天山林區六種灌木生物量的建模及其器官分配的適應性
2015-02-11 02:06:27常順利張毓濤王文棟王慧杰
生態學報 2015年23期
仇 瑤, 常順利,*,張毓濤,王文棟,何 平,王慧杰,謝 錦
1 新疆大學資源與環境科學學院綠洲生態教育部重點實驗室,烏魯木齊 830046 2 新疆林科院森林生態研究所,烏魯木齊 830063
天山林區六種灌木生物量的建模及其器官分配的適應性
仇 瑤1, 常順利1,*,張毓濤2,王文棟2,何 平1,王慧杰1,謝 錦1
1 新疆大學資源與環境科學學院綠洲生態教育部重點實驗室,烏魯木齊 830046 2 新疆林科院森林生態研究所,烏魯木齊 830063
灌木全株生物量估算模型的構建仍存在一定困難,對灌木生物量在器官分配上所體現的適應性研究也不夠充分。以天山林區6種常見灌木為研究對象,在天山的東段、中段、西段林區分別設置樣地進行群落調查,由此以全株收獲法取得6種常見灌木若干標準株的全株、根、枝、葉及各徑級根的生物量,將D2H(地徑平方與高度的乘積)與V(冠幅面積與高度的乘積)分別選為估測模型的自變量,通過回歸分析法建立了各種灌木全株生物量的最優估算模型,然后比較了此6種灌木全株生物量在營養器官上分配差異以及根系生物量在徑級上的分配差異。結果表明:(1)天山林區6種常見灌木中,小檗(BerberisheteropodaSchrenk)、忍冬(LonicerahispidaPall. ex Roem. et Schuet.)、栒子(CotoneastermelanocarpusLodd.)的全株生物量約為8.48—9.01 kg,薔薇(RosaspinosissimaL.)、繡線菊(SpiraeahypericifoliaL.)、方枝柏(JuniperuspseudosabinaFisch. et Mey.)的全株生物量約為2.71—3.20kg;(2)薔薇、繡線菊、栒子的全株生物量最優估測模型是以V為自變量的函數,小檗、忍冬、方枝柏的全株生物量最優估測模型是以D2H為自變量的函數,各模型R2值均在0.850以上,且在P<0.05水平上達到顯著,模型模擬結果達到了較高的準確度;(3)6種灌木全株生物量在根、枝上的分配比重差異不顯著,僅在葉上的分配比重有差異(P<0.05);根系生物量在徑級上的分配均呈現隨根系徑級下降而減少的規律,6種灌木在徑級大于2 mm根上的分配比重存在差異(P<0.05,徑級大于20mm根為P<0.01水平);(4)6種灌木全株生物量在營養器官上的分配差異以及根系生物量在徑級上的分配差異均體現了各物種對其生境選擇的適應策略。
生物量建模;根系分級;適應策略;灌木;天山
灌木層是森林中的重要片層,灌木生物量是群落的重要數量特征之一[1-3],對灌木生物量的關注有助于群落的物質循環、消耗、分配、轉化、積累等研究工作的深入開展[4-5]。在林業傳統上,對喬木生物量和蓄積量的研究較為普遍,但對灌木生物量的關注較為欠缺。目前,測定灌木生物量的方法仍然主要是全株收獲法[6-7],但該法費時耗力且取樣數量又很難滿足統計學大樣本的要求,存在較大的不確定性。因此,采用易測因子來估算灌木生物量更為可行,成為當前學界較為主流的工作思路。
早期進行的灌木生物量研究方法主要有樣方法、平均木法、相對生長法等,隨著人們對各種灌木生物量研究的深入,多數研究人員傾向于使用回歸模型和數量化的方法[8-10]。以易測因子作為自變量,以生物量為因變量,通過數理統計的回歸分析方法,建立起這些因子之間的相對生長方程,以此來推算、預測整體的生物量[4]。對灌木生物量進行建模能夠在非破壞性、減小工作量基礎上提供較為簡單且準確的估算方法[11],使得對林區灌木生物量的連續清查成為可能。
植株生物量在各器官中的分配策略是植物生活史理論研究的主要內容[12-13]。植物在生長發育的過程中,通過不斷優化自身的資源分配來提高其適應環境變化的能力[14],故生物量在根、枝、葉之間的分配是植物對環境長期適應的結果[12,15],在很大程度上反映了植物的生態適應對策。當前,對生物量在植株各器官上分配的研究多集中在對不同徑級根系的生物量及生理功能的比較上[16-20],對其分配策略的生態適應性研究尚不充分,對灌木生物量的器官分配及其反映出的生態適應性問題更是鮮有報道。
本研究以天山林區6種常見灌木為研究對象,為排除水熱空間分布梯度因素的影響,在天山東部、中部、西部分別設置灌木樣地進行群落調查,在群落調查基礎上取該6種灌木的標準株若干,以全株收獲法求算全株生物量和各器官生物量,以實測數據構建天山林區6種灌木的單株生物量估算模型,并深入分析該6種灌木單株生物量在營養器官上的分配規律,探討各物種在全株生物量的營養器官分配方面以及根系生物量的徑級分配方面所體現出的生態適應性,對于更深入了解天山林區灌木的生物學特性具有一定意義。
