三峽水庫奉節以東秭歸和巫山段消落帶植物群落動態特征
2015-02-11 02:06:28朱妮妮郭泉水秦愛麗裴順祥馬凡強簡尊吉
生態學報 2015年23期
朱妮妮,郭泉水,* ,秦愛麗,裴順祥,馬凡強,朱 莉,3,簡尊吉
1 中國林業科學研究院森林生態環境與保護研究所,國家林業局森林生態環境重點實驗室, 北京 100091;2 中國林業科學研究院華北林業實驗中心,北京 102300 3 河南科技大學農學院,洛陽 471003
三峽水庫奉節以東秭歸和巫山段消落帶植物群落動態特征
朱妮妮1,郭泉水1,*,秦愛麗1,裴順祥2,馬凡強1,朱 莉1,3,簡尊吉1
1 中國林業科學研究院森林生態環境與保護研究所,國家林業局森林生態環境重點實驗室, 北京 100091;2 中國林業科學研究院華北林業實驗中心,北京 102300 3 河南科技大學農學院,洛陽 471003
基于2008—2012 年對三峽水庫奉節以東秭歸和巫山段消落帶固定樣地不同海拔區段植物群落的5a定位監測,研究消落帶植物群落的物種組成、優勢植物、植物生活型和物種多樣性的動態變化,結果表明:1)截止2012年,消落帶海拔156—172 m區段共經歷了4 次水庫水位漲落。經歷首次后(2009 年),消落帶原生植物由55 科147 種減少到18 科33 種,經歷4 次后(2012 年),減少到14 科39 種。與經歷水庫水位漲落前(2008 年)比較,經歷首次后的科數減少了67.3%,種數減少了77.6%;經歷4 次后的科數減少了74.5%,種數減少了73.5%。在消落帶原生植物減少的同時,出現了許多“新”植物。經歷首次后出現了49 種,經歷4 次后出現了23 種,分別占調查當年樣地植物種類總數的59.8%和32.9%。海拔172—175 m區段共經歷了2 次水庫水位漲落,消落帶原生植物由40科91 種(2008 年)減少到了13 科20種。與經歷水庫水位漲落前比較,科數減少了67.5%,種數減少了78.0%。出現“新”植物21 種,約占調查當年樣地植物種類總數的44.7%。通過對歷次調查中消落帶植物“消失”和“出現”的數量比較表明,消落帶植物對經歷首次水庫水位漲落的反應最為敏感,此后,雖又經歷過幾次水庫水位漲落,但其變化速率趨于減小。2)不同海拔區段、不同生態適應型植物的“消長”動態和優勢種組成不完全相同。