O3濃度升高和太陽輻射減弱對(duì)小麥根際土壤微生物功能多樣性的影響
2015-02-06 03:42:42吳芳芳鄭有飛吳榮軍王錦旗
生態(tài)學(xué)報(bào) 2015年12期
吳芳芳, 鄭有飛, 吳榮軍, 王錦旗, 李 萍
南京信息工程大學(xué)環(huán)境科學(xué)與工程學(xué)院,江蘇大氣環(huán)境監(jiān)測(cè)與污染控制高技術(shù)研究重點(diǎn)實(shí)驗(yàn)室, 南京 210044
O3濃度升高和太陽輻射減弱對(duì)小麥根際土壤微生物功能多樣性的影響
吳芳芳, 鄭有飛*, 吳榮軍, 王錦旗, 李 萍
南京信息工程大學(xué)環(huán)境科學(xué)與工程學(xué)院,江蘇大氣環(huán)境監(jiān)測(cè)與污染控制高技術(shù)研究重點(diǎn)實(shí)驗(yàn)室, 南京 210044
采用開頂箱(OTC)法和遮光網(wǎng)技術(shù),設(shè)置100 nL/L臭氧熏氣與3個(gè)輻射減弱梯度結(jié)合,模擬臭氧濃度升高和太陽輻射減弱的復(fù)合大氣背景。用BIOLOG生態(tài)測(cè)試板,采用孔平均顏色變化率法(AWCD)測(cè)定冬小麥根際土壤微生物利用不同碳源的能力,計(jì)算微生物群落多樣性指數(shù),對(duì)不同碳源的利用率進(jìn)行了主成分分析。兩年試驗(yàn)結(jié)果顯示,臭氧熏氣與太陽輻射減弱復(fù)合作用,降低了土壤微生物對(duì)碳源的利用速度和利用總量;除了聚合物以外其它碳源利用率顯著降低;對(duì)土壤微生物多樣性沒有直接的影響;對(duì)碳源降解的抑制效應(yīng)大于增強(qiáng)的O3與減弱的太陽輻射兩因素各自的單獨(dú)作用。太陽輻射減弱20%,一定程度上增加了對(duì)聚合物類的分解。O3熏氣條件下太陽輻射減弱,糖類、胺類代謝變異度較高,受環(huán)境影響較大。
臭氧增加; 太陽輻射減弱; 復(fù)合作用; 小麥; 微生物多樣性
Effects of ozone fumigation and depressed solar irradiance on soil microbial
目前我國華東地區(qū)大氣背景中O3濃度已達(dá)50 nL/L,已經(jīng)超過了敏感作物O3傷害閥值A(chǔ)OT40的25%[1],而且每年還以0.5%—2%的速度遞增[2]。空氣中SO2、NOx、黑碳、飛灰、土壤塵粒子等,引起大氣氣溶膠光學(xué)厚度逐年增加,導(dǎo)致地面變暗變冷[3]。僅亞洲大氣棕色云的直接輻射強(qiáng)迫引起的年均地表變暗就達(dá)14—16 W/m2(大約6%)[4]。近地層臭氧濃度增加和大氣氣溶膠引起的太陽輻射減弱等因素構(gòu)成了復(fù)雜的大氣背景。近地層大氣太陽輻射減弱和O3濃度增加是全球科學(xué)家和公眾密切關(guān)注的重要環(huán)境問題。地表O3和太陽輻射變化對(duì)土壤微生物的影響研究較少,Aneja M K等認(rèn)為O3濃度增加降低了土壤微生物的群落結(jié)構(gòu)[5]。陳展等認(rèn)為臭氧濃度升高降低了盆栽小麥根系的生長和土壤微生物功能[6]。Voidarou 等認(rèn)為O3濃度增加降低了環(huán)境中指示細(xì)菌的生長[7]。Johnson RM和李果梅分別報(bào)道了O3濃度升高對(duì)麥田土壤酶活性及酚酸類物質(zhì)含量產(chǎn)生影響[8-9]。太陽輻射減弱通過改變光合有效輻射(PAR),使作物光合作用受抑制,作物減產(chǎn),改變了碳、氮在植物中的分配[10- 14],改變了土壤中凋落物以及植物根系分泌物的數(shù)量和成分,改變了土壤溫、濕度和pH值[15- 17],對(duì)土壤生態(tài)系統(tǒng)產(chǎn)生的影響有大量報(bào)道,而對(duì)土壤微生物的影響研究很少,特別是地表O3濃度增加和太陽輻射減弱復(fù)雜大氣背景下,土壤微生物多樣性的響應(yīng)趨勢(shì)國內(nèi)外鮮見報(bào)道。土壤微生物多樣性影響土壤生態(tài)系統(tǒng)的結(jié)構(gòu)功能及過程,是維持土壤生產(chǎn)力的重要組分[18],土壤微生物種群的改變是土壤有機(jī)質(zhì)變化的早期指標(biāo)[19],可以敏感地指示氣候和土壤環(huán)境條件的變化[20]。