1 材料與方法
1.1 研究區概況
天山林區以天山云杉(PiceaschrenkianaFisch. et Mey.)純林為主,林緣、林窗及林下的灌木主要有栒子(CotoneastermelanocarpusLodd.)、小檗(BerberisheteropodaSchrenk及B.nummulariaBge.)、薔薇(RosaspinosissimaL.)、繡線菊(SpiraeahypericifoliaL.)、方枝柏(JuniperuspseudosabinaFisch. et Mey.)、錦雞兒(CaraganaleucophloeaPojark.)、忍冬(LonicerahispidaPall. ex Roem. et Schuet.)等,林下草本植物主要有老鸛草(GeraniumrotundifoliumL.)、羽衣草(AlchemillatianschanicaJuz.)、羊角芹(AegopodiumpodagrariaL.)等。林下土壤均為山地灰褐色森林土[21]。
根據云杉森林長勢、地形地貌、氣候等條件的不同,把整個天山北坡從東到西分為東天山,中天山和西天山三段分別進行群落調查和采樣。
天山東段以哈密林場為調查采樣區,主要依托巴里坤縣白石頭景區(93°37′—93°41′E,N43°9′—43°13′)開展工作,該地屬溫帶大陸性干旱半干旱氣候,夏季炎熱、冬季寒冷、四季分明、干燥少雨,年平均氣溫為4.7 ℃,1月份極端最低氣溫為-42.6 ℃,7月份極端最高氣溫43 ℃;年平均降水量為176 mm,蒸發量2141 mm,無霜期平均156 d,年日照時數2280—3230h。
天山中段以烏魯木齊縣板房溝林場為調查采樣區,主要依托中國森林生態系統研究網絡(CFERN)天山森林生態系統定位站研究站(87°07′—87°28′E,43°14′—43°26′N)開展工作,該地屬溫帶大陸性氣候,年總輻射量達5.85×105 J cm-2a-1),年均氣溫約為2—3 ℃,歷年極端最高溫為30.5 ℃,極端最低溫為-30.2 ℃;年降水量400—600mm,最大積雪深度為65 cm。
天山西段以昭蘇林場為調查采樣區,主要依托昭蘇縣阿克蘇鄉(80°48′—80°57′E,42°41—42°44′N)開展工作,該地屬溫帶大陸性氣候,年平均溫度5—7 ℃,1月平均氣溫為-10℃,7月平均氣溫為22.6 ℃;年平均降水量為600—800mm,全年60%的降水量集中在5—8月,年平均蒸發量1100—1200mm,年平均相對濕度70%以上,無霜期120d。
從東至西,天山林區呈現出較為明顯的水分增多趨勢和較不明顯的熱量減少趨勢。
1.2 群落調查與全株生物量收獲
2013年7—8月,在天山東段哈密林場、中段板房溝林場、西段昭蘇林場分別設置栒子、小檗、薔薇、繡線菊、方枝柏、忍冬等6種代表性灌木的樣地,每種灌木在每個調查采樣區至少做1個10m×10m的調查樣方。在樣方調查數據的基礎上,以地徑(D, cm)、株高(H, m)、冠幅(CA, m2)為指標,分別對應選取6種灌木的標準株。每個物種在3個調查采樣區各全株采樣1—2個標準株,共5—6株。樣方設置與標準株采樣情況見表1。
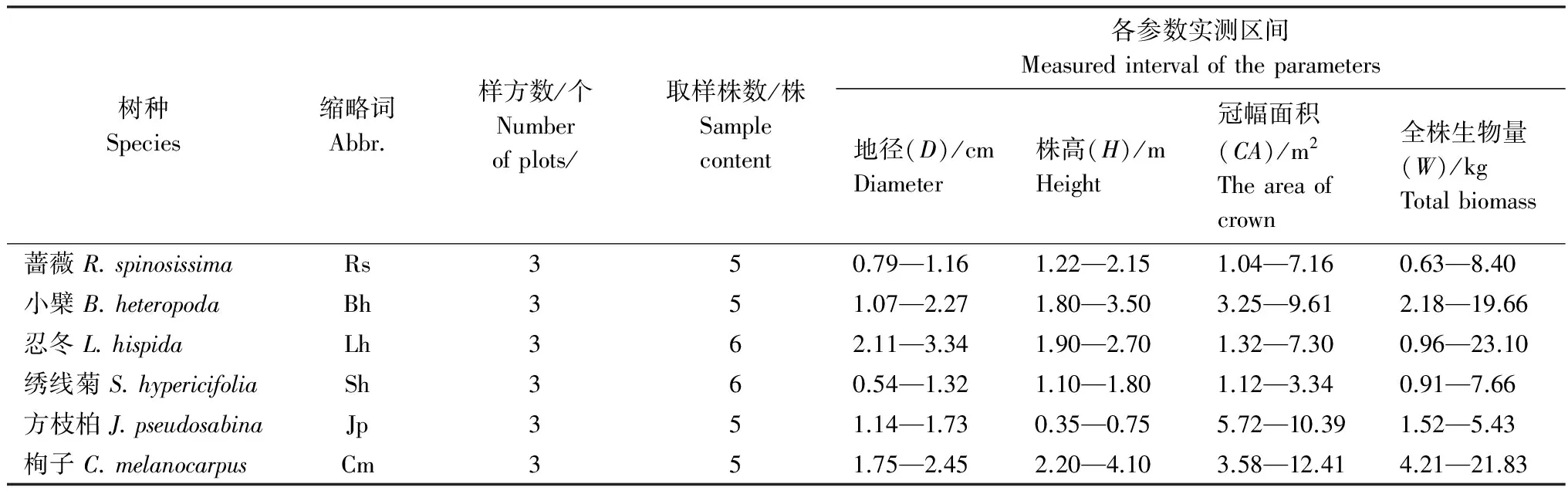
表1 樣方調查與標準株取樣統計表Table 1 The results of sample plots survey
用全收獲法將各標準株整株挖出,深度為根系分布所達范圍。全株分為根、枝、葉3個營養器官,其中又按徑級將根分為根樁(>20mm)、粗根(10—20mm)、中根(5—10mm)、小根(2—5 mm)和細根(<2 mm)等5個層級。分別稱取每個采樣株各營養器官以及各徑級根的鮮質量,再按比例稱取各組份樣品帶回實驗室,于105 ℃條件下烘干至恒重稱重。通過各采樣株各器官的鮮重、干重、含水率,計算各采樣株灌木全株生物量以及各器官的干物質量。
1.3 生物量建模方法
根據前人的研究,本研究先選取變量地徑(D,cm)、株高(H,cm)、冠幅面積(CA=π×C1×Cw/4,m2)、地徑平方與高度的乘積(D2H,cm3)、植株體積(V=CA×H,m3)、全株生物量(W,kg)等6個因素進行相關分析,篩選出與全株生物量相關性較好的狀態因子,再以相關性較好的幾個狀態因子(地徑(D)、高度(H)、冠幅面積(CA=π×C1×Cw/4)、植株體積(V=CA×H)、D2H)為自變量,全株生物量(W)為因變量進行回歸分析,最后選用以下幾種生物量研究中常用的回歸模型(見式1至式7)建立預測林下灌木生物量的回歸模型:
y=b0+b1x
(1)
y=b0+b1x+b2x2
(2)
y=b0(b1)x
(3)
y=b0+b1lnx
(4)
y=b0+b1x+b2x2+b3x3
(5)
y=b0e(b1x)
(6)
y=b0xb1
(7)
通過判定系數(R2)和標準誤(SEE)的大小及回歸檢驗顯著水平來評價每個物種的各方程的優劣,從中選出擬合度最好、相關最密切的數學模型作為該種灌木全株生物量的估算模型。
1.4 數據分析方法
本研究中的數據采用Excel進行數據統計,并用SPSS17.0統計軟件進行相關分析、回歸分析和多重比較(LSD)。
2 結果與分析
2.1 6種灌木的生物量模型構建
2.1.1 生物量回歸方程自變量的選擇
對樣地內選取的各灌木全株生物量(W)與其地徑(D)、高度(H)、冠幅面積(CA)等因子之間進行相關分析(表2)。
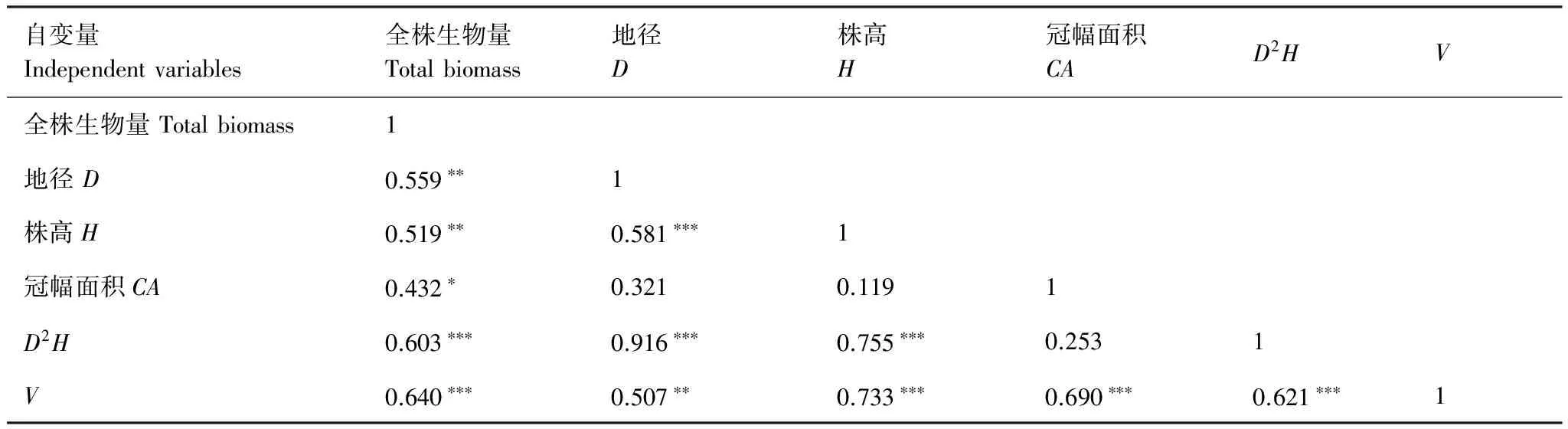
表2 6種灌木重要變量間的相關分析Table 2 The correlation analysis among independent variables for six shrubs
D:地徑 Diameter;H:株高 Height;CA:冠幅面積The area of crown(CA=π×C1×Cw/4)C1:crown major axis;Cw:crown minor axis;D2H:地徑平方與高度的乘積 Square of diameter(D2) multiplied by height(H);V:植株體積 The volume of shrub(V=CA×H); *P<0.05;**P<0.01;***P<0.001
表2可知,各個變量與全株生物量之間都有較強的相關關系,其中,包含地徑平方與高度的乘積(D2H)、植株體積(V=CA×H)與全株生物量的相關系數最大,相關性在P<0.001水平上達到顯著。因此,建立天山林區6種灌木生物量回歸模型時,分別選用D2H、V作為自變量。
2.1.2 生物量模型的確立
為了提高建立的回歸方程的準確度,分別用V和D2H兩個自變量與幾個常用的生物量回歸方程進行擬合,通過篩選判定系數(R2)較高、標準誤(SEE)較小以及通過回歸檢驗的方程,找到擬合度最好的方程,結果見表3。
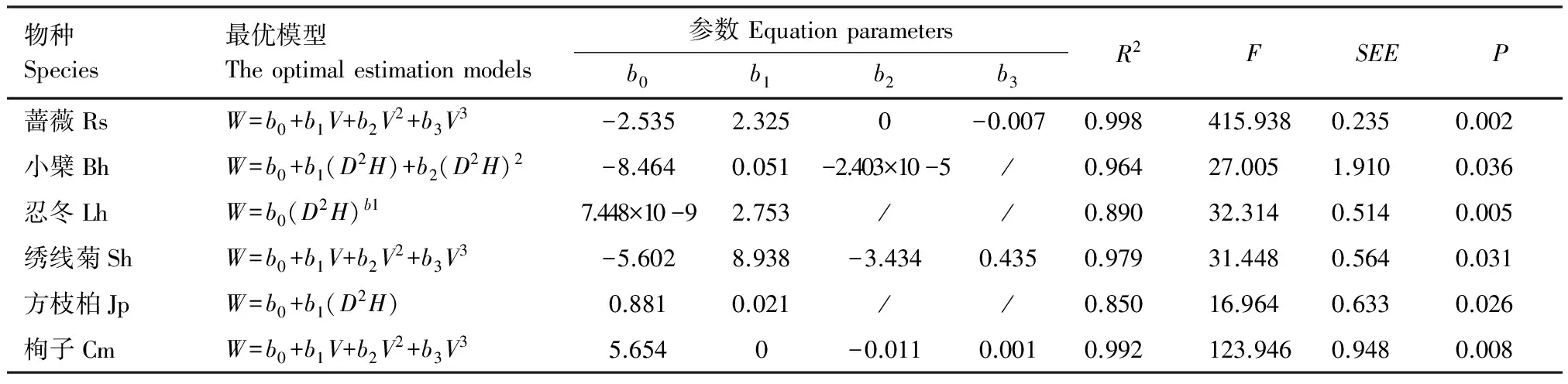
表3 6種灌木全株生物量的最優估算模型Table 3 The optimal estimation models for total biomass of six shrub species
薔薇Rs:R.spinosissima;小檗Bh:B.heteropoda;忍冬Lh:L.hispida;繡線菊Sh:S.hypericifolia;方枝柏Jp:J.pseudosabina;栒子Cm:C.melanocarpus;R2為判別系數;F為方差分析的F值the value of ANOVA;SEE為估計值的標準誤差the standard error of estimate;P為顯著性水平
薔薇、繡線菊和栒子的全株生物量與V相關性最好,最優模型為y=b0+b1V+b2V2+b3V3,R2分別為0.998、0.979 和 0.992。小檗、忍冬和方枝柏與D2H相關性最好,小檗的最優模型為y=b0+b1(D2H)+b2(D2H)2,R2為0.964。忍冬的最優模型為y=b0(D2H)b1,R2為0.890。方枝柏的最優模型為y=b0+b1(D2H),R2為0.850。薔薇、忍冬和栒子在P<0.01水平上達到顯著,小檗、繡線菊和方枝柏在P<0.05水平上達到顯著。天山林區6個灌木物種的全株生物量估算模型結果見式8—式13。
薔薇Rs:
W=-2.535+2.325V-0.007V3
(8)
繡線菊Sh:
W=-5.602+8.938V-3.434V2+0.435V3
(9)
栒子Cm:
W=5.654-0.011V2+0.001V3
(10)
小檗Bh:
W=-8.464+0.051(D2H)-(2.403×10-5)(D2H)2
(11)
忍冬Lh:
W=(7.448-9)(D2H)2.753
(12)
方枝柏Jp:
W=0.881+0.021(D2H)