海拔156—172 m區段,經歷4 次水庫水位漲落后,在消落帶植物群落中占優勢的草本植物種為菊科(Compositae)的鬼針草(Bidenspilosa)、禾本科(Gramineae)的狗牙根(Cynodondactylon)、毛馬唐(Digitariachrysoblephara)、狗尾草(Setariaviridis)、莎草科(Cyperaceae)的碎米莎草(Cyperusiria),占優勢的灌木樹種為漆樹科(Anacardiaceae)的鹽膚木(Rhuschinensis)和大戟科(Euphorbiaceae)的算盤子(Glochidionpuberum);在海拔172—175 m區段,除鬼針草、毛馬唐仍為優勢種外,還增加了大戟科的湖北算盤子(Glochidionwilsonii),馬鞭草科(Verbenaceae)的黃荊(Vitexnegundo),葡萄科(Vitaceae)的五葉地錦(Parthenocissusquniquefolia)等樹種。3)消落帶植物群落的優勢生活型為一年生和多年生草本;物種多樣性隨著水庫水位漲落次數的增加總體變化呈減少趨勢。4)三峽水庫水位周期性漲落導致消落帶發生水陸環境交替變化,不同生態適應型植物對變化生境的適應能力有所不同,是消落帶植物群落發生變化的主要驅動因素。
群落動態;植物生活型;物種多樣性;三峽水庫;消落帶
三峽庫區消落帶相對高差30m,總面積348.93 km2,是目前我國相對高差和面積最大的水庫消落帶。受水庫水位周期性漲落的影響,現已成為庫區生態系統中物質轉化、能量流動、信息傳輸與轉換等最為活躍且最不穩定的生態脆弱帶[1]。消落帶植物群落是消落帶生態系統的重要組成部分,在維護消落帶生態系統穩定和生態功能發揮等方面起著重要作用,也是水庫管理的重要對象[2-3]。及時準確掌握消落帶植物群落動態,對于深入了解三峽水庫消落帶生態環境的變化以及消落帶植被恢復和重建都具有重要的理論和實踐意義。
植物群落的物種組成和結構變化是植物群落演變的具體表現特征[4]。近年來,許多學者圍繞三峽庫區消落帶植物群落結構(如種類組成、物種豐富度或多樣性、優勢植物種群結構等)的變化和群落演替開展了廣泛的調查和研究。熊平生等[5]、王勇等[6]和白寶偉[7]等基于植物群落演替理論和三峽自然消落帶的植被分析,對消落帶植物群落動態進行了推測;盧志軍等[8]通過固定樣方調查,對被水淹過的海拔156 m以下區段和當年未被水淹區段的植物種類組成、物種多樣性和草本層生物量進行過比較;胡波等[1-2, 9-10]采用“空間推時間”的方法,對消落帶植物群落的演替趨勢進行了預測;郭泉水等[3]通過固定樣地調查,研究了三峽庫區消落帶陸生植被首次經歷水陸生境變化的響應。