土壤微生物多樣性在研究方法、研究范圍等方面得到了廣泛的重視,大量的研究認(rèn)為,植被、土壤的管理方式包括農(nóng)藥的應(yīng)用、施肥、耕作方式等,通過影響土壤有機(jī)碳和氮的水平、土壤含水量、溫度、通氣性及pH值等來影響土壤微生物多樣性[21- 24]。土壤生態(tài)系統(tǒng)通過植物倍受大氣環(huán)境的影響,Lou Y S報(bào)道紫外線增強(qiáng)降低了土壤微生物量碳氮的含量[25]。展小云等認(rèn)為,加倍CO2濃度顯著提高土壤微生物的活性及功能多樣性[26];Gallo M,張乃莉等,認(rèn)為氣候變暖和大氣N沉降改變了土壤微生物多樣性[6,27]。然而孤立地分析單一環(huán)境因子對(duì)地下生化過程的影響,無法準(zhǔn)確全面地評(píng)估土壤微生物群落對(duì)未來全球變化響應(yīng)的真實(shí)情況。因?yàn)樽匀簧鷳B(tài)系統(tǒng)中,各種全球變化現(xiàn)象間的交互作用無處不在。為了研究地表O3濃度增加和太陽輻射減弱對(duì)土壤微生物多樣性的交互影響,本文定點(diǎn)進(jìn)行了兩年試驗(yàn),研究土壤微生物碳代謝指紋的變化,計(jì)算微生物多樣性,為研究復(fù)雜大氣背景對(duì)土壤微生態(tài)系統(tǒng)的影響提供參考。
1 材料與方法1.1 大田試驗(yàn)概況
于2008年至2011年在南京信息工程大學(xué)農(nóng)業(yè)氣象試驗(yàn)站(32.16°N,118.86°E)進(jìn)行冬小麥生長季田間觀測(cè)試驗(yàn)。當(dāng)?shù)囟嗄昶骄鶞囟葹?5.6 ℃,多年平均降水為每年1 100 mm。供試土壤為潴育型水稻土(灰馬肝土屬),土壤質(zhì)地為壤質(zhì)黏土,耕層土壤黏粒含量為26.10%,土壤pH(H2O) 值為6.20,有機(jī)碳和全氮的含量分別為19.40 g/kg、1.15 g/kg。
1.2 臭氧增加與輻射減弱復(fù)合作用的大田試驗(yàn)設(shè)計(jì)
試驗(yàn)采用直徑3 m,高2.5 m 的開頂箱(OTC)作為臭氧布?xì)獾娜菁{裝置,O3發(fā)生器(江蘇省南京市萬杰臭氧機(jī)電設(shè)備廠,WJ-H-Y5 型)通過電解空氣中的O2產(chǎn)生O3;風(fēng)機(jī)把空氣和由發(fā)生器產(chǎn)生的O3混合后,由布?xì)庀到y(tǒng)均勻地吹送到作物冠層;采用O3檢測(cè)儀(新西蘭aeroQUAL公司,S200型)對(duì)氣室內(nèi)O3濃度進(jìn)行監(jiān)測(cè),通過閥門對(duì)室內(nèi)濃度進(jìn)行調(diào)節(jié)和控制。O3濃度設(shè)置為100 nL/L (T),為農(nóng)田本底濃度的2 倍。OTC內(nèi)設(shè)置臭氧濃度傳感器(精度:1 nL/L),對(duì)臭氧濃度傳感器進(jìn)行預(yù)先設(shè)定濃度閾值,當(dāng)OTC內(nèi)臭氧濃度未達(dá)到此閾值時(shí),臭氧發(fā)生器持續(xù)工作并向OTC內(nèi)通入臭氧氣體,當(dāng)OTC內(nèi)濃度超過此閾值時(shí),臭氧傳感器則提示報(bào)警,其信號(hào)通過微型電子控制儀的指令控制與臭氧發(fā)生器相連的電磁閥斷開,此時(shí)臭氧發(fā)生器停止工作。這樣可實(shí)現(xiàn)OTC內(nèi)臭氧濃度的自動(dòng)調(diào)控,每天臭氧處理時(shí)間段為08:00—16:00,遇有雨天則關(guān)閉整個(gè)裝置。
遮蔭棚采用鍍鋅鋼管為主要支柱,搭建成3 m×3 m×3 m的立方體,兩兩間隔3 m。遮蔭棚頂部用鐵絲搭建為可上下調(diào)節(jié)的網(wǎng)狀結(jié)構(gòu),用以支撐遮蔭網(wǎng),采用TBQ- 2型總輻射表對(duì)不同處理下達(dá)到冬小麥冠層的太陽總輻射強(qiáng)度進(jìn)行了監(jiān)測(cè),監(jiān)測(cè)時(shí)間為6:00—18:00,步長為1 min,每日實(shí)際監(jiān)測(cè)到的太陽總輻射強(qiáng)度瞬時(shí)值(W m-2min-1)(圖1)。