(13)
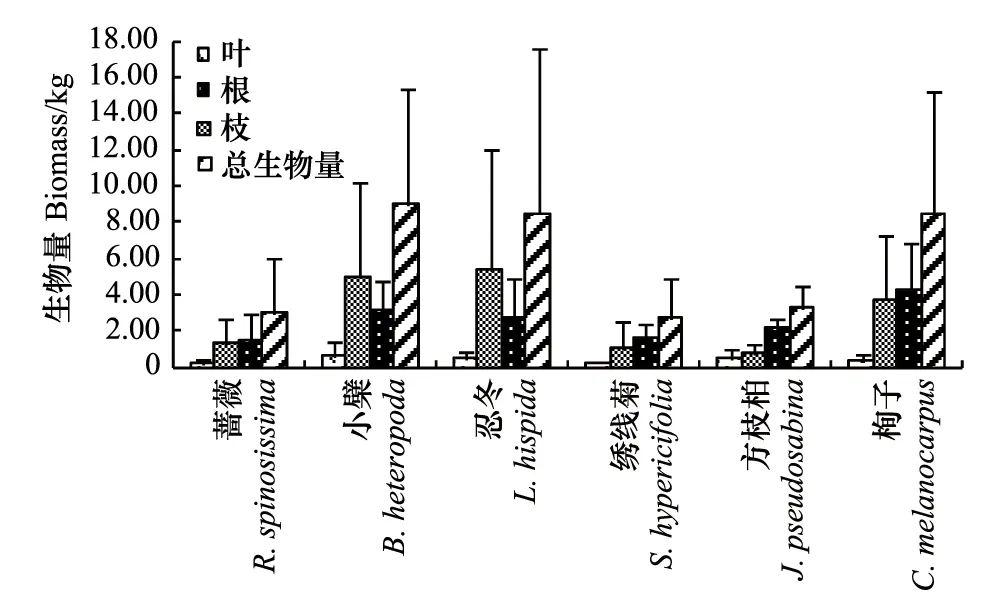
圖1 6種灌木全株及營養器官的生物量 Fig.1 The biomass of leaf, branch, root and total biomass of six shrubs
2.2 6種灌木全株生物量的基本情況
對6種灌木標準木的全株生物量和根、枝、葉生物量分別在多個重復中求均值進行比較分析,結果見圖1。圖1表明,6種灌木中,小檗、忍冬、栒子的全株生物量較大,在8.48—9.01 kg之間;薔薇、繡線菊、方枝柏全株生物量較小,在2.71—3.20kg之間。6種灌木的根生物量在1.38—4.20kg之間;枝生物量在0.70—5.37 kg之間,其中小檗、忍冬的略大為5.00kg和5.37 kg,薔薇、繡線菊、方枝柏、栒子略小,在0.70—3.73 kg之間;葉生物量均在0.70kg以內。
2.3 6種灌木全株生物量在營養器官上的分配
對6種灌木生物量在營養器官根、枝、葉間的分配進行比較分析,進行物種間的差異比較,得到圖2。圖2表明,6種灌木在營養器官根、枝、葉間的生物量分配呈現出大體相同的趨勢,但同一器官在不同灌木全株生物量中所占比例不同。
圖2可見,各營養器官生物量分配總體上表現為根生物量>枝生物量>葉生物量。6種灌木根、枝、葉生物量占全株生物量的比例各不相同,根生物量所占比例在47.8%—72.9%之間,物種間差異不顯著;枝條生物量所占比例在20.6%—46.5%之間,物種間差異也不顯著;而葉生物量所占比例在2.4%—10.1%之間,其中方枝柏為10.1%、栒子為3.5%、繡線菊為2.4%,僅呈現出方枝柏與繡線菊、栒子有顯著性差異(P<0.05),而其他物種間差異不顯著。即,6種灌木間生物量在根、枝上的分配差異不顯著,僅在葉上的分配有差異。
2.4 6種灌木根系生物量在徑級上的分配
對6種灌木生物量在根系不同徑級間的分配進行比較分析,進行物種間的差異比較,得到圖3。圖3表明,6種灌木在根系不同徑級間的生物量分配呈現出大體相同的趨勢,但同一徑級的根在不同灌木總根系中生物量所占比例不同。
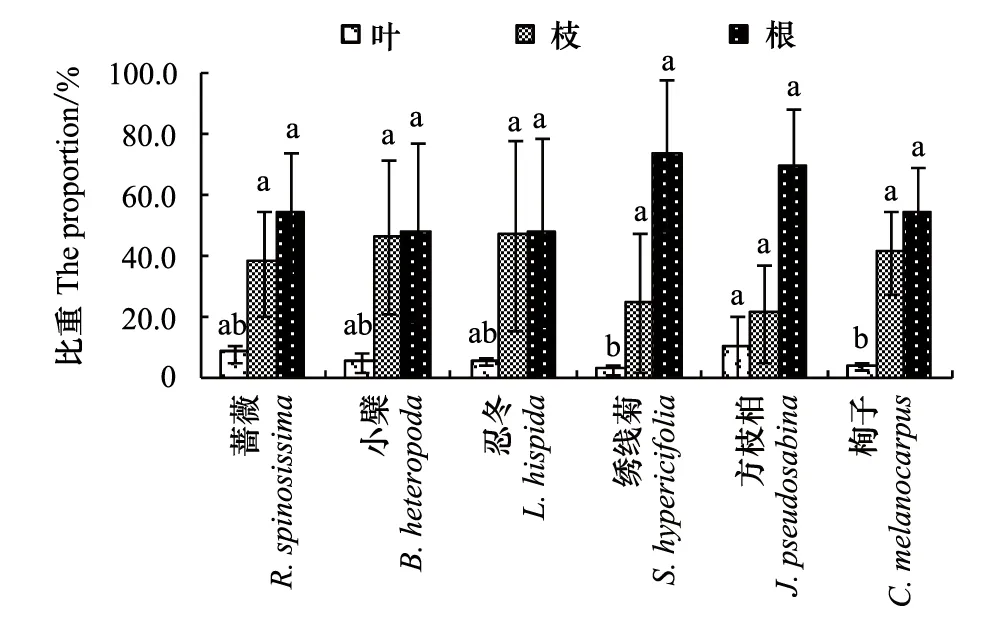
圖2 6種灌木全株生物量在各營養器官中的分配Fig.2 The allocation of six shrubs` total biomass in vegetative organs
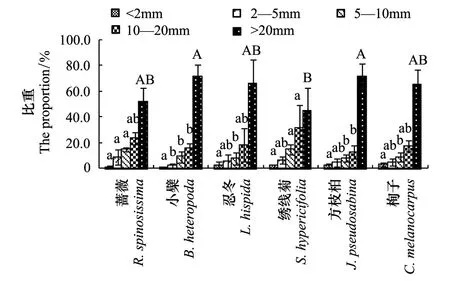
圖3 6種灌木根系生物量在徑級上的分配Fig.3 The allocation of six shrubs` underground biomass in organs小寫字母a、b表示統計量的差異在 P<0.05水平上;大寫字母A、B表示統計量的差異在P <0.01的水平上
圖3可見,灌木根系各不同徑級生物量分配大小均表現為根樁(>20mm)>粗根(10—20mm)>中根(5—10mm)>小根(2—5 mm)>細根(<2 mm)。通過方差分析和LSD多重比較可以得出,6種灌木間根系生物量在各徑級根上的分配有所區別。
在根樁(>20mm)生物量的分配上,方枝柏最大為71.6%、繡線菊最小為44.9%,方枝柏、小檗與繡線菊有極顯著性差異(P<0.01),而其他物種間均無顯著性差異。
在粗根(10—20mm)生物量的分配上,繡線菊最大為31.2%、方枝柏最小為13.1%,繡線菊與小檗、方枝柏間差異顯著(P<0.05),而其他物種間均無顯著性差異。
在中根(5—10mm)生物量的分配上,薔薇和繡線菊最大均為14.9%、忍冬最小為8.0%,薔薇、繡線菊與其他4種灌木間均有顯著差異(P<0.05)。
在小根(2—5 mm)生物量的分配上,薔薇最大為8.9%、小檗最小為2.5%,薔薇與小檗間有顯著差異(P<0.05),其他物種間均無顯著性差異。
在細根(<2 mm)生物量的分配上,6種灌木均不超過3.3%,且6種灌木間無顯著性差異。
3 結論與討論
3.1 關于灌木生物量模型構建的探討
本研究分別在天山東段、中段和西段各自設立林下灌木樣地,對天山云杉森林林區常見的6種林下灌木進行全株生物量的取樣,利用回歸分析的方法建立了這6種灌木全株生物量的最優估測模型。
在生物量估算建模中,回歸模型自變量的選取是難點所在[1,5-6,22-26]。本研究通過對地徑(D)、株高(H)、冠幅面積(CA)、D2H、V等易測因子與全株生物量進行相關分析發現,D2H、V與全株生物量相關性最大,均在P<0.001水平上達到顯著,與前人同類的研究較為一致[6]。這表明利用二因子復合變量比單因子變量進行模型擬合的效果要好,故本文分別用D2H和V對6種灌木進行回歸模型的擬合,從中篩選出每種灌木的最優模型。