研究結論一致表明:受三峽水庫水位漲落的影響,消落帶植物的生存條件發生了巨大改變;消落帶植物群落的正常發育和演替過程已受到嚴重干擾;不同植物因適應水陸交替變化生境的能力不同,其“消長”動態表現各異。這些研究提供了許多關于消落帶植物群落動態變化的信息,對于深入揭示消落帶動態變化規律具有一定的理論參考價值,不足之處在于:(1)以往研究大多集中在水庫蓄水位達到海拔175 m之前,而對達到海拔175 m以后植物群落的調查研究較少。三峽水庫正常運行的水位在海拔145—175 m,因此,其研究結果大多反映的是消落帶局部地段植物群落的變化。(2)大多數研究采用的是“一次性調查”或“空間推時間”的方法,對于經歷多次水庫水位漲落持續干擾下消落帶植物群落動態變化的定位研究還很少涉足,特別是缺乏對三峽庫區奉節以東地區消落帶植物群落動態的持續監測和研究。經歷水庫水位漲落后,在消落帶上出現的大多是森林群落次生演替早期的先鋒植物[3],這些植物種的“消長”波動性很大。“一次性調查”或以此調查為基礎采用“空間推時間”的方法,對消落帶植物群落動態變化趨勢預測的結果必然會存在較大的不確定性。三峽庫區的地勢東部高,西南部低。奉節位于庫區東部和西南部的分界處。奉節以東庫岸以中山山地為主[11],為峽谷地貌。消落帶的坡度大,水土流失嚴重,為三峽庫區滑坡、崩塌等地質災害多發區。地形是區域生態環境的一個重要因子。地形不同,氣候、土壤、植被等資源及其分布以及人類對資源的利用方式和程度都會有所不同[11]。生物多樣性是環境的產物。受不同生態環境因子及其生態過程的綜合影響,消落帶植物群落的組成、結構及其變化過程都會有所不同[12-14]。而目前尚未見針對三峽水庫奉節以東峽谷地貌區消落帶植物群落動態的研究報道。
本文以2008 年在三峽水庫奉節以東巫山和秭歸段典型消落帶上設置的4 塊固定監測樣地為依托,在獲得經歷水庫水位漲落前消落帶植物群落物種組成和結構等初始資料的基礎上,區分海拔156—172 m和172—175 m區段,對消落帶植物群落的種類組成及不同植物的“消長”動態、植物生活型和物種多樣性的變化進行了為期5a的定位監測,旨在通過對多年定位觀測結果的比較和分析,揭示三峽水庫奉節以東地區消落帶植物群落在水庫水位漲落持續干擾下的動態變化規律和發展趨勢,為全面評估三峽水庫運行對消落帶植物群落的影響以及消落帶植被恢復和重建中的物種選擇提供科學依據。
1 材料與方法
1.1 研究區概況