通過計(jì)算確定各處理組實(shí)際接收的太陽輻射強(qiáng)度分別比對(duì)照減弱(20±5)%、(40±5)%、(60±5)%,田間大氣O3濃度和太陽輻射強(qiáng)度沒有人為干擾。并隨著作物生長高度,調(diào)節(jié)遮蔭網(wǎng)與冬小麥冠層間的距離,根據(jù)情況適當(dāng)更換遮蔭網(wǎng),保證網(wǎng)下空氣流通,使到達(dá)植物冠層的太陽總輻射變化量控制在預(yù)設(shè)值±5%范圍內(nèi)。在通入100 nL/L臭氧的OTC棚上方分別搭建不同透光度(網(wǎng)孔密度不同)的黑色聚乙烯遮蔭網(wǎng),使棚內(nèi)處于100 nL/L O3熏氣與太陽輻射分別減弱20%、40%、60%的復(fù)合環(huán)境中。試驗(yàn)設(shè)置T+S1(O3濃度為100 nl/L+太陽輻射強(qiáng)度減弱20%)、T+S2(O3濃度為100 nL/L+太陽輻射強(qiáng)度減弱40%)、T+S3(O3濃度為100 nl/L+太陽輻射強(qiáng)度減弱60%)三組雙因子處理,另設(shè)T(O3濃度為100 nl/L)和S1(太陽輻射減弱20%)的單獨(dú)作用,共5個(gè)處理,12個(gè)OTC棚和12個(gè)遮蔭棚,實(shí)現(xiàn)每處理3次重復(fù)。以不通入O3不加遮蔭網(wǎng)的OTC棚為對(duì)照。同步測(cè)定O3濃度和輻射通量,使O3誤差控制在8%,使輻射誤差控制在5%以內(nèi)。
1.3 田間管理和取樣
供試小麥品種為揚(yáng)麥13號(hào),水肥供應(yīng)適量而充分, 其他農(nóng)田管理措施均相同, 無病蟲害及雜草的影響。播種日期、熏氣起止日期、采樣日期以及AOT40累積量如表1所示。按五點(diǎn)取樣法,定點(diǎn)分別選取5株小麥,將其0—20 cm耕層根系區(qū)土樣挖出,抖掉根系外圍土,取緊貼在根表附近的土樣,混合后作為根際土,用四分法取適量裝于無菌紙袋中,立即帶回實(shí)驗(yàn)室,4 ℃下冰箱保存,用于土壤微生物碳代謝指紋的測(cè)定。
1.4 微生物多樣性的BIOLOG板檢測(cè)
土壤微生物碳源利用多樣性應(yīng)用BIOLOG生態(tài)測(cè)試板(ECO MicroPlate, 美國Matrix Technologies Corporation生產(chǎn))測(cè)定,Biolog EcoPlatesTM 技術(shù)是一種酶聯(lián)免疫吸附劑測(cè)定方法(ELISA,enzyme linked immunosorbent assay)[28]。每塊Biolog EcoPlate微孔板上有96 個(gè)孔, 3 份重復(fù),每份重復(fù)有32 孔(包括1個(gè)對(duì)照孔和31個(gè)單一碳源孔),除對(duì)照孔僅有水和指示劑以外,其余31 孔都裝有單一碳源和四唑染料。微生物利用單一碳源的代謝過程中產(chǎn)生氧化還原反應(yīng),使四唑染料變成紫色,根據(jù)每孔顏色變化程度檢測(cè)微生物的代謝能力。稱取抽穗期各處理新鮮土10 g,加入90 mL 0.85% NaCl無菌溶液于三角瓶中,無菌棉花塞封口后在搖床上震蕩(200 r/min)30 min,按10 倍稀釋法用0.85% NaCl無菌溶液將其稀釋至原來的1 /1000,稀釋液經(jīng)離心去除殘留的土壤后,上清液用于接種。接種懸浮液于生態(tài)測(cè)試板微孔板中,每孔150 μL,每樣3 次重復(fù),將接種好的測(cè)試板加蓋在(25±1)℃下暗箱培養(yǎng);連續(xù)培養(yǎng)10 d,期間每隔24 h用ELISA反應(yīng)微平板讀數(shù)器在590 nm處讀取吸光度[ 29- 30]。
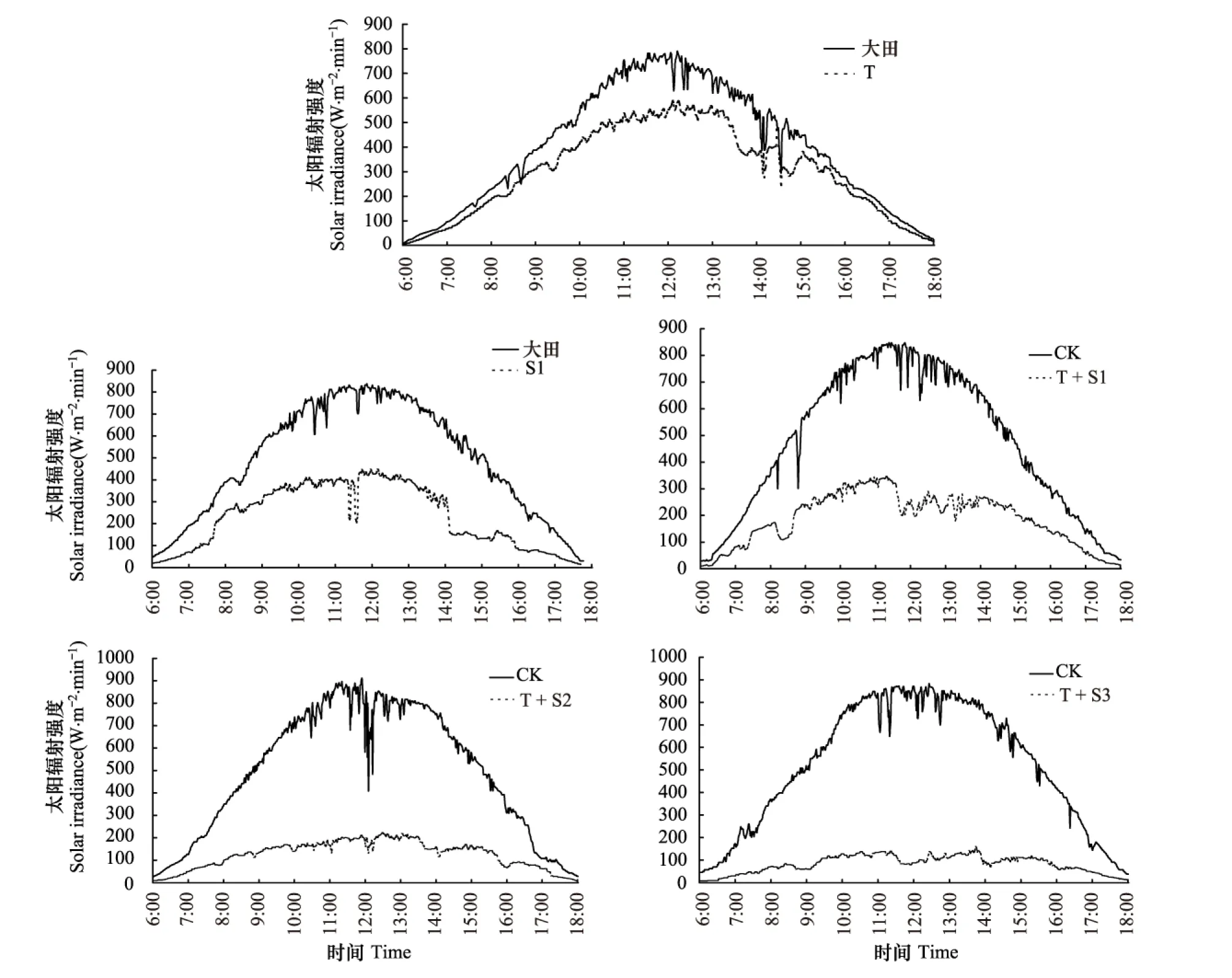
圖1 不同處理下冬小麥冠層太陽輻射強(qiáng)度
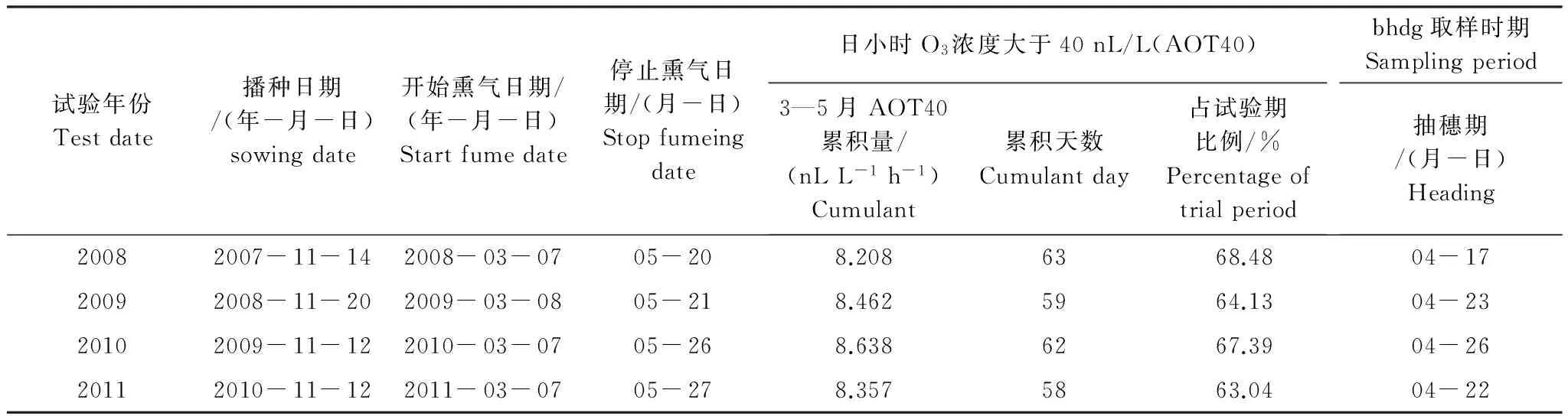
表1 大田熏氣和采樣時(shí)間表Table 1 Fumeing and sampling date
1.5 數(shù)據(jù)處理及統(tǒng)計(jì)分析
采用孔平均顏色變化率法(AverageWelL-Color Development, AWCD)測(cè)定土壤微生物利用單一碳源的能力[28]。AWCD的計(jì)算公式為:
AWCD =∑(Ci-R) /n