從模型模擬的結果來看,每種灌木選取的自變量和最優方程并不完全相同,這是因為不同種灌木的形態是不同的,而生物量估測參數的選擇與灌木形態關系密切相關[27],因此本文不能用同一種變量對6種不同灌木去構建形式上相對統一的方程。
有些灌木的生長形態類似于圓柱型或圓錐形,以D2H為自變量進行模型的模擬有較高的準確性。而有些較為矮小的多分枝叢生灌木,以橫向發展冠幅來增加光合作用面積[28],故以V為自變量來估算生物量具有較高的精度[24]。本研究中灌木生物量最優模型形式多數為三次方程W=b0+b1x+b2x2+b3x3,這與前人同類工作較為一致[6]。另外,有些前人同類工作也采用W=b0(X)b1模型對生物量進行描述[22-23,29-32],本研究中只有忍冬的生物量最優模型為W=b0(D2H)b1。
雖然本研究限于條件對每種灌木的取樣仍然偏少,但在實測數據的基礎上,6種灌木模型模擬的R2值均在0.850以上,在P<0.05水平上達到顯著。因此,仍然可以認為模擬結果達到了較高的準確度。
林下灌木的生長狀況會因林分、土壤等立地條件存在一定的差異,可能會出現同物種在不同立地條件下估算模型不同的情況。雖然天山林區從東至西呈現出較為明顯的水分增多和較不明顯的熱量減少趨勢,且本研究在天山東、中、西段分別得到了6種灌木的全株生物量,各有5—6個重復,但若在東、中、西三段天山間進行6種灌木單株生物量及其各自估算模型的比較的話,則在每段研究區的取樣重復數都是不足的(表1)。若開展此分析,工作量將繼續成倍增加。因此,本文是將每種灌木在三段天山的取樣均作為重復,分別構建了6種灌木在整個天山林區統一的生物量估測模型。在天山林區以外,應用本文的6種灌木生物量估算模型時,需進行進一步驗證,當存在冠幅、植株高度變量等的外推時也需做進一步的驗證。
3.2 天山林區灌木全株生物量在營養器官上分配策略的適應性分析
在植物的整個生長發育過程中,可利用的光合固定產物是有限的,因此植物必需對有限的資源進行分配,在獲取光合固定產物的地上部分的器官和吸收土壤中水分和養分的地下部分的器官之間進行權衡[33]。
單株生物量在營養器官上分配格局的不同,反映了不同物種的對其生境的適應策略。最優分配理論認為,植物通過調節生物量在各器官中的分配來適應外界的環境條件,更多的生物量總是分配給那些獲取最受限制資源的器官[12,34]。當光照資源成為最主要的限制因子時,植物將更多的生物量分配給地上器官(枝、葉)以盡可能多的獲取光照;當水分或養分成為最主要的限制因子時,植物將更多的生物量分配給地下器官(根系)以盡可能多的獲取土壤水分或養分[35]。
從總體來看,天山林區6種代表性灌木全株生物量均呈現出根>枝>葉的規律,與前人研究結果一致[36-37],但這6種灌木全株生物量在營養器官的分配上又可以分為三類較有差異的生境適應類型(圖4)。
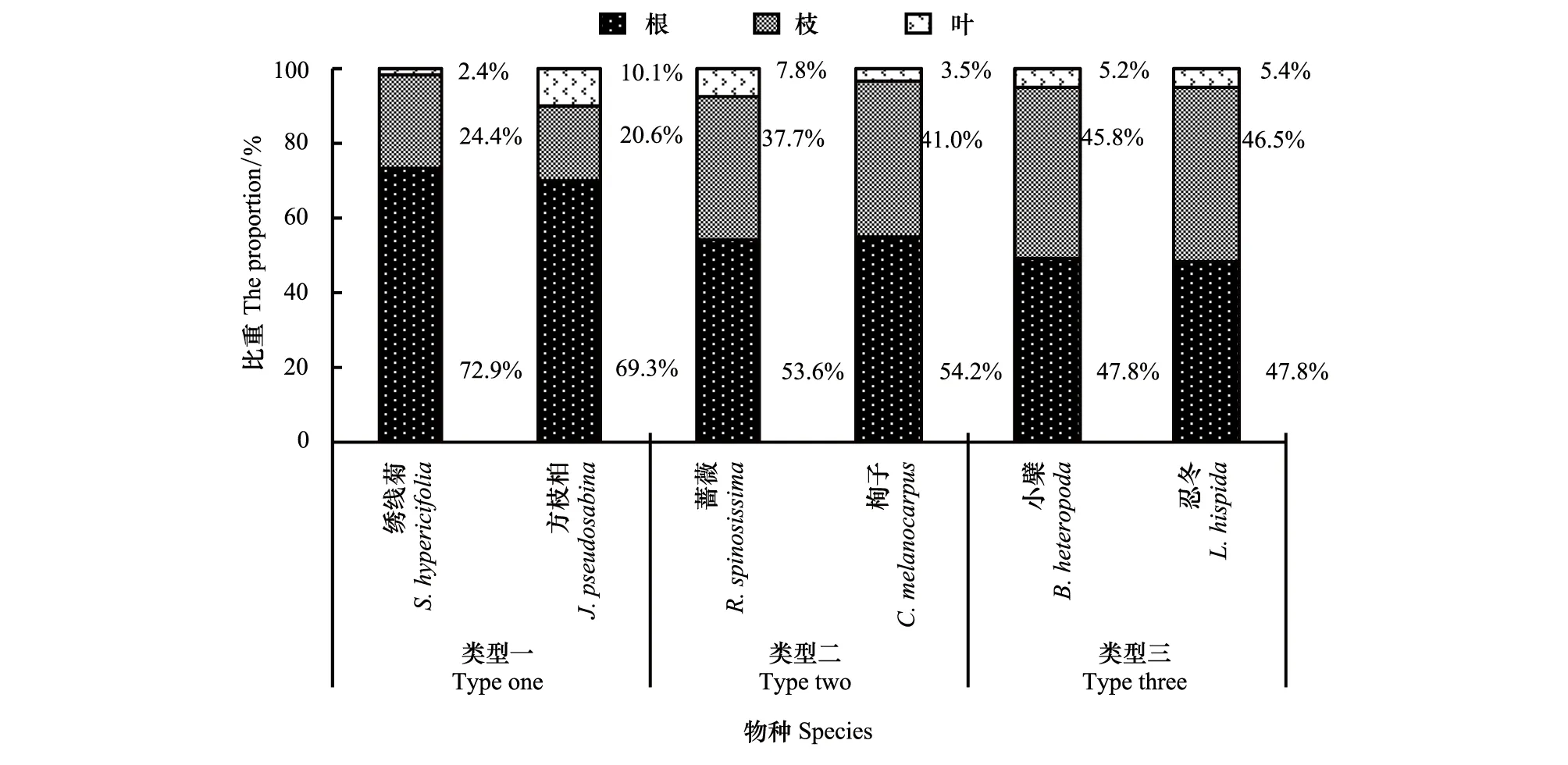
圖4 6種灌木全株生物量在營養器官分配上的適應性Fig.4 Adaptability of the allocation of six shrubs` total biomass in vegetative organs
類型一包括繡線菊、方枝柏。該類型的兩個物種主要分布在陽坡半陽坡、土層淺、礫石含量高的生境,它們的株形普遍矮小,全株生物量較低(圖1),根占全株生物量的大部分(70%左右),枝占全株生物量的超過20%(圖4)。此種類型表明,從土壤中獲取水分和養分是這兩個物種最嚴峻的生態壓力,對根系生物量的大投入體現了它們對較干旱和貧瘠生境的適應。
類型二包括薔薇、栒子。該類型的兩個物種主要分布在半陽坡、陰坡的林緣和林窗,它們株形較為高大,植株密度較高,全株生物量較高(圖1),根占全株生物量的略超過50%,枝占全株生物量的40%左右,葉生物量占全株生物量比例波動較大(圖4)。此種類型表明,種內和種間對光照的競爭在一定程度上是這兩個物種較主要的生存壓力,對營養器官生物量的分配體現了它們對水分養分條件較好、光照相對充足生境的適應特點。
類型三包括小檗、忍冬。該類型的兩個物種主要分布在林下、陰坡半陰坡的林緣或林窗,它們的株形相對較為纖細,全株生物量較高(圖1),根占全株生物量的略低于50%,地上部分的枝、葉生物量占全株生物量的50%以上,且葉生物量占全株生物量比例較為穩定(圖4)。此種類型表明,對光照的競爭是這兩個物種最主要的生態壓力,對營養器官生物量的分配體現了它們對光照條件稀缺生境的適應特點。