研究區位于三峽水庫奉節以東干流的秭歸段和巫山段。其典型的地理特征是:水庫干流兩側山體的海拔多在1200m左右。山勢陡峭,長江“三峽”中著名的西陵峽和巫峽兩個最大的峽即位于此。消落帶的坡度一般在25°以上;2/3區段的基巖為石灰巖,只有秭歸段部分地段分布著花崗巖。研究區的地形和地質條件在三峽水庫奉節以東峽谷地貌區具有一定的代表性。
固定樣地設在秭歸縣茅坪鎮和巫山縣巫峽鎮消落帶內。兩地氣候類型一致,均屬中亞熱帶濕潤季風氣候。其中,秭歸縣茅坪鎮的年均溫18 ℃,年均降雨量1 049.3 mm,≥10℃年活動積溫 5 723.6 ℃,全年無霜期305 d;巫山縣巫峽鎮年均溫18.4 ℃,年均降雨量1 100mm,≥10℃ 年活動積溫 5 857.3 ℃,全年無霜期306 d。地帶性植被類型也一致,均為亞熱帶常綠闊葉林。但由于人類的長期干擾,在水庫蓄水前消落帶的地帶性植被已被全部破壞,取而代之的是人類干預下的各種次生植被和人工植被。為了較全面反映研究區的植被特征,固定樣地從在研究區有一定代表性的次生植被和人工植被中選擇。各樣地經歷水庫水位漲落前的自然概況見表1。
消落帶水陸交替變化與水庫水位漲落同步。不同年份水庫水位漲落變化有所不同。2008 年冬季水庫最高蓄水位首次達到海拔172 m,此前,最高蓄水位為海拔156 m。2009 年與2008 年相同。2010年冬季水庫最高蓄水位上升至海拔175 m,此后,其最高水位一直維持在該海拔高度上運行。截止2012年8月,海拔156—172 m區段共經歷了4 次水庫水位漲落,海拔172—175 m區段共經歷了2次。研究時段內不同年份的水庫水位變化見圖1[15]。
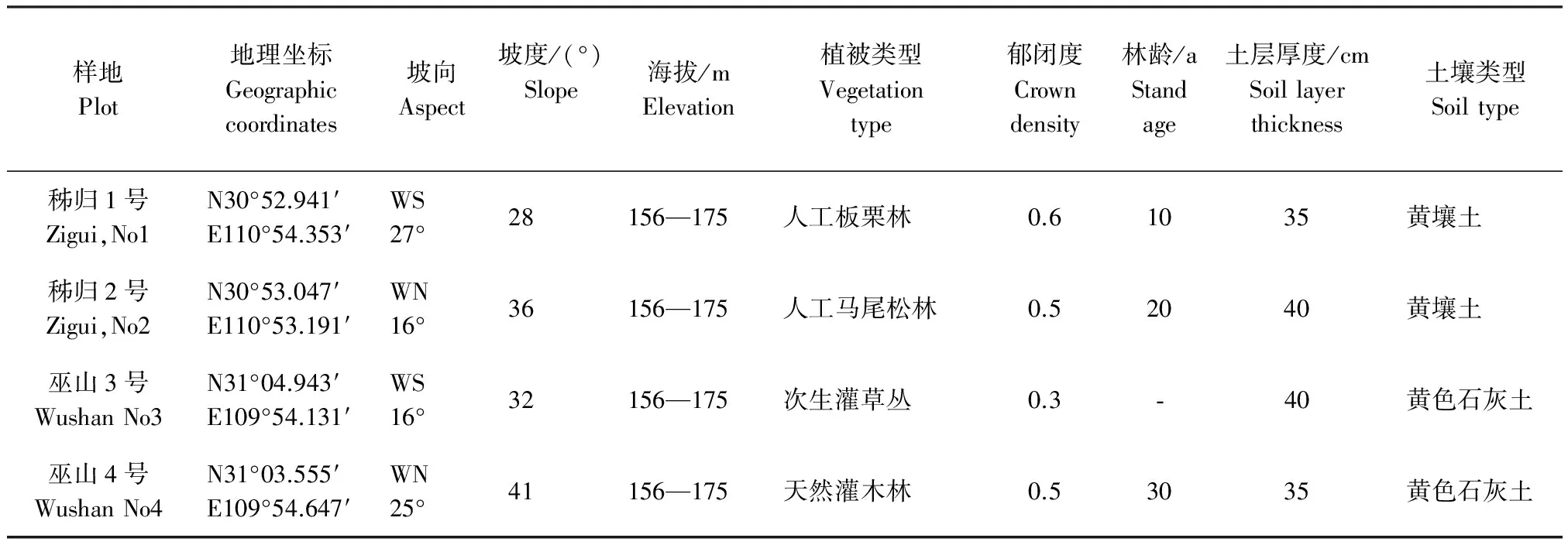
表1 樣地自然概況Table 1 Natural conditions of the sample plot
2008年秭歸1號和2號樣地的板栗和馬尾松被砍伐
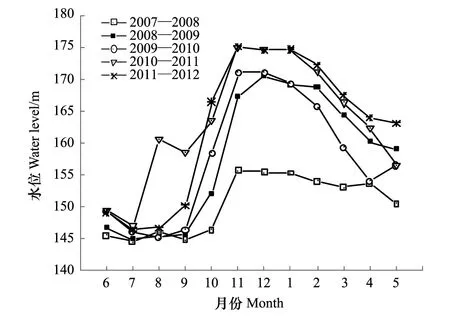
圖1 三峽水庫水位變化(2007—2012年) Fig.1 Water-level fluctuation of the Three Gorges Reservoir(2007—2012)
1.2 樣地設置和調查