式中,Ci為各反應(yīng)孔在590 nm下的吸光度,R為對(duì)照孔的吸光值,Ci-R≤0的孔在計(jì)算中記為0。n為培養(yǎng)基碳源種類數(shù),本研究中為31。
當(dāng)能觀測(cè)到大部分微孔中微生物群落的反應(yīng),而且大部分活躍的微生物群落已達(dá)到顏色變化的漸近線時(shí)的培養(yǎng)時(shí)間為96 h,因此選擇培養(yǎng)96 h時(shí)的數(shù)據(jù),采用DPS軟件,根據(jù)土壤微生物群落對(duì)31 種單一碳源利用的相似性,應(yīng)用表2中的方法計(jì)算多樣性指數(shù),進(jìn)行新復(fù)極差檢驗(yàn)和主成分分析。

表2 土壤微生物群落功能多樣性指數(shù)的計(jì)算公式Table 2 The equations of function diversity indexes of soil microbe community
2 結(jié)果與分析
2.1 O3熏氣條件下太陽輻射減弱土壤微生物利用碳源的動(dòng)力學(xué)特征
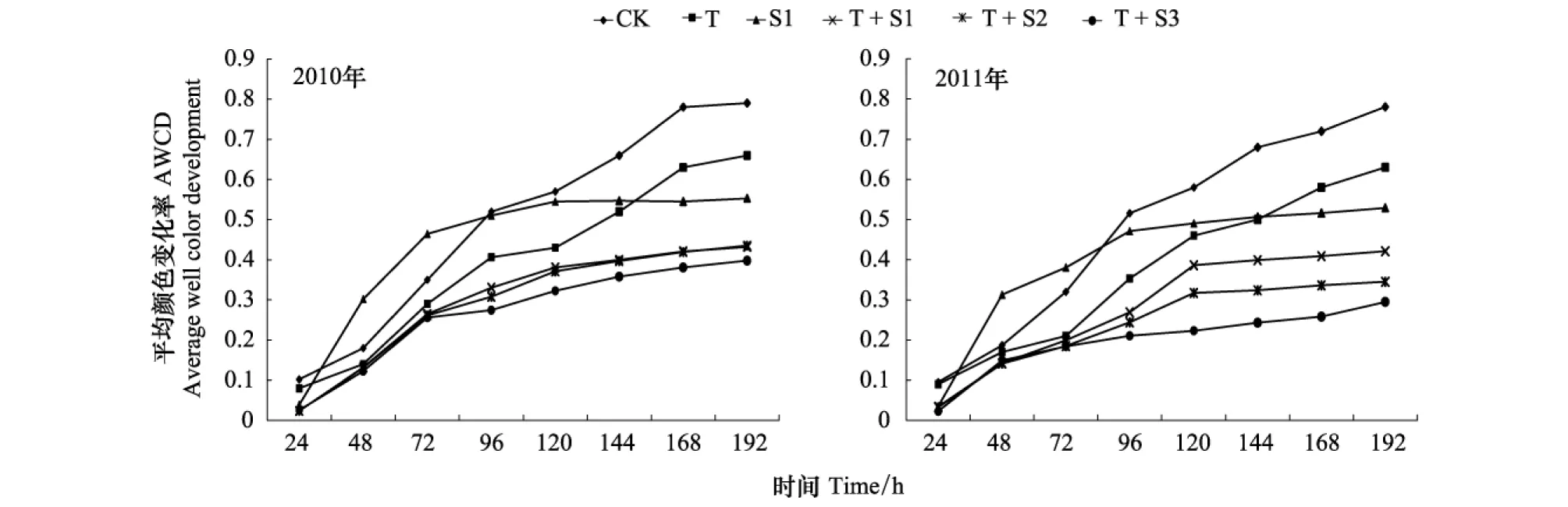
圖2 不同處理土壤平均顏色變化率(AWCD) 192h培養(yǎng)期間的變化
2010年與2011年兩年試驗(yàn)結(jié)果顯示,O3熏氣與太陽輻射減弱雙因子處理下,土壤微生物對(duì)碳源的降解速率(24—96 h降解曲線的斜率)和最終AWCD 值都顯著低于CK、T和S1,如圖2所示。這說明,增強(qiáng)的O3與減弱的太陽輻射兩種因素雙因子處理對(duì)碳源降解速率的抑制效應(yīng)大于它們的單獨(dú)作用。單獨(dú)O3熏氣(T),土壤微生物對(duì)碳源的降解速率、AWCD最大值均顯著低于對(duì)照。太陽輻射減弱20%單獨(dú)作用(S1),土壤微生物對(duì)碳源降解速率與CK無顯著差異,但AWCD最大值顯著降低。結(jié)果說明,O3熏氣降低了土壤微生物碳源利用的速率和最終碳源利用總量,太陽輻射減弱20%時(shí)對(duì)土壤微生物的碳源降解速率沒有顯著影響,但降低了最終碳源利用總量。
2.2 O3熏氣和太陽輻射減弱土壤微生物對(duì)不同類別碳源的利用強(qiáng)度

圖3 O3熏氣和太陽輻射減弱下土壤微生物對(duì)不同類型碳源的利用
采用土壤ELISA反應(yīng)96 h的數(shù)據(jù)分析土壤微生物對(duì)不同類別碳源的利用強(qiáng)度,從圖3可看出,T,T+S1,T+S2,T+S3處理顯著降低了碳源利用總量,S1對(duì)碳源總量的利用則沒有顯著影響。不同類型碳源的利用強(qiáng)度如表3所示,糖類、氨基酸、羧酸、酚酸和胺類的利用率在T、T+S1、T+S2、T+S3作用下顯著降低,S1對(duì)它們則沒有明顯影響。聚合物類的AWCD值較小,說明難以被微生物分解,S1卻能顯著提高聚合物的利用強(qiáng)度。兩年試驗(yàn)變化趨勢(shì)相同。結(jié)果說明,適度的太陽輻射減弱(減弱20%),一定程度上增加了對(duì)聚合物類的分解,O3濃度單獨(dú)增加處理,以及O3熏氣和太陽輻射減弱雙因子處理下,除了聚合物類以外其它碳源利用率顯著降低。增加的O3和減弱的輻射兩因素復(fù)合作用比O3單因素作用對(duì)碳源利用強(qiáng)度的抑制效應(yīng)強(qiáng)。
2.3 O3熏氣和太陽輻射減弱對(duì)土壤微生物群落功能多樣性指數(shù)的影響
O3熏氣和太陽輻射減弱條件下,土壤微生物群落功能多樣性指數(shù):Simpson指數(shù)(D)、 Shannon指數(shù)(H′)、McIntosh(Dmc)指數(shù)以及Simpson均勻度在兩年的試驗(yàn)中均沒有顯著變化;只有評(píng)估微生物豐富度的Brillouin指數(shù)(H)比對(duì)照及S1處理低(表4)。可見增強(qiáng)的O3與減弱的太陽輻射處理,對(duì)麥田土壤常見微生物類群影響不大,對(duì)不同種類微生物生長繁衍的刺激作用沒有很大差異,沒有使某些物種大幅度增長或減少,群落均勻性變化不顯著。微生物豐富度是否降低還需長期試驗(yàn)觀察。
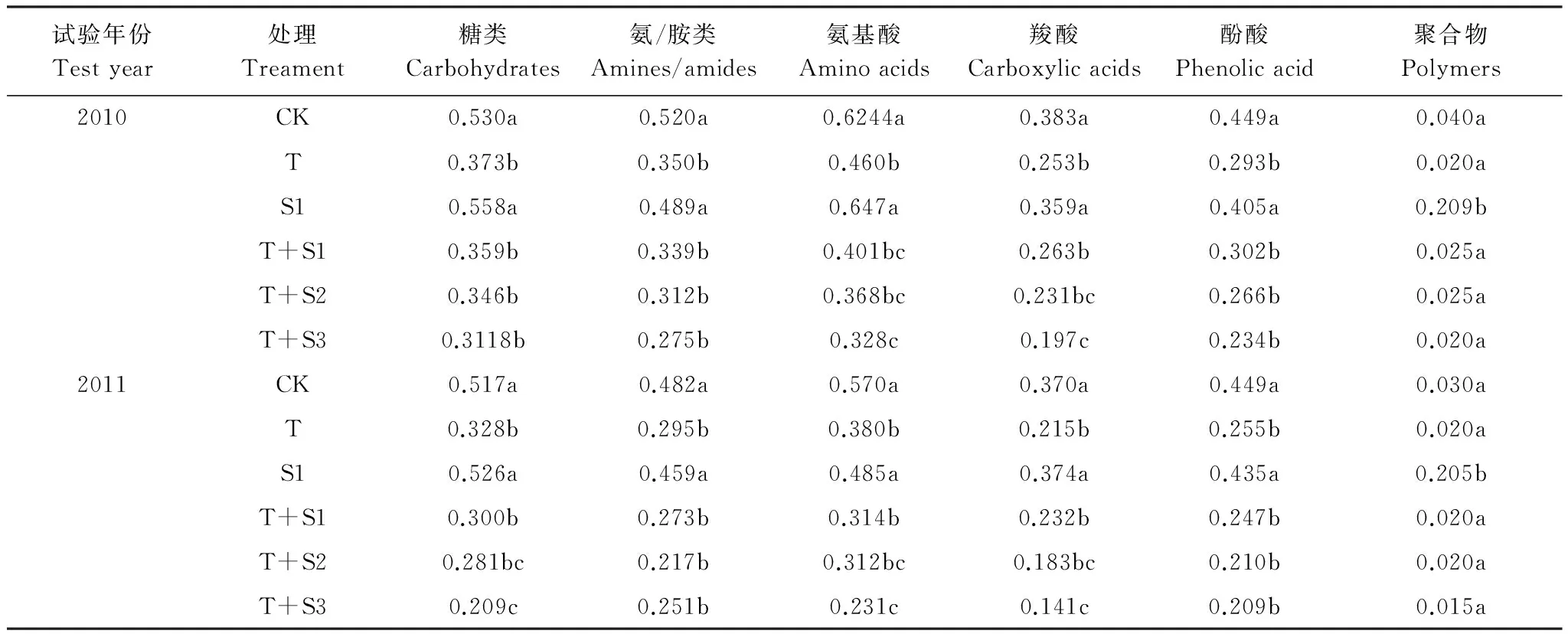