3.3 天山林區灌木根系生物量在徑級上分配策略的適應性分析
植物根系起著固定植株、攝取運輸和貯存水分與營養等重要作用[26]。不同徑級的根的功能是有一定區別的[38-39],其中,粗根通常在根系生物量中占主要組分,是主要的貯藏器官并起著固定植株的作用,但在水分和養分吸收、根土相互作用等方面的功能較小[26];而細根通常在根系生物量中占比較小[40],但細根是植物根系獲得水分和養分的最重要的途徑[19]。因此,植物對特定生境的適應不僅通過調節根系生物量占全株生物量比例來體現,也往往通過各徑級根系生物量占根系整體生物量比例來體現[41]。
從總體來看,天山林區6種代表性灌木均呈現出各徑級根系生物量隨徑級減小而占比減小的規律,這符合喬灌木物種通常的根系構型。但這6種灌木的各徑級根系生物量占比分配上又可以分為兩類較有差異的生境適應類型(圖5)。
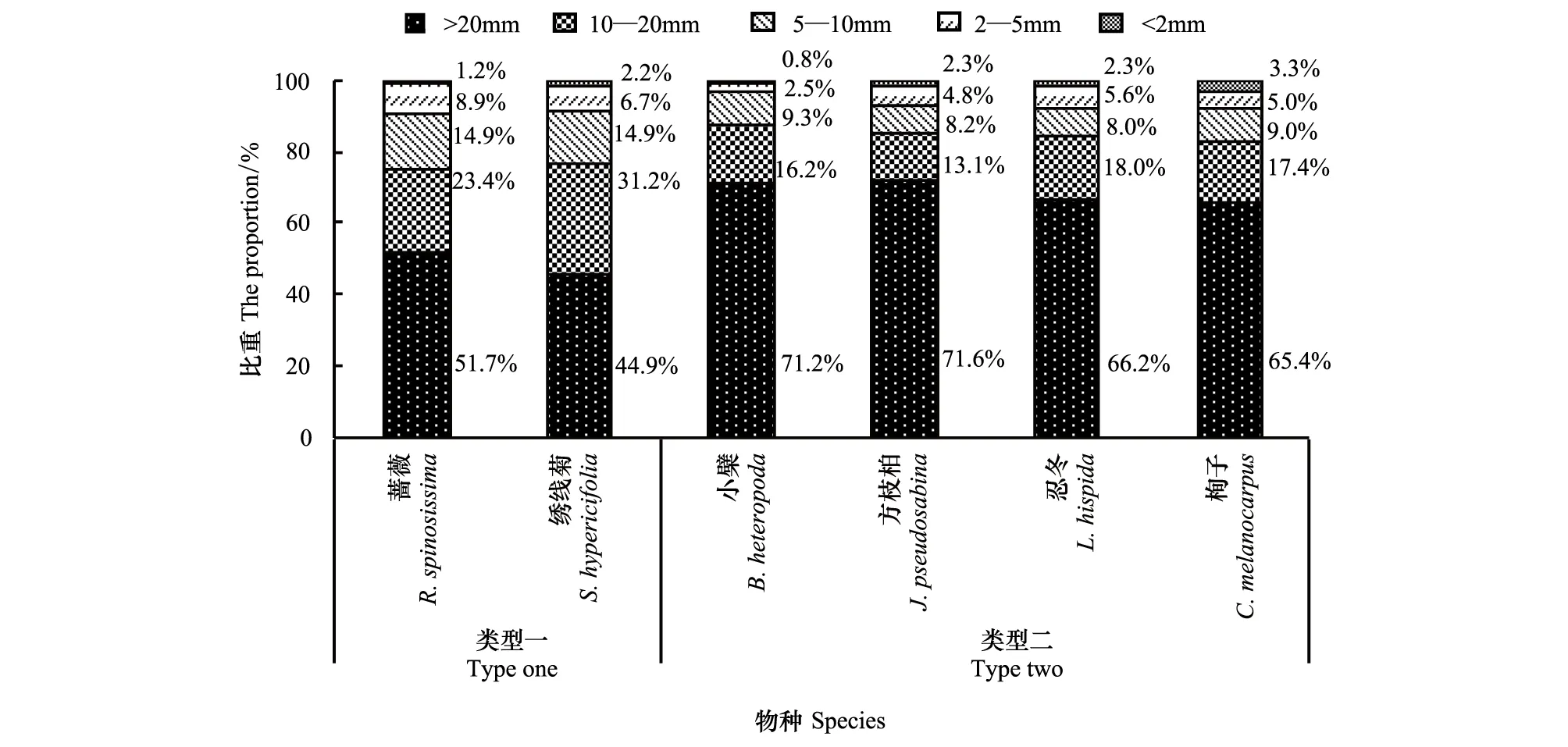
圖5 6種灌木根系生物量在徑級上分配的適應性Fig.5 Adaptability of the allocation of six shrubs` underground biomass in organs
類型一包括薔薇、繡線菊。該類型的兩個物種中,起固定植株作用的根樁(>20mm)占根系總生物量的50%左右,起延展根系空間作用的粗根(10—20mm)占有20%以上的比重,而在水分、養分的吸收方面發揮重要作用的中根(5—10mm)、小根(2—5 mm)、細根(<2 mm)占25%左右的較大比重(圖5)。此種類型表明,這兩個物種在各自的生境中要么存在較強的種內或種間的競爭壓力(例如薔薇),要么面對較為貧瘠干旱的土壤環境(例如繡線菊),因此需要大量更富有活性的徑級在10mm以下的根系來滿足植株旺盛生長對營養和水分的需求。因此,此類型屬于在競爭生境(薔薇)或脅迫生境(繡線菊)中適應的較強者。
類型二包括小檗、方枝柏、忍冬、栒子。該類型的4個物種中,起固定植株作用的根樁(>20mm)占根系生物量的70%左右,起延展根系空間作用的粗根(10—20mm)占有15%—20%的較穩定比重,而在水分、養分的吸收方面發揮重要作用的中根(5—10mm)、小根(2—5 mm)、細根(<2 mm)僅占15%左右的比重(圖5)。此種類型表明,這4個物種在各自的生境中要么處在光照條件稀缺的生境中(例如小檗、忍冬),要么面對較為貧瘠干旱的群落演替早期的土壤環境(例如方枝柏),要么面對較強的種內或種間競爭壓力的抑制(例如栒子),從而導致了它們的生長速率相對低下,其正常生長過程對營養和水分的需求相對較低,使得富有活性的徑級小于10mm的根系在根系總生物量中的占比較小。因此,此類型屬于適應脅迫生境(小檗、忍冬)、先鋒生境(方枝柏)或競爭生境(栒子)中適應的較弱者。
3.4 結論
(1)將D2H與V選為天山林區6種常見灌木全株生物量估測模型的自變量,得到各物種的全株生物量估測模型,各模型R2值均在0.850以上,且在P<0.05水平上達到顯著,表明模型模擬結果達到了較高的準確度。
(2)天山林區6種灌木的全株生物量在營養器官根、枝、葉上的分配均呈現出根>枝>葉的現象,且在根、枝上的分配比重差異不顯著,僅在葉上的分配比重有差異(P<0.05)。根系生物量在不同徑級上的分配均呈現為根樁(>20mm)>粗根(10—20mm)>中根(5—10mm)>小根(2—5 mm)>細根(<2 mm),6種灌木間僅在細根上的分配差異不顯著(P<0.05),在其他徑級根上的分配均有差異(P<0.05,其中根樁為P<0.01)。
(3)天山林區6種常見灌木全株生物量在營養器官上分配差異體現出各物種對生境差異的適應性,類型一為繡線菊、方枝柏,它們對較干旱和貧瘠生境較為適應;類型二為薔薇、栒子,它們對水分養分條件較好、光照相對充足生境較為適應;類型三為小檗、忍冬,它們對光照條件稀缺的生境較為適應。
(4)天山林區6種常見灌木根系生物量在徑級上的分配差異也體現出各物種對生境選擇的適應策略,類型一為薔薇、繡線菊,它們屬于在競爭生境(薔薇)或脅迫生境(繡線菊)中適應的較強者;類型二為小檗、方枝柏、忍冬、栒子,它們屬于適應脅迫生境(小檗、忍冬)、先鋒生境(方枝柏)或競爭生境(栒子)中適應的較弱者。
[1] 張峰, 上官鐵梁, 李素珍. 關于灌木生物量建模方法的改進. 生態學雜志, 1993, 12(6):67- 69.