固定樣地設置于2008 年8 月。在巫山和秭歸消落帶內各設置2 塊。巫山的2 塊樣地設在水庫干流左右兩岸,秭歸的2 塊樣地均設在水庫干流的右側。各樣地的寬度和長度的投影距離均為15 m和19 m。寬邊設在海拔156 m(2008 年水庫最高蓄水位),上線設在海拔175 m。樣方設置采用相鄰格子法。樣方大小因調查對象而異。喬木幼苗幼樹和灌木樣方為2 m × 2 m,草本為1 m × 1 m。調查內容包括:植物種類,不同植物的植株數量、平均高度、蓋度和植物生活型。其中,叢生草本和灌木植株數量調查的方法是:以地面以上是否為單株存在為判別標準,至于其地面以下植株與植株之間或不同植株的根系之間是否連接不予考慮。
固定樣地設置后,沿樣地4邊,每隔投影距離5 m,埋1 根水泥樁,并在頂端涂上紅漆,以做長期觀測的永久標記。調查時間分別為2008年8月、2009年8月、2012年8月。2008年8月首次調查,獲取海拔156—175 m區段經歷水庫水位漲落前消落帶植物群落的初始資料,2009年8月和2012年8月對經歷首次和4 次水庫水位漲落后植物群落的動態進行了兩次監測調查。
1.3 數據處理與分析
由于消落帶海拔156—172 m區段和海拔172—175 m區段經歷的水庫水位漲落次數不同,為此,在資料整理時,將兩個海拔區段作為兩個獨立的統計單元進行處理。
統計指標包括:不同植物在不同調查年份的生命表征、植物生活型組成、重要值、群落的物種豐富度、Shannon-Weiner指數、Simpson指數、Pielou均勻度指數。其中,消落帶植物生活型劃分參考《中國植被》[16]的生活型分類系統,共區分出喬木、灌木、藤本、多年生草本、一年生草本等5種生活型。植物重要值、物種豐富度及多樣性指數參考有關文獻[17-18]提供的計算公式計算。
數據整理、計算和制圖均采用Excel 7.0和Origin 9軟件完成。
2 結果與分析
2.1 消落帶植物群落種類組成及其動態變化
2.1.1 消落帶植物群落不同植物的“消長”變化
區分海拔156—172 m和172—175 m兩個區段,按不同調查年份,對4 塊固定樣地中的不同植物在樣地中出現和“消失”的調查記錄分別統計,結果見表2。
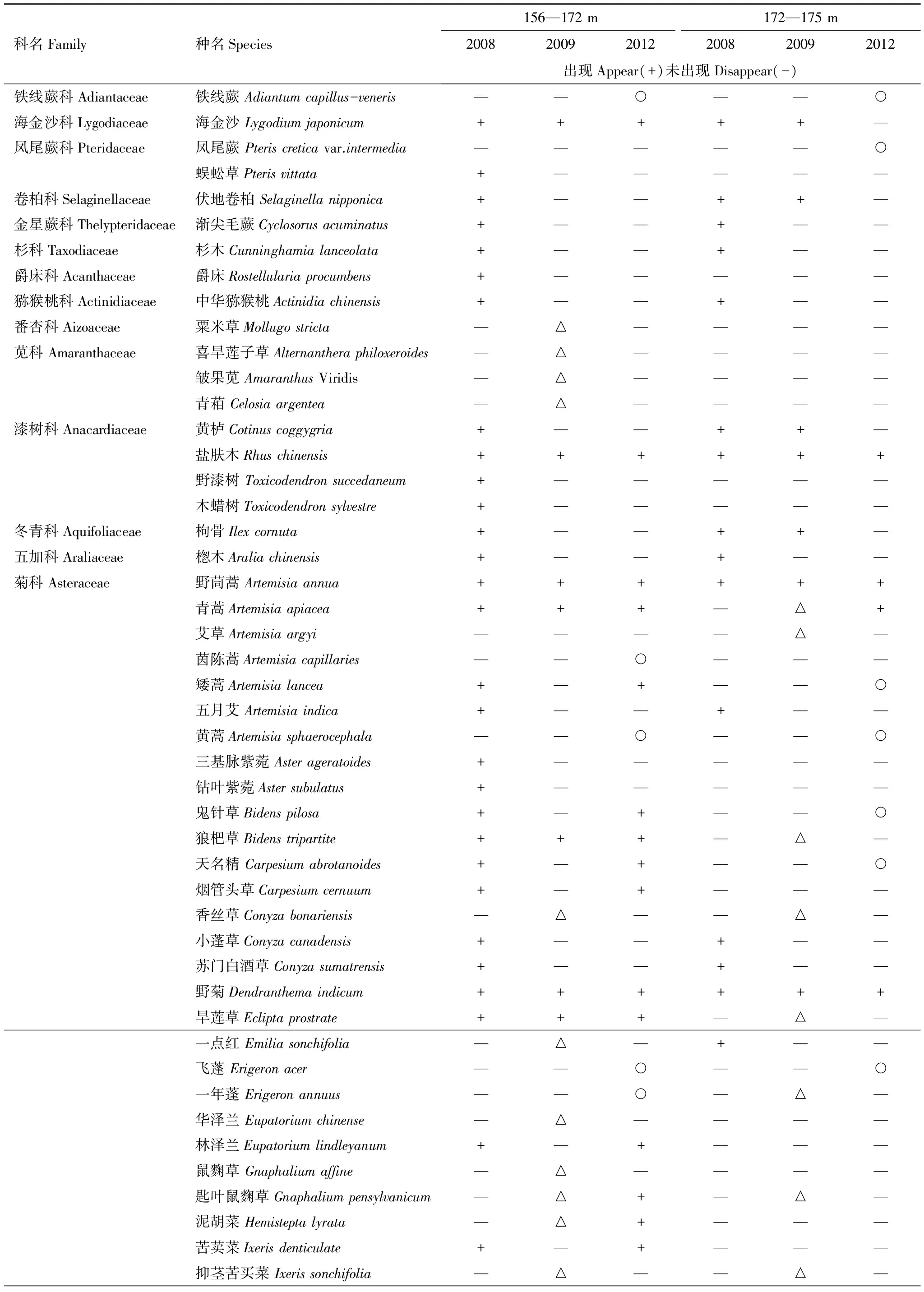
表2 2008—2012年三峽水庫消落帶植物的消長動態Table 2 Plant community "growth" dynamics in Hydro-fluctuation Belt of the Three Gorges Reservoir(2008—2012)
* 表中△表示在2008年未出現,2009年新出現的植物; ○表示2008年和2009年均未出現,僅在2012年出現的植物
