂,難取樣,難觀測等特殊的生物學(xué)和生態(tài)學(xué)特性,使水母相關(guān)研究異常困難,加之對水母的生命周期、數(shù)量變動及與海洋環(huán)境的相互影響等缺乏相應(yīng)的科學(xué)數(shù)據(jù),需進(jìn)行大量的試驗以及海洋調(diào)查以彌補(bǔ)數(shù)據(jù)和證據(jù)方面的不足。雖然目前還沒有足夠的證據(jù)來證明全球水母數(shù)量增加是以后水母發(fā)展的一種必然趨勢,但不可否認(rèn)水母的增加已經(jīng)造成了人類的經(jīng)濟(jì)損失以及對海洋生態(tài)系統(tǒng)的破壞,目前為止水母暴發(fā)的機(jī)理、水母消亡與環(huán)境的相互關(guān)系以及水母暴發(fā)的預(yù)報治理等仍未有確切的定論,研究水母暴發(fā)消亡的原因、與環(huán)境的作用關(guān)系及如何應(yīng)對預(yù)防仍是一個世界性難題,因此對水母暴發(fā)消亡等的相關(guān)研究仍是科學(xué)界的關(guān)注重點,需加大力度。
今后水母消亡的生態(tài)環(huán)境效應(yīng)研究應(yīng)特別關(guān)注:1)水母作為養(yǎng)分泵的運轉(zhuǎn)機(jī)制,水母消亡時釋放的碳、氮、磷的形態(tài)變化、變化趨勢及變化原因;2)水母消亡時的物質(zhì)轉(zhuǎn)換與特征物質(zhì)的釋放,特征物質(zhì)的分離鑒定以及特征物質(zhì)對海洋生物的促進(jìn)、抑制作用研究,水母消亡的惡臭來源,水母消亡所引起的pH值變化和低溶解氧的根本原因;3)水母消亡可能引發(fā)的次生災(zāi)害,揭示水母消亡帶來的生態(tài)后果等。
[1] Brotz L, Cheung W W L, Kleisner K, Pakhomov E, Pauly D. Increasing jellyfish populations: trends in large marine ecosystems. Hydrobiologia, 2012, 690(1): 3-20.
[2] Mills C E. Jellyfish blooms: are populations increasing globally in response to changing ocean conditions?. Hydrobiologia, 2001, 451(1/3): 55-68.
[3] Licandro P, Conway D V P, Yahia M N D, De Puelles M L F, Gasparini S, Hecq J H, Tranter P, Kirby R R. A blooming jellyfish in the northeast Atlantic and Mediterranean. Biology Letters, 2010, 6(5): 688-691.
[4] Fuentes V L, Angel D L, Bayha K M, Atienza D, Edelist D, Bordehore C, Gili J M, Purcell J E. Blooms of the invasive ctenophore,Mnemiopsisleidyi, span the Mediterranean Sea in 2009. Hydrobiologia, 2010, 645(1): 23-37.
[5] Daryanabard R, Dawson M N. Jellyfish blooms:Crambionellaorsini(Scyphozoa: Rhizostomeae) in the Gulf of Oman, Iran, 2002—2003. Journal of the Marine Biological Association of the UK, 2008, 88(3): 477-483.
[6] Xian W W, Kang B, Liu R Y. Jellyfish blooms in the Yangtze estuary. Science, 2005, 307(5706): 41-41.
[7] 董華炳. 浙江部分漁場水母泛濫. 中國漁業(yè)經(jīng)濟(jì)研究, 2000, (1): 47-48.
[8] 葛立軍, 何德民. 生態(tài)危機(jī)的標(biāo)志性信號-霞水母旺發(fā)今年遼東灣海蜇大面積減產(chǎn). 中國水產(chǎn), 2004, (9): 23-25.
[9] Uye S I, Fujii N, Takeoka H. Unusual aggregations of the scyphomedusaAureliaauritain coastal waters along western Shikoku, Japan. Plankton Biology and Ecology, 2003, 50(1): 17-21.
[10] 劉旋, 李德剛. 當(dāng)海蜇群洶涌襲來. 華北電業(yè), 2008, (4): 66-69.
[11] Lynam C P, Lilley M K S, Bastian T, Doyle T K, Beggs S E, Hays G C. Have jellyfish in the Irish Sea benefited from climate change and overfishing? Global Change Biology, 2011, 17(2): 767-782.
[12] Brodeur R D, Decker M B, Ciannelli L, Purcell J E, Bond N A, Stabeno P J, Acuna E, Hunt G L Jr. Rise and fall of jellyfish in the eastern Bering sea in relation to climate regime shifts. Progress in Oceanography, 2008, 77(2): 103-111.