[2] 趙振勇, 王讓會, 張慧芝, 王雷. 天山南麓山前平原檉柳灌叢地上生物量. 應用生態學報, 2006, 17(9):1557- 1562.
[3] 劉長成, 魏雅芬, 劉玉國, 郭柯. 貴州普定喀斯特次生林喬灌層地上生物量. 植物生態學報, 2009, 33(4):698- 705.
[4] 鄭紹偉, 唐敏, 鄒俊輝, 慕長龍. 灌木群落及生物量研究綜述. 成都大學學報:自然科學版, 2007, 26(3):189- 192.
[5] 曾慧卿, 劉琪璟, 馬澤清, 曾珍英. 基于冠幅及植株高度的檵木生物量回歸模型. 南京林業大學學報:自然科學版, 2006, 30(4):101- 104.
[6] 趙蓓, 郭泉水, 牛樹奎, 孫武, 張志旭. 大崗山林區幾種常見灌木生物量估算與分析. 東北林業大學學報, 2012, 40(9):28- 33.
[7] 袁素芬, 陳亞寧, 李衛紅, 劉加珍, 孟麗紅, 張麗華. 新疆塔里木河下游灌叢地上生物量及其空間分布. 生態學報, 2006, 26(6):1818- 1824.
[8] 盧振龍, 龔孝生. 灌木生物量測定的研究進展. 林業調查規劃, 2009, 34(4):37- 40, 45- 45.
[9] 劉恩斌, 周國模, 姜培坤, 葛宏立, 杜華強. 生物量統一模型構建及非線性偏最小二乘辯識—以毛竹為例. 生態學報, 2009, 29(10):5561- 5569.
[10] 劉恩斌, 李永夫, 周國模, 施擁軍, 莫路鋒. 生物量精確估算模型與參數辨識方法及應用. 生態學報, 2010, 30(10):2549- 2561.
[11] 黎燕瓊, 鄭紹偉, 宿以明, 龔良春, 慕長龍. 岷江上游干旱河谷引種番麻的生態適應性及生物量預測模型. 生態學報, 2009, 29(9):4820- 4826.
[12] 楊昊天, 李新榮, 劉立超, 賈榮亮, 王增如, 李小軍, 李剛. 荒漠草地4種灌木生物量分配特征. 中國沙漠, 2013, 33(5):1340- 1348.
[13] Weiner J. Allocation, plasticity and allometry in plants. Perspectives in Plant Ecology, Evolution and Systematics, 2004, 6(4):207- 215.
[14] Fabbro T, Korner C. Altitudinal differences in flower traits and reproductive allocation. Flora - Morphology, Distribution, Functional Ecology of Plants, 2004, 199(1):70- 81.
[15] 馮麗, 張景光, 張志山, 郭群, 李新榮. 騰格里沙漠人工固沙植被中油蒿的生長及生物量分配動態. 植物生態學報, 2009, 33(6):1132- 1139.
[16] 王政權, 郭大立. 根系生態學. 植物生態學報, 2008, 32(6):1213- 1216.
[17] 劉興良, 馬欽彥, 楊冬生, 史作民, 宿以明, 周世強, 劉世榮, 楊玉坡. 川西山地主要人工林種群根系生物量與生產力. 生態學報, 2006, 26(2):542- 551.
[18] 岳永杰, 余新曉, 慕長龍, 劉興良, 宿以明, 鄭紹偉. 岷江上游干旱河谷區人工林根系生物量及分布格局. 東北林業大學學報, 2008, 36(2):16- 18.
[19] 王向榮, 王政權, 韓有志, 谷加存, 郭大立, 梅莉. 水曲柳和落葉松不同根序之間細根直徑的變異研究.植物生態學報, 2005, 29(6):871- 877.
[20] 王力朋, 晏紫伊, 李吉躍, 王軍輝, 何茜, 蘇艷, 陳博, 馬建偉, 董菊蘭. 指數施肥對楸樹無性系生物量分配和根系形態的影響. 生態學報, 2012, 32(23):7452- 7462.
[21] 阿米娜木·艾力, 常順利, 張毓濤, 仇瑤, 何平. 天山云杉森林土壤有機碳沿海拔的分布規律及其影響因素. 生態學報, 2014, 34(7):1626- 1634.
[22] 曾慧卿, 劉琪璟, 馮宗煒, 馬澤清, 胡理樂. 紅壤丘陵區林下灌木生物量估算模型的建立及其應用. 應用生態學報, 2007, 18(10):2185- 2190.
[23] 陳遐林, 馬欽彥, 康峰峰, 曹文強, 張國華, 陳宗偉. 山西太岳山典型灌木林生物量及生產力研究. 林業科學研究, 2002, 15(3):304- 309.
[24] 楊昆, 管東生. 森林林下植被生物量收獲的樣方選擇和模型. 生態學報, 2007, 27(2):705- 714.
[25] 曾慧卿, 劉琪璟, 馬澤清, 張海清, 蔡哲, 曾珍英, 李軒然. 千煙洲灌木生物量模型研究. 浙江林業科技, 2006, 26(1):13- 17.
[26] 何列艷, 亢新剛, 范小莉, 高延, 馮啟祥. 長白山區林下主要灌木生物量估算與分析. 南京林業大學學報:自然科學版, 2011, 35(5):45- 50.
[27] Paton D, Nunez J, Bao D, Munoz A. Forage biomass of 22 shrub species from Monfrague Natural Park (SW Spain) assessed by log-log regression models. Journal of Arid Environments, 2002, 52(2):223- 231.