從表2可概括出2008—2012年消落帶植物群落種類組成變化的基本特征如下:
(1)原來生長在消落帶的大多數植物經歷水庫水位漲落后消失,同時也有許多“新”植物出現。不同植物的消長過程不同,有些種經歷首次水位漲落后即“永久消失”,如棕櫚(Trachycarpusfortunei)、毛葡萄(Vitisheyneana)以及多種喬灌木;有些種在經歷首次水庫水位漲落后 “消失”,但經歷多次水位漲落后又有出現,如矮蒿(Artemisialancea),苧麻(Boehmerianivea)等; 也有一些種在樣地上始終存在,如野茼蒿(Artemisiaannua)、野菊(Dendranthemaindicum)等。不同植物的消長動態,反映了不同生態適應型植物對消落帶水陸變化生境響應程度上的差異。
(2)不同調查年份、不同海拔區段消落帶植物群落的數量特征隨著經歷水庫水位漲落次數、淹水深度和持續水淹時間長短而變化。經歷水庫水位漲落前(2008年8月),在海拔156—172 m區段共調查到原生植物55 科147 種。經歷首次水庫水位漲落后(2009年8月),減少到18 科33 種。經歷4 次水庫水位漲落后(2012年8月),減少到14 科39 種。與2008年比較,2009年和2012年科數分別減少了67.3%和74.5%,種數分別減少了77.6%和73.5%。對不同調查年份科、種的變化速率的比較發現,消落帶植物對首次經歷水庫水位漲落的反應最為敏感,而保存下來的植物雖又經歷過幾次水庫水位漲落,但其變化速率趨于減小。如2009—2012年這3年間,原生植物的科數僅減少了22.2%,與經歷首次水庫水位漲落后的變化(減少了67.3%)明顯不同。
在海拔172—175 m區段,2008年8月調查到原生植物40科91 種,2009年8月減少到24科37 種,2012年8月減少到13科20種。與2008年比較,2009年和2012年的科數分別減少了40.0%和67.5%,種數減少了59.3%和78.0%。該區段從2010年冬季開始受水庫水位漲落影響,截止2012年8月,僅經歷了2 次水庫水位漲落的影響。2008年8月到2009年8月植物科、種的變化,屬自然環境下植物種群的消長過程。將此期間的變化率(40.0%)與經歷2次水庫水位漲落影響下的變化率(67.5%)比較,凸顯了水庫水位漲落對消落帶植物種群的強烈干擾作用。將該區段科數和種數減少的幅度與海拔156—172 m區段比較可見,海拔172—175 m區段的科數減少的幅度小,而種數減少的幅度相差無幾。反映出經歷水庫水位漲落次數、淹水深度、水淹持續時間等對消落帶植物群落不同的影響效應。