[13] Sweetman A K, Chapman A. First observations of jelly-falls at the seafloor in a deep-sea fjord. Deep Sea Research Part I: Oceanographic Research Papers, 2011, 58(12): 1206-1211.
[14] Billett D S M, Bett B J, Jacobs C L, Rouse I P, Wigham B D. Mass deposition of jellyfish in the deep Arabian Sea. Limnology and Oceanography, 2006, 51(5): 2077-2083.
[15] Murty S J, Bett B J, Gooday A J. Megafaunal responses to strong oxygen gradients on the Pakistan margin of the Arabian Sea. Deep Sea Research Part II: Topical Studies in Oceanography, 2009, 56(6/7): 472-487.
[16] Alamaru A, Bronstein O, Dishon G, Loya Y. Opportunistic feeding by the fungiid coralFungiascruposaon the moon jellyfishAureliaaurita. Coral Reefs, 2009, 28(4): 865-865.
[17] Yamamoto J, Hirose M, Ohtani T, Sugimoto K, Hirase K, Shimamoto N, Shimura T, Honda N, Fujimori Y, Mukai T. Transportation of organic matter to the sea floor by carrion falls of the giant jellyfishNemopilemanomuraiin the Sea of Japan. Marine Biology, 2008, 153(3): 311-317.
[18] Lebrato M, Pitt K A, Sweetman A K, Jones D O B, Cartes J E, Oschlies A, Condon R H, Molinero J C, Adler L, Gaillard C, Lloris D, Billett D S M. Jelly-falls historic and recent observations: a review to drive future research directions. Hydrobiologia, 2012, 690(1): 227-245.
[19] Condon R H, Steinberg D K, del Giorgio P A, Bouvier T C, Bronk D A, Graham W M, Ducklow H W. Jellyfish blooms result in a major microbial respiratory sink of carbon in marine systems. Proceedings of the National Academy of Sciences of the United States of America, 2011, 108(25): 10225-10230.
[20] Niggl W, Naumann M S, Struck U, Manasrah R, Wild C. Organic matter release by the benthic upside-down jellyfishCassiopeasp. fuels pelagic food webs in coral reefs. Journal of Experimental Marine Biology and Ecology, 2010, 384(1/2): 99-106.
[21] Hansson L J, Norrman B. Release of dissolved organic carbon (DOC) by the scyphozoan jellyfishAureliaauritaand its potential influence on the production of planktic bacteria. Marine Biology, 1995, 121(3): 527-532.
[22] Titelman J, Riemann L, S?rnes T A, Nilsen T, Griekspoor P, B?mstedt U. Turnover of dead jellyfish: stimulation and retardation of microbial activity. Marine Ecology Progress Series, 2006, 325: 43-58.
[23] 宋金明, 馬清霞, 李寧, 李學(xué)剛, 袁華茂, 段麗琴, 曲寶曉. 沙海蜇(Nemopilemanomurai)消亡過程中海水溶解氧變化的模擬研究. 海洋與湖沼, 2012, 43(3): 502-506.
[24] 馬清霞, 李寧, 李學(xué)剛, 袁華茂, 段麗琴, 曲寶曉, 宋金明. 模擬研究沙海蜇消亡過程中海水pH變化及對海水酸化的影響. 海洋科學(xué), 2012, 36(12): 12-18.
[25] Lebrato M, de Jesus Mendes P, Steinberg D K, Cartes J E, Jones B M, Birsa L M, Benavides R, Oschlies A. Jelly biomass sinking speed reveals a fast carbon export mechanism. Limnology and Oceanography, 2013, 58(3): 1113-1122.
[26] Larson R J. Water content, organic content, and carbon and nitrogen composition of medusae from the northeast Pacific. Journal of Experimental Marine Biology and Ecology, 1986, 99(2): 107-120.
[27] Doyle T K, Houghton J D R, McDevitt R, Davenport J, Hays G C. The energy density of jellyfish: estimates from bomb-calorimetry and proximate-composition. Journal of Experimental Marine Biology and Ecology, 2007, 343(2): 239-252.
[28] Lucas C. Biochemical composition ofAureliaauritain relation to age and sexual maturity. Journal of Experimental Marine Biology and Ecology, 1994, 183(2): 179-192.
[29] Schneider G. Chemische zusammensetzung und biomasseparameter der ohrenqualleAureliaaurita. Helgol?nder Meeresuntersuchungen, 1988, 42(2): 319-327.