表 3 O3熏氣和太陽輻射減弱處理下土壤不同類別碳源AWCD值Table 3 AWCD of different types carbon sources under ozone fumigation and depressed solar irradiance
不同字母表示處理間經(jīng)Duncan氏多重極差檢驗(yàn)差異顯著(P<0.05)
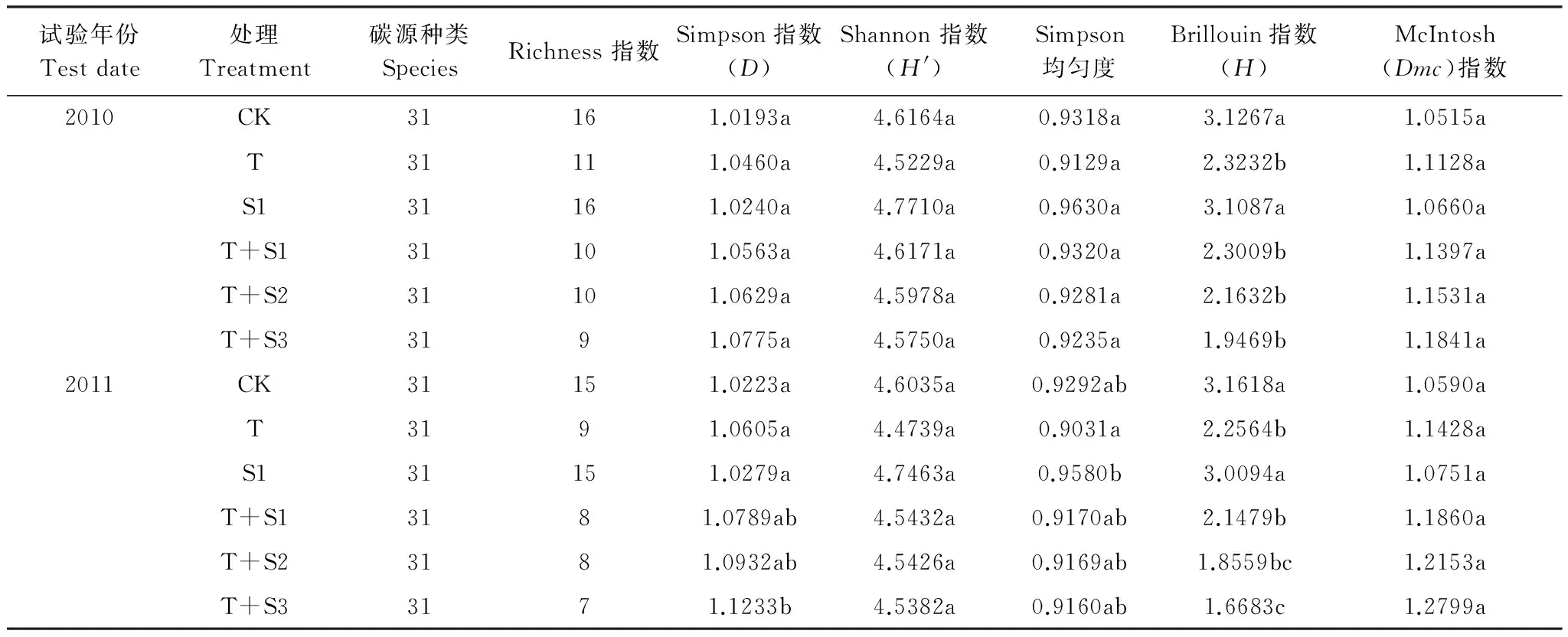
表 4 O3熏氣條件下太陽輻射減弱土壤微生物群落功能多樣性指數(shù)Table 4 Function diversity index of soil microbe community under ozone fumigation and depressed solar irradiance
2.4 O3熏氣和太陽輻射減弱條件下微生物群落功能主成分分析(PCA)
對(duì)培養(yǎng)96 h的AWCD數(shù)據(jù)進(jìn)行微生物群落功能主成分分析(PCA),土壤微生物碳源利用功能多樣性2010年相關(guān)的主成分1(PC1)、主成分2(PC2)依次解釋變量方差的70.24%、20.16%,PC1,PC2的累計(jì)貢獻(xiàn)率達(dá)到90.40%;2011年相關(guān)的主成分1、主成分2依次解釋變量方差的76.17%、12.80%,PC1,PC2的累計(jì)貢獻(xiàn)率達(dá)到88.97%;PC1、PC2是變異的主要來源,可以解釋變異的絕大部分信息(表5)。再將土壤碳利用主成分進(jìn)行分權(quán)計(jì)算,并計(jì)算出各因子在各主成分上的載荷:載荷=(特征向量×特征值1/2)2[31],得到表6,得出31 種碳源在兩個(gè)主成分上的載荷值。由表7可知,2010年,PC1主要綜合了15種碳源的信息,分別屬于糖類(5 種)、氨基酸(4 種)、羧酸(4 種)、聚合物(1 種)酚酸(1 種),說明有15 種碳源使土壤微生物群落代謝功能多樣性在PC1 上差異顯著,決定PC1 的分異。與PC2 顯著相關(guān)的碳源有6 種,分別屬于糖類(5 種)、胺類(1 種),說明有6 種碳源使土壤微生物群落代謝功能多樣性在PC2 上差異顯著,決定PC2的分異。2011年,PC1主要綜合了15 種碳源的信息,分別屬于糖類(6種)、氨基酸(4 種)、羧酸(3 種)、聚合物(1 種)酚酸(1 種),說明有15 種碳源使土壤微生物群落代謝功能多樣性在PC1上差異顯著,決定PC1的分異。與PC2顯著相關(guān)的碳源有3 種,分別屬于糖類(2 種)、胺類(1 種),說明有3 種碳源使土壤微生物群落代謝功能多樣性在PC2 上差異顯著。由于大氣O3濃度增加和太陽輻射減弱引起土壤中微生物環(huán)境出現(xiàn)差異造成15種碳源分解有顯著變化。
表5 O3熏氣和太陽輻射減弱條件下根際土壤微生物碳源主成分特征值
Table 5 Principal component eigenvalues of different carbon sources types by rhizosphere soil microbes under ozone fumigation and depressed solar irradiance (2010,2011)
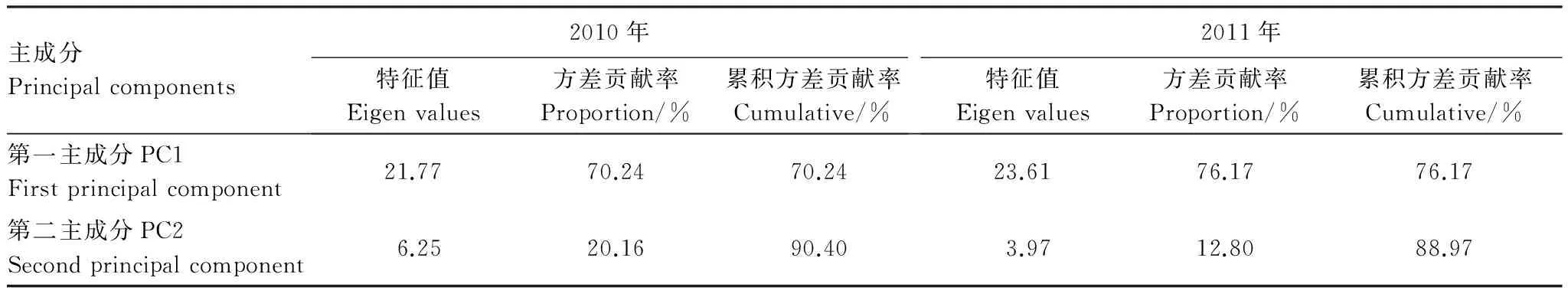
主成分Principalcomponents2010年特征值Eigenvalues方差貢獻(xiàn)率Proportion/%累積方差貢獻(xiàn)率Cumulative/%2011年特征值Eigenvalues方差貢獻(xiàn)率Proportion/%累積方差貢獻(xiàn)率Cumulative/%第一主成分PC1Firstprincipalcomponent21.7770.2470.2423.6176.1776.17第二主成分PC2Secondprincipalcomponent6.2520.1690.403.9712.8088.97
3 討論