(3)在海拔156—172 m區段,經歷首次水庫水位漲落后(2009年8月),共記錄到49 種“新”植物,約占調查當年樣地植物種類總數的59.8%;經歷4 次水庫水位漲落后(2012年8月),為 23 種,約占調查當年樣地上植物種類總數的32.9%。該結果顯示,消落帶上“新”植物也是以經歷首次水位漲落后出現最多,而隨著經歷水庫水位漲落次數的增加,“新”出現的植物也在逐漸減少。在海拔172—175 m區段,出現了21 種“新”植物,約占調查當年樣地上植物種總數的44.7%。明顯小于海拔156—172 m區段經歷首次水庫水位漲落后“新”植物在群落中所占的比例(59.8%)。
2.1.2 消落帶的優勢植物及其變化
對2008—2012年不同調查年份、不同植物的重要重要值分別計算,并提取在此期間某個調查年份植物重要值在10以上的植物種類列如表3。
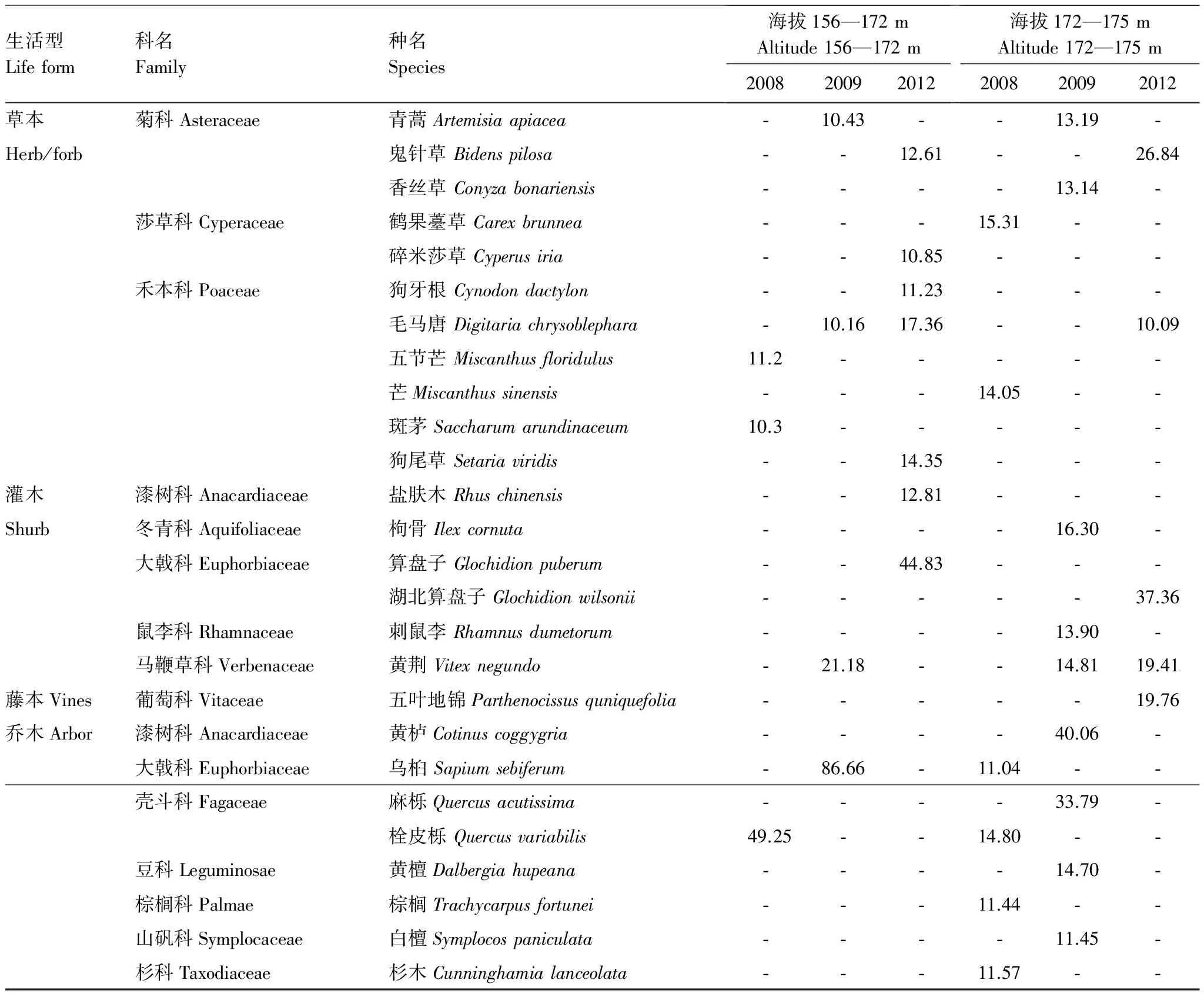
表3 2008—2012年三峽水庫消落帶優勢植物的重要值Table 3 Important values of dominant plants in Hydro-fluctuation Belt of the Three Gorges Reservoir(Year 2008—2012)
從表3可以看出,在海拔156—172 m區段,經歷4 次水庫水位漲落后(2012年),草本植物的優勢種為菊科(Asteraceae)的鬼針草(Bidenspilosa)、莎草科(Cyperaceae)的碎米莎草(Cyperusiria),禾本科(Poaceae)的狗牙根(Cynodondactylon)、毛馬唐(Digitariachrysoblephara)、狗尾草(Setariaviridis)。灌木優勢種為漆樹科(Anacardiaceae)的鹽膚木(Rhuschinensis)、大戟科(Euphorbiaceae)的算盤子(Glochidionpuberum),未見喬木優勢種。在海拔172—175 m區段,除鬼針草和毛馬唐仍為優勢種外,還增加了大戟科的湖北算盤子(Glochidionwilsonii),馬鞭草科(Verbenaceae)的黃荊(Vitexnegundo),葡萄科(Vitaceae)的五葉地錦 (Parthenocissusquniquefolia)。與海拔156—172 m區段比較,海拔172—175 m區段灌木和藤本的優勢種有所增加。