[30] 丁峰元, 程家驊. 東海區(qū)沙海蜇的動態(tài)分布. 中國水產(chǎn)科學(xué), 2007, 14(1): 83-89.
[31] 張婷, 王作華, 石曉勇, 張傳松. 黃海、東海溶解有機(jī)碳的分布特征. 海洋環(huán)境科學(xué), 2011, 30(2): 162-166.
[32] Pitt K A, Koop K, Rissik D. Contrasting contributions to inorganic nutrient recycling by the co-occurring jellyfishes,CatostylusmosaicusandPhyllorhizapunctata(Scyphozoa, Rhizostomeae). Journal of Experimental Marine Biology and Ecology, 2005, 315(1): 71-86.
[33] West E J, Welsh D T, Pitt K A. Influence of decomposing jellyfish on the sediment oxygen demand and nutrient dynamics. Hydrobiologia, 2009, 616(1): 151-160.
[34] 李學(xué)剛, 李寧, 馬清霞, 袁華茂, 段麗琴, 宋金明. 沙海蜇(Nemopilemanomurai)模擬消亡過程中氮與磷的釋放. 海洋與湖沼, 2012, 43(3): 507-512.
[35] Pitt K A, Kingsford M J, Rissik D, Koop K. Jellyfish modify the response of planktonic assemblages to nutrient pulses. Marine Ecology Progress Series, 2007, 351: 1-13.
[36] Shimauchi H, Uye S I. Excretion and respiration rates of the scyphomedusaAureliaauritafrom the Inland sea of Japan. Journal of Oceanography, 2007, 63(1): 27-34.
[37] Purcell J E, Breitburg D L, Decker M B, Graham W M, Youngbluth M J, Raskoff K A. Pelagic cnidarians and ctenophores in low dissolved oxygen environments: a review // Coastal Hypoxia: Consequences for Living Resources and Ecosystems. Washington, DC: American Geophysical Union, 2001: 77-100.
[38] Tinta T, Malej A, Kos M, Turk V. Degradation of the Adriatic medusaAureliasp. by ambient bacteria. Hydrobiologia, 2010, 645(1): 179-191.
[39] 蔡楊鵬. 沙蜇Stomopholusmeleagris有機(jī)小分子化合物的提取、分離純化和結(jié)構(gòu)鑒定 [D]. 青島: 中國海洋大學(xué), 2011.
[40] Zavolokin A V, Glebov I I, Kosenok N S. Distribution, quantitative composition, and feeding of jellyfish in the Western Bering Sea in summer and fall. Russian Journal of Marine Biology, 2008, 34(7): 461-467.
[41] Cui Y, Wu Y, Zhang J, Wang N. Potential dietary influence on the stable isotopes and fatty acid compositions of jellyfishes in the Yellow Sea. Journal of the Marine Biological Association of the United Kingdom, 2012, 92(6): 1325-1333.
[42] Lebrato M, Jones D O B. Mass deposition event ofPyrosomaatlanticumcarcasses off Ivory Coast (West Africa). Limnology and Oceanography, 2009, 54(4): 1197-1209.
The effects of jellyfish decomposition on marine ecological environments
QU Changfeng1,2, SONG Jinming1,*, LI Ning1
1InstituteofOceanology,ChineseAcademyofSciences,Qingdao266071,China2GraduateUniversityofChineseAcademyofSciences,Beijing100049,China

jellyfish decomposition, eco-environmental effects, biogenic elements
國家重點基礎(chǔ)研究(973)項目(2011CB403602); 國家自然科學(xué)基金(41376092)
2014-01-24;
日期:2014-11-19
10.5846/stxb201401240177
*通訊作者Corresponding author.E-mail: jmsong@qdio.ac.cn
曲長鳳, 宋金明, 李寧.水母消亡對海洋生態(tài)環(huán)境的影響.生態(tài)學(xué)報,2015,35(18):6224-6232.
Qu C F, Song J M, Li N.The effects of jellyfish decomposition on marine ecological environments.Acta Ecologica Sinica,2015,35(18):6224-6232.
猜你喜歡
中學(xué)生數(shù)理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當(dāng)代陜西(2021年2期)2021-03-29 07:41:24
家庭影院技術(shù)(2020年10期)2020-12-14 07:54:18
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(shù)(2016年3期)2016-03-26 07:13:38
知識經(jīng)濟(jì)·中國直銷(2016年3期)2016-02-27 16:15:49
現(xiàn)代檢驗醫(yī)學(xué)雜志(2014年6期)2014-02-02 03:02:04
閱讀與作文(小學(xué)低年級版)(2011年3期)2011-01-01 00:00:00