復(fù)雜的大氣背景中,多種大氣成分相互影響,NASA Goddard報(bào)道,臭氧對(duì)硫酸化合物的濃度影響很大[32]。硫酸化合物是重要的負(fù)輻射強(qiáng)迫物質(zhì),造成到達(dá)地面的太陽輻射減少。單文坡等認(rèn)為,近地面O3濃度與太陽輻射和氣溫有很好的相關(guān)性[33],這主要是由于太陽輻射和氣溫是許多與O3相關(guān)的大氣化學(xué)或光化學(xué)過程的控制因素,而相對(duì)濕度可以直接影響大氣中自由基的數(shù)量,從而對(duì)O3化學(xué)產(chǎn)生影響。晴天地表各污染物濃度的變化比較平穩(wěn),陰天變化波動(dòng)較大,這也說明了太陽輻射在近地面大氣化學(xué)中的重要作用。O3熏氣能使太陽負(fù)輻射強(qiáng)迫增強(qiáng),增強(qiáng)了因太陽輻射減弱而給農(nóng)田生態(tài)帶來的負(fù)面影響。太陽輻射減弱促進(jìn)了植物對(duì)O3的化學(xué)吸收。兩年的定點(diǎn)試驗(yàn)得出,O3熏氣條件下與太陽輻射減弱復(fù)合作用均顯著降低了不同類型碳源的降解速率和降解總量,糖類、氨基酸類的分解變異度較高。輻射減弱與O3熏氣結(jié)合能加劇O3對(duì)碳源分解的抑制作用,O3與輻射減弱之間存在協(xié)同關(guān)系。
表6 O3熏氣和太陽輻射減弱條件下土壤微生物對(duì)不同碳源利用的主成分特征向量
Table 6 Principal component eigenvectors of different carbon sources types by rhizosphere soil microbes under ozone fumigation and depressed solar irradiance (2010,2011)
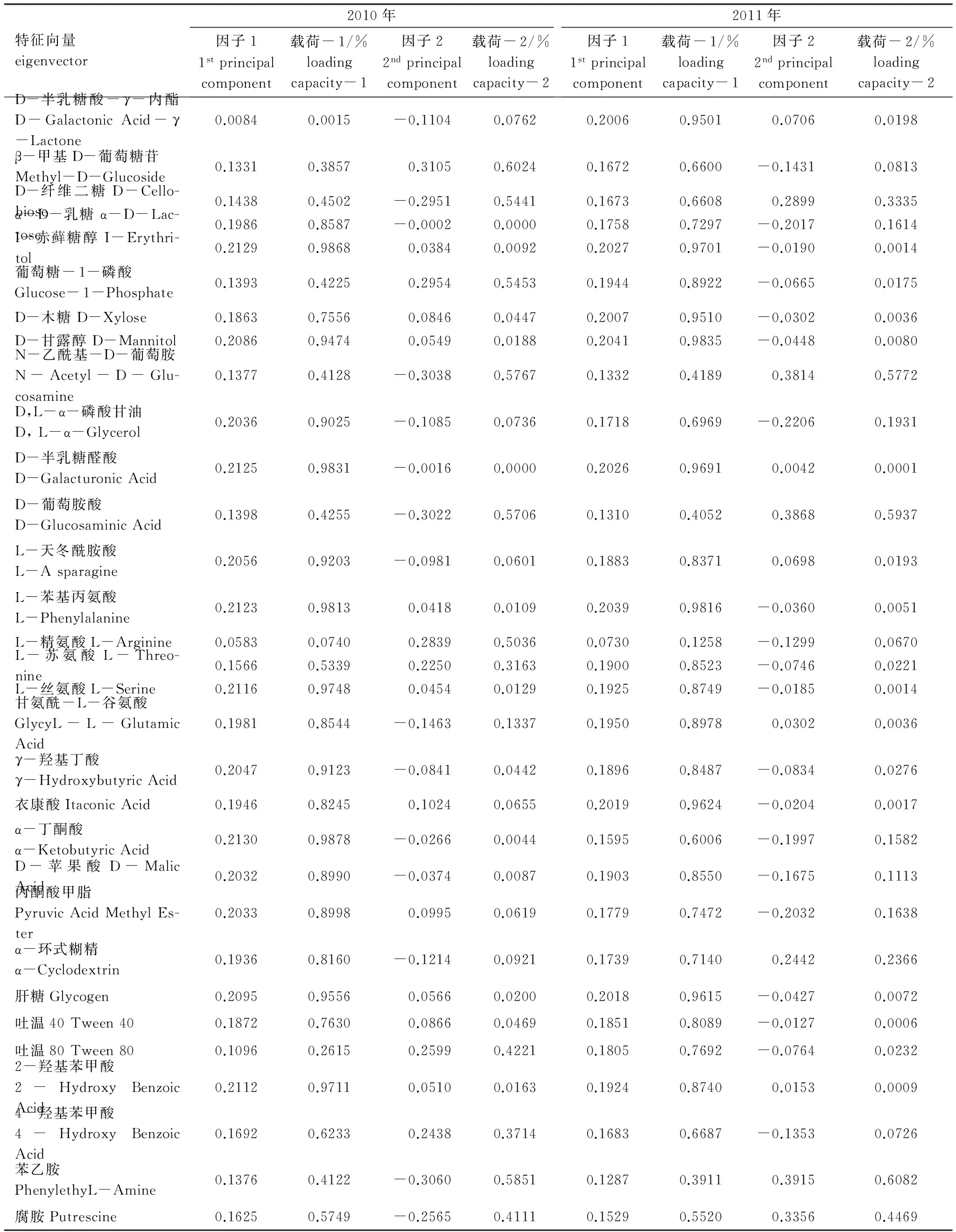
載荷- 1,2分別指各主成分上承載的各因子的方差百分率
土壤微生物對(duì)大氣環(huán)境變化的響應(yīng)受到土壤溫度、CO2、養(yǎng)分利用效率和水分狀況等因素的制約[34],O3濃度升高和太陽輻射減弱長期作用引起土壤養(yǎng)分有效性[35]和水分利用效率的變化將改變土壤微生物最初的響應(yīng)。O3與太陽輻射減弱的交互作用,在不同年份差異顯著性不同,這可能是土壤微生物異質(zhì)性的結(jié)果。由于長期環(huán)境變化對(duì)生態(tài)系統(tǒng)微生物過程及結(jié)果的預(yù)測(cè)受到有限的生態(tài)學(xué)信息的制約,很難確定在較長時(shí)間尺度上O3濃度升高和太陽輻射減弱對(duì)土壤微生物的作用,更不能進(jìn)行小尺度向大尺度的推繹,因?yàn)樵诓煌臅r(shí)間尺度土壤微生物對(duì)大氣環(huán)境變化的響應(yīng)可能很不一致。
大氣環(huán)境變化對(duì)植物根際土壤微生物多樣性的影響是通過植物生長代謝間接作用的結(jié)果,不僅隨著植物類型改變, 還隨著植物的生長發(fā)育過程改變, 具有非常強(qiáng)的時(shí)空特征[36]。土壤微生物多樣性對(duì)大氣環(huán)境的響應(yīng)與植物生長發(fā)育過程有關(guān)。太陽輻射減弱增加了植物對(duì)O3的吸收,加重了O3傷害,O3濃度升高與太陽輻射減弱的交互作用加劇了對(duì)土壤根際微生物活性的抑制。
表征微生物群落功能多樣性的指數(shù)沒有顯著變化,說明O3熏氣條件下與太陽輻射減弱在較短時(shí)期內(nèi)沒有改變土壤微生物的群落結(jié)構(gòu)和功能多樣性。要確定在較長時(shí)間尺度上O3濃度升高和太陽輻射減弱對(duì)土壤微生物多樣性的影響,需長期的后續(xù)研究。
4 結(jié)論
(1) O3熏氣和太陽輻射減弱條件下,土壤微生物代謝活性下降,不同類型碳源的分解速率、分解總量下降。
(2)輻射減弱20%的單因素作用對(duì)碳源的分解沒有顯著影響,但與O3結(jié)合則能加劇了O3對(duì)碳源分解的抑制作用,O3與遮蔭之間存在協(xié)同關(guān)系。O3濃度增加與太陽輻射減弱復(fù)合背景下,糖類、氨基酸類的分解變異度較高。
(3) O3熏氣和太陽輻射減弱條件下,表征微生物優(yōu)勢(shì)度、均勻度、豐富度的多樣性指數(shù)在兩年試驗(yàn)中沒有顯著變化,微生物群落結(jié)構(gòu)功能多樣性在短期內(nèi)沒有明顯影響。
[1] Tausz M, Grulke N E, Wieser G. Defense and avoidance of ozone under global change. Environmental Pollution, 2007, 147(3): 525- 531.