[28] 孫書存, 錢能斌. 刺旋花種群形態參數的通徑分析與亞灌木個體生物量建模. 應用生態學報, 1999, 10(2):155- 158.
[29] 林偉, 李俊生, 鄭博福, 郭建明, 胡理樂. 井岡山自然保護區12種常見灌木生物量的估測模型. 武漢植物學研究, 2010, 28(6):725- 729.
[30] 張倩媚, 溫達志, 葉萬輝, 孔國輝. 南亞熱帶常綠闊葉林林下層植物的生物量及其測定方法的探討. 生態科學, 2000, 19(4):62- 66.
[31] 王新云, 郭藝歌, 陳林, 劉任濤, 楊明秀, 謝騰騰. 荒漠草原不同林齡檸條灌叢生物量模型研究. 生物數學學報, 2013, 28(2):377- 383.
[32] 董道瑞, 李霞, 萬紅梅, 林海軍. 塔里木河下游檉柳灌叢地上生物量估測. 西北植物學報, 2012, 32(2):384- 390.
[33] 曾凡江, 郭海峰, 劉波, 曾杰, 邢文娟, 張曉蕾. 多枝檉柳和疏葉駱駝刺幼苗生物量分配及根系分布特征. 干旱區地理, 2010, 33(1):59- 64.
[34] Schenk H J, Jackson R B. Rooting depths, lateral root spreads and below-ground/above-ground allometries of plants in water-limited ecosystems. Journal of Ecology, 2002, 90(3):480- 494.
[35] Osone Y, Tateno M. Applicability and limitations of optimal biomass allocation models:a test of two species from fertile and infertile habitats. Annals of Botany, 2005, 95(7):1211- 1220.
[36] 黎燕瓊, 鄭紹偉, 龔固堂, 陳俊華, 唐波, 朱自芳, 吳雪仙, 慕長龍. 不同年齡柏木混交林下主要灌木黃荊生物量及分配格局. 生態學報, 2010, 30(11):2809- 2818.
[37] 劉興良, 郝曉東, 楊冬生, 劉世榮, 宿以明, 蔡小虎, 何飛, 馬欽彥. 臥龍巴郎山川滇高山櫟灌叢地上生物量及其模型. 生態學雜志, 2006, 25(5):487- 491.
[38] 劉瑞, 靳虎甲, 馬全林, 王耀琳, 李銀科, 孫濤, 宋德偉, 朱國慶. 甘肅景電灌區不同栽植年限枸杞生物量分配特征. 生態學雜志, 2012, 31(10):2493- 2500.
[39] Salas E, Ozier-Lafontaine H, Nygren P. A fractal root model applied for estimating the root biomass and architecture in two tropical legume tree species. Annals of Forest Science, 2004, 61(4):337- 345.
[40] 郭忠玲, 鄭金萍, 馬元丹, 韓士杰, 李慶康, 于貴瑞, 范春楠, 劉萬德, 邵殿坤. 長白山幾種主要森林群落木本植物細根生物量及其動態. 生態學報, 2006, 26(9):2855- 2862.
[41] 牛存洋, 阿拉木薩, 宗芹, 駱永明, 押田敏雄, 孫貴凡, 劉謙. 科爾沁沙地小葉錦雞兒地上-地下生物量分配格局. 生態學雜志, 2013, 32(8):1980- 1986.
Biomass estimation modeling and adaptability analysis of organ allocation in six common shrub species in Tianshan Mountains forests, China
QIU Yao1, CHANG Shunli1,*, ZHANG Yutao2, WANG Wendong2, HE Ping1, WANG Huijie1, XIE Jin1
1KeyLaboratoryofOasisEcology,CollegeofResourceandEnvironmentScience,XinjiangUniversity,Urumqi830046,China2InstituteofForestEcology,XinjiangAcademyofForestry,Urumqi830063,China
Estimation modeling of the total biomass of shrubs is associated with certain difficulties. Moreover, merely studying the effect of organ allocation adaptability on shrub biomass is not sufficient. Thus, to investigate the effectiveness of biomass estimation modeling on organ allocation in plants, we focused on six common shrub species found in the forests of Tianshan Mountains, northwest China. To conduct a community investigation, we selected three sample areas in the eastern, central, and western regions of Tianshan Mountains. A number of whole standard plants from six shrub species were harvested to determine total biomass, which was divided into the, biomass of the roots, branches, and leaves, along with different classes of root diameter.D2H(the square of the diameter multiplied by height) andV(the area of the crown multiplied by height) were selected as the independent variables in the estimation models. Optimal estimation models for the total biomass of the six shrub species were constructed using regression analysis. These models were used to compare differences in total biomass allocation to vegetative organs and in biomass allocation to roots of each root diameter class in the shrub species. The following results were obtained. First, the total biomass ofBerberisheteropoda,Lonicerahispida, andCotoneastermelanocarpuswas approximately 8.48 to 9.01 kg, whereas that ofRosaspinosissima,Spiraeahypericifolia, andJuniperuspseudosabinawas approximately 2.71 to 3.20kg. Second, the optimal estimation models for the total biomass ofR.spinosissima,S.hypericifolia, andC.melanocarpuswere functions based onVas an independent variable, whereas those forB.heteropoda,L.hispida, andJ.pseudosabinawere functions based onD2Has an independent variable. The value ofR2for each estimation model was greater than 0.850, and all the models reached a high degree of accuracy at the 0.05 significance level. Third, differences in the proportional allocation of total biomass to roots and branches of the six shrub species were not significant, whereas that to leaves was significant at the 0.05 significance level. The root biomass of all six shrub species decreased with decreasing root diameter class, with significant differences in the proportional allocation of biomass to roots in roots with a diameter greater than 2 mm (significant at the 0.05 and 0.01 significance levels for roots with diameters greater than 20mm). For all examined shrub species, we conclude that the differences in total biomass allocation to vegetative organs and in biomass allocation to the roots of each diameter class reflect adaptive strategies to their respective habitats.
biomass modeling; root classification; adaptive strategy; shrub; Tianshan Mountains
新疆維吾爾自治區財政林業科技專項資金項目(xjlk(2013)001); 國家自然科學基金項目(31060075); 新疆自治區公益性科研院所基本科研業務費專項資金項目(《天山東部與西部灌木林生物量及生產力研究》201414)
2014- 04- 28; < class="emphasis_bold">網絡出版日期:
日期:2015- 05- 19
10.5846/stxb201404280841
*通訊作者Corresponding author.E-mail:ecocsl@163.com
仇瑤, 常順利,張毓濤,王文棟,何平,王慧杰, 謝錦.天山林區六種灌木生物量的建模及其器官分配的適應性.生態學報,2015,35(23):7842- 7851.
Qiu Y, Chang S L, Zhang Y T, Wang W D, He P, Wang H J, Xie J.Biomass estimation modeling and adaptability analysis of organ allocation in six common shrub species in Tianshan Mountains forests, China.Acta Ecologica Sinica,2015,35(23):7842- 7851.
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
艦船科學技術(2022年13期)2022-08-11 09:30:02
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
鐵道通信信號(2020年9期)2020-02-06 09:15:22
數學大王·趣味邏輯(2019年5期)2019-06-13 20:27:43
小學科學(學生版)(2019年5期)2019-05-21 01:00:18
經濟技術協作信息(2018年30期)2018-11-22 06:20:24
光學精密工程(2016年6期)2016-11-07 09:07:19
核科學與工程(2015年4期)2015-09-26 11:59:03