[2] Fuhrer J, Sk?rby L, Ashmore M R. Critical levels for ozone effects on vegetation in Europe. Environmental Pollution, 1997, 97(1/2): 91- 106.
[3] Forster P, Ramaswamy V, Artaxo P, Berntsen T, Betts R, Fahey D W, Haywood J, Lean J, Lowe D C, Myhre G, Nganga J, Prinn R, Raga G, Schulz M, Van Dorland R. Changes in atmospheric constituents and in radiative forcing // Solomon S, Qin D, Manning M, Chen Z, Marquis M, Averyt K B, Tignor M, Miller H L. Climate Change 2007: The Physical Science Basis. Working Group I Contribution to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, United Kingdom and New York: Cambridge University Press, 2007.
[4] Satheesh S K, Ramanathan V. Large differences in tropical aerosol forcing at the top of the atmosphere and earth′s surface. Nature, 2000, 405(6782): 60- 63.
[5] Aneja M K, Sharma S, Fleischmann F, Stich S, Heller W, Bahnweg G, Charles M J, Schloter M. Influence of ozone on litter quality and its subsequent effects on the initial structure of colonizing microbial communities. Microbial Ecology, 2007, 54(1): 151- 160.
[6] Gallo M, Amonette R, Lauber C, Sinsabaugh R L, Zak D R. Microbial community structure and oxidative enzyme activity in nitrogen-amended north temperate forest soils. Microbial Ecology, 2004, 48(2): 218- 229.
[7] Voidarou C, Tzora A, Skoufos I, Vassos D, Galogiannis G, Alexopoulos A., Bezirtzoglou E. Experimental effect of ozone upon some indicator bacteria for preservation of an ecologically protected watery system. Water, Air and Soil Pollution, 2007, 181(1/4): 161- 171.
[8] Johnson R M, Pregitzer K S. Concentration of sugars, phenolic acids, and amino acids in forest soils exposed to elevated atmospheric CO2and O3. Soil Biology and Biochemistry, 2007, 39(12): 3159- 3166.
[9] 李果梅, 王殳屹, 史奕, 陳欣. O3濃度升高及溫度對(duì)麥田土壤酶活性及酚酸類物質(zhì)含量的影響. 農(nóng)業(yè)環(huán)境科學(xué)學(xué)報(bào), 2008, 27(1): 121- 125.
[10] 劉賢趙, 康紹忠, 邵明安, 王力. 土壤水分與遮陰水平對(duì)棉花葉片光合特性的影響研究. 應(yīng)用生態(tài)學(xué)報(bào), 2000, 11(3): 377- 381.
[11] 楊渺, 毛凱, 馬金星. 遮陰生境下假儉草的形態(tài)變化與能量分配研究. 中國草地, 2004, 26(2): 44- 48,62.
[12] 王迅, 張新全, 劉金平. 草坪草對(duì)遮蔭脅迫的生理反應(yīng). 草業(yè)科學(xué), 2006, 23(4): 86- 90.
[13] 文軍. 遮蔭對(duì)粵西香根草光合特性和生長的影響 [D]. 蘭州: 甘肅農(nóng)業(yè)大學(xué), 2007.
[14] 韋海建, 楊惠敏, 趙亮. 遮蔭環(huán)境對(duì)白三葉草氣孔和光合特性的影響. 草業(yè)科學(xué), 2007, 24(10): 94- 97.
[15] 翟薇. 大氣氣溶膠輻射效應(yīng)對(duì)長江三角洲地區(qū)主要作物生產(chǎn)的影響 [D]. 北京: 中國氣象科學(xué)研究院, 2007.
[16] 蔡瑞國, 王振林, 張敏, 王曉英, 張?bào)w彬, 周印富, 王文頗, 李彥生. 挑旗-開花期遮光對(duì)強(qiáng)筋小麥干物質(zhì)積累的影響. 河北科技師范學(xué)院學(xué)報(bào), 2009, 23(1): 12- 19.
[17] Chen Z, Wang X K, Feng Z Z, Xiao Q, Duan X N. Impact of elevated O3on soil microbial community function under wheat crop. Water, Air, and Soil Pollution, 2009, 198(1/4): 189- 198.
[18] Bastida F, Zsolnay A, Hemández T, Garcia C. Past, present and future of soil quality indices: a biological perspective. Geoderma, 2008, 147(3/4): 159- 171.
[19] Lundquist E J, Jackson L E, Scow K M, Hsu C. Changes in microbial biomass and community composition, and soil carbon and nitrogen pools after incorporation of rye into three California agricultural soils. Soil Biology and Chemistry, 1999, 31(2): 221- 236.
[20] Zelles L. Fatty acid patterns of phospholipids and lipopolysaccharides in the characterisation of microbial communities in soil: a review. Biology and Fertility of Soils, 1999, 29(2): 111- 129.
[21] 楊永華, 姚健, 華曉梅. 農(nóng)藥污染對(duì)土壤微生物群落功能多樣性的影響. 微生物學(xué)雜志, 2000, 20(2): 23- 25, 47- 47.
[22] 徐華勤, 肖潤林, 鄒冬生, 宋同清, 羅文, 李盛華. 長期施肥對(duì)茶園土壤微生物群落功能多樣性的影響. 生態(tài)學(xué)報(bào), 2007, 27(8): 3355- 3361.
[23] 王加龍, 劉堅(jiān)真, 陳杖榴, 陳林. 恩諾沙星殘留對(duì)土壤微生物數(shù)量及群落功能多樣性的影響. 應(yīng)用與環(huán)境生物學(xué)報(bào), 2005, 11(1): 86- 89.
[24] 區(qū)余端, 蘇志堯, 彭桂香, 劉剛. 車八嶺山地常綠闊葉林冰災(zāi)后土壤微生物群落功能多樣性. 生態(tài)學(xué)報(bào), 2009, 29(11): 6156- 6164.
[25] Lou Y S, Ren L X, Li Z P, Cheng H Y, Zhang T L. Effect of elevated ultraviolet-B radiation on microbial biomass carbon and nitrogen in barley rhizosphere soil. Water, Air, and Soil Pollution, 2011, 219(1/4): 501- 506.
[26] 展小云, 吳冬秀, 張琳, 張燦娟, 周雙喜, 楊云霞. 小葉錦雞兒根際微生物群落功能多樣性對(duì)環(huán)境變化的響應(yīng). 生態(tài)學(xué)報(bào), 2010, 30(12): 3087- 3097.
[27] 張乃莉, 郭繼勛, 王曉宇, 馬克平. 土壤微生物對(duì)氣候變暖和大氣N沉降的響應(yīng). 植物生態(tài)學(xué)報(bào), 2007, 31(2): 252- 261.
[28] Zheng H, Ouyang Z Y, Fang Z G, Zhao T Q. Application of biolog to study on soil microbial community functional diversity. Acta Pedologica Sinica, 2004, 41(3): 456- 461.
[29] Zogg G P, Zak D R, Pregitzer K S, Bruton A J. Microbial immobilization and the retention of anthropogenic nitrate in a northern hardwood forest. Ecology, 2000, 81(7): 1858- 1866.
[30] Sparling G P. Soil microbial biomass, activity and nutrient cycling as indicators of soil health // Pankhurst C, Doube B M, Gupta V V S R. Biological Indicators of Soil Health. Wallingford, UK: CAB International, 1997: 97- 119.
[31] 邱莉萍, 劉軍, 王益權(quán), 孫慧敏, 和文祥. 土壤酶活性與土壤肥力的關(guān)系研究. 植物營養(yǎng)與肥料學(xué)報(bào), 2004, 10(3): 277- 280.
[32] 張燕. 臭氧和硫化物對(duì)地球氣候變化的影響. 科學(xué)與管理, 2006, (5): 94- 94.
[33] 單文坡. 大氣臭氧濃度變化規(guī)律及相關(guān)影響因素研究 [D]. 濟(jì)南: 山東大學(xué), 2006.
[34] Tscherko D, Kandeler E, Jones T H. Effect of temperature on below-ground N-dynamics in a weedy model ecosystem at ambient and elevated atmospheric CO2levels. Soil Biology and Biochemistry, 2001, 33(4/5): 491- 501.
[35] 吳芳芳, 鄭有飛, 吳榮軍, 李萍, 王錦旗. 4年O3熏氣對(duì)小麥根際土壤氮素微生物轉(zhuǎn)化的影響. 生態(tài)學(xué)報(bào), 2013, 33(24): 7679- 7689.
[36] 吳芳芳, 鄭有飛, 吳榮軍, 王錦旗. 近地層臭氧對(duì)小麥抗氧化酶活性變化動(dòng)態(tài)的影響. 生態(tài)學(xué)報(bào), 2011, 31(14): 4019- 4026.
functional diversity in winter wheat rhizosphere
WU Fangfang, ZHENG Youfei*, WU Rongjun, WANG Jinqi, LI Ping
JiangsuKeyLaboratoryofAtmosphericEnvironmentalMonitoringandPollutionControlHigh-techResearch,NanjingUniversityofInformationScience&Technology,Nanjing210044,China
Enhanced ozone concentration and depressed solar irradiation were simulated by using open-top chambers (OTCs) method and shading net techniques at 100 nL/L ozone fumigation assembling with three different levels of depressed solar irradiance. The effect of enhanced O3combination with depressed solar irradiance was also studied. Two experiments were performed over two years. The combination of three OTCs with six shaded chambers was set up. Each OTC fed with 100 nL/L ozone. In the shaded chambers, solar irradiance was reduced to different levels of 20%, 40%, 60% respectively. Combined actions of O3and depressed solar irradiance were tested by shading net above OTCs. Using unshaded OTC and unfed with O3as controls. OTC fed with 100 nL/L ozone only, and shaded chamber solar irradiance reduced 20% only were applied too. Each chamber was equivalent and repeated three times. Winter wheat was grown to all life stage in OTCs, shading net and their combine chambers. From reviving to mature stages, in OTCs the plants were treated for 8 h (8:00—16:00) every sunny day with O3, in shading chambers covered net all life stage. The ability of winter wheat (Triticumaestivum)rhizosphere microbial to use different carbon sources were measured by applying BIOLOG ECO MicroPlate and Average WelL-Color Development (AWCD). Microbial diversity index and Principal Component Analysis (PCA) were calculated on the utilization of different carbon sources. Each treatment was assayed in triplicate. The data were initially compared by analysis of variance and differences between means were detected using the Duncans Multiple Range Tests. Values ofP<0.05 indicated significance. Quite similar results were obtained during the two years′ experiments. The results suggesting that under ozone fumigation and reduced solar irradiance, AWCD was reduced, catabolic ability of single carbon source was declined too. No obvious influence of O3combined with shading on soil microbial diversity was found. Principal Component Analysis suggested that different forms of carbon source have different sensitivity. The variation degree of carbohydrates′ is higher than others. Under ozone fumigation combined with reduced solar irradiance, AWCD and catabolic ability of single carbon sources were lower than enhanced O3or lower solar irradiance single acting. For soil microbial, two years′ results showed that combination effects of ozone fumigation and reduced solar irradiation reduced both rate and total utilization of carbon sources except polymers. However, there was no direct impact on the diversity of soil microbial. For Inhibitory effect on the degradation of carbon, combined effect of enhanced ozone concentration and reduced solar irradiation was greater than each factor functioning alone. 20% reduction of solar radiation increased the decomposition of polymers to some extent. On the conditions of combined O3fumigation and solar radiation weakened, that makes the metabolism of carbohydrates and amines highly variable, as they are largely affected by the environment. Enhanced O3and lower solar irradiation have a cooperative action. Reduced solar irradiance is benefit for winter wheat absorbing O3,resulting in a serious injury.
ozone; depressed solar irradiance; recombination action;Triticumaestivum; soil microbial diversity
教育部高等學(xué)校博士學(xué)科點(diǎn)專項(xiàng)科研基金(20123228110003); 國家自然科學(xué)基金(41075114)
2013- 08- 05;
2014- 10- 13
10.5846/stxb201308052029
*通訊作者Corresponding author.E-mail: zhengyf@nuist.edu.cn
吳芳芳, 鄭有飛, 吳榮軍, 王錦旗, 李萍.O3濃度升高和太陽輻射減弱對(duì)小麥根際土壤微生物功能多樣性的影響.生態(tài)學(xué)報(bào),2015,35(12):3949- 3958.
Wu F F, Zheng Y F, Wu R J, Wang J Q, Li P.Effects of ozone fumigation and depressed solar irradiance on soil microbial functional diversity in winter wheat rhizosphere.Acta Ecologica Sinica,2015,35(12):3949- 3958.
猜你喜歡
軍事文摘(2023年10期)2023-06-09 09:15:06
中學(xué)生數(shù)理化·八年級(jí)物理人教版(2022年3期)2022-03-16 05:55:08
空間科學(xué)學(xué)報(bào)(2021年4期)2021-08-30 08:31:18
當(dāng)代陜西(2021年2期)2021-03-29 07:41:24
媽媽寶寶(2017年3期)2017-02-21 01:22:28
河北書畫研究(2016年2期)2016-08-24 02:14:50
新農(nóng)業(yè)(2016年18期)2016-08-16 03:28:27
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(shù)(2016年3期)2016-03-26 07:13:38