2.2 消落帶植物群落的生活型組成及其變化
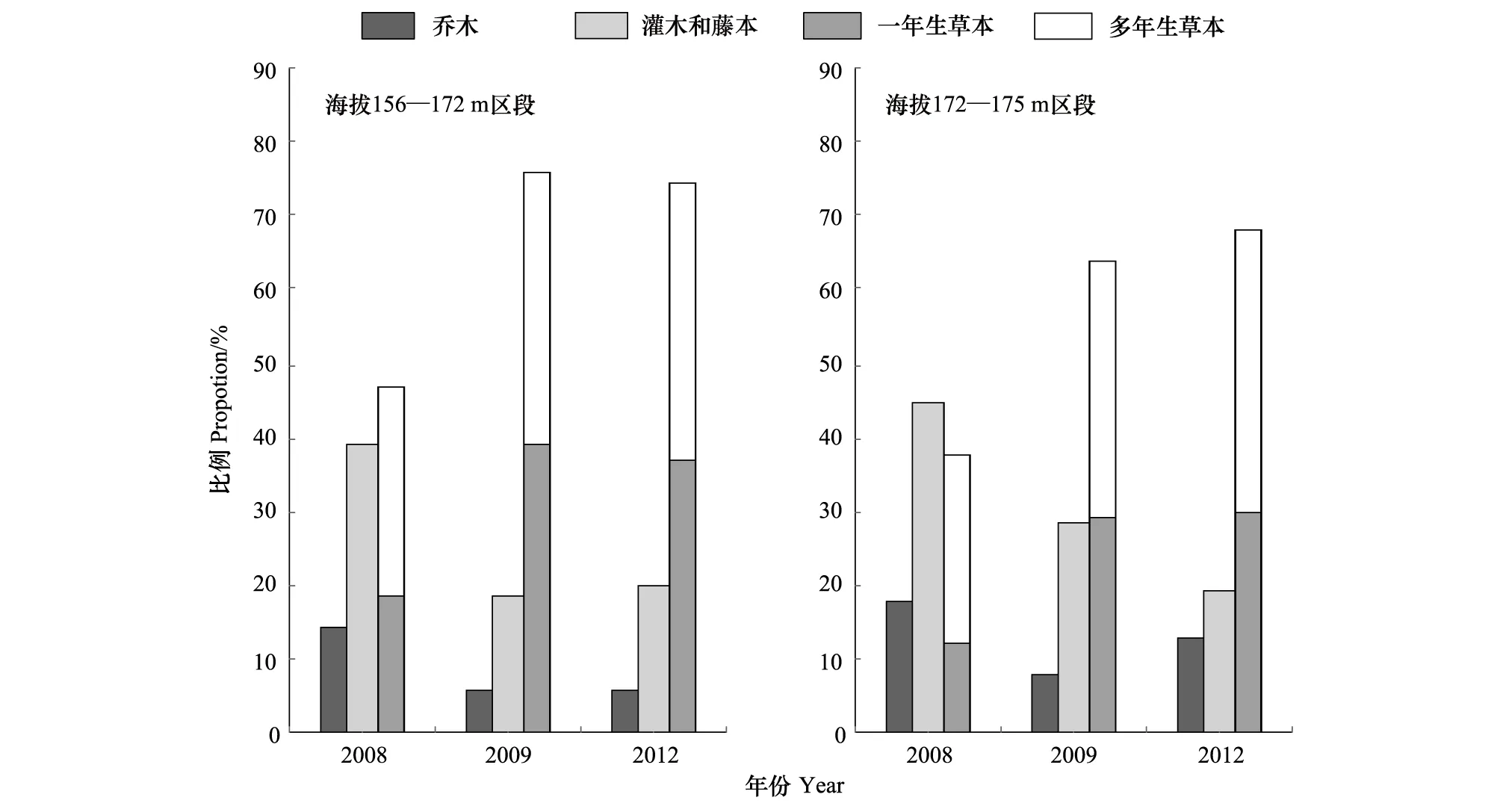
圖2 2008—2012年三峽水庫消落帶植物生活型組成的變化Fig.2 Changes in the composition of plant life form in Hydro-fluctuation Belt of the Three Gorges Reservoir(2008—2012)
2008—2012年,消落帶不同海拔區段植物生活型組成的總體變化趨勢是:草本植物在群落中所占比例增加,木本植物所占比例減少(圖2)。海拔156—172 m區段,經歷首次水庫水位漲落后,灌木和藤本在群落中所占比例由2008年的38.8%,減少到18.3%,喬木由14.3%減少到6.1%,而一年生草本則由18.4%增加到39.0%,多年生草本由28.6%增加到36.6%。經歷4 次水庫水位漲落影響后(2012年),灌木和藤本在群落中所占比例減少到20.0%,喬木減少到5.7%,一年生草本增加到37.1%,多年生草本增加到37.1%。海拔172—175 m區段經歷2次水庫水位漲落后,灌木和藤本在群落中所占比例由2008年的45.1%,減少到19.1%,喬木由17.6%,減少到12.8%,一年生草本由12.1%,增加到29.8%,多年生草本由25.3%增加到38.3%。
2.3 消落帶植物群落的物種多樣性及其變化
對群落的物種豐富度R、Shannon-Weiner指數H′、Simpson指數Ds、Pielou均勻度指數J的計算結果見圖3。
由圖3可以看出,在2008—2012年,海拔156—172 m區段草本的R、H′、Ds指數均呈先升后降的變化趨勢,均勻度J指數為單調下降;灌木和藤本的H′、Ds、R和J指數均為單調下降。海拔172—175 m區段,草本的H′、Ds指數為單調下降,R指數為先升后降,J指數為先降后升。灌木和藤本的H′、Ds、R指數為單調下降,J指數為單調上升;喬木各項測定指標的變化趨勢與灌木和藤本相同。
從總體變化趨勢分析,灌木和藤本與喬木的物種多樣性指數下降最為明顯。海拔156—172 m區段和海拔172—175 m區段的均勻度指數變化趨勢相反;草本植物的物種多樣性H′、Ds、J指數變化總體上呈下降趨勢;經歷首次水庫水位漲落后,物種豐富度增加,而經歷多次水庫水位漲落后又有所下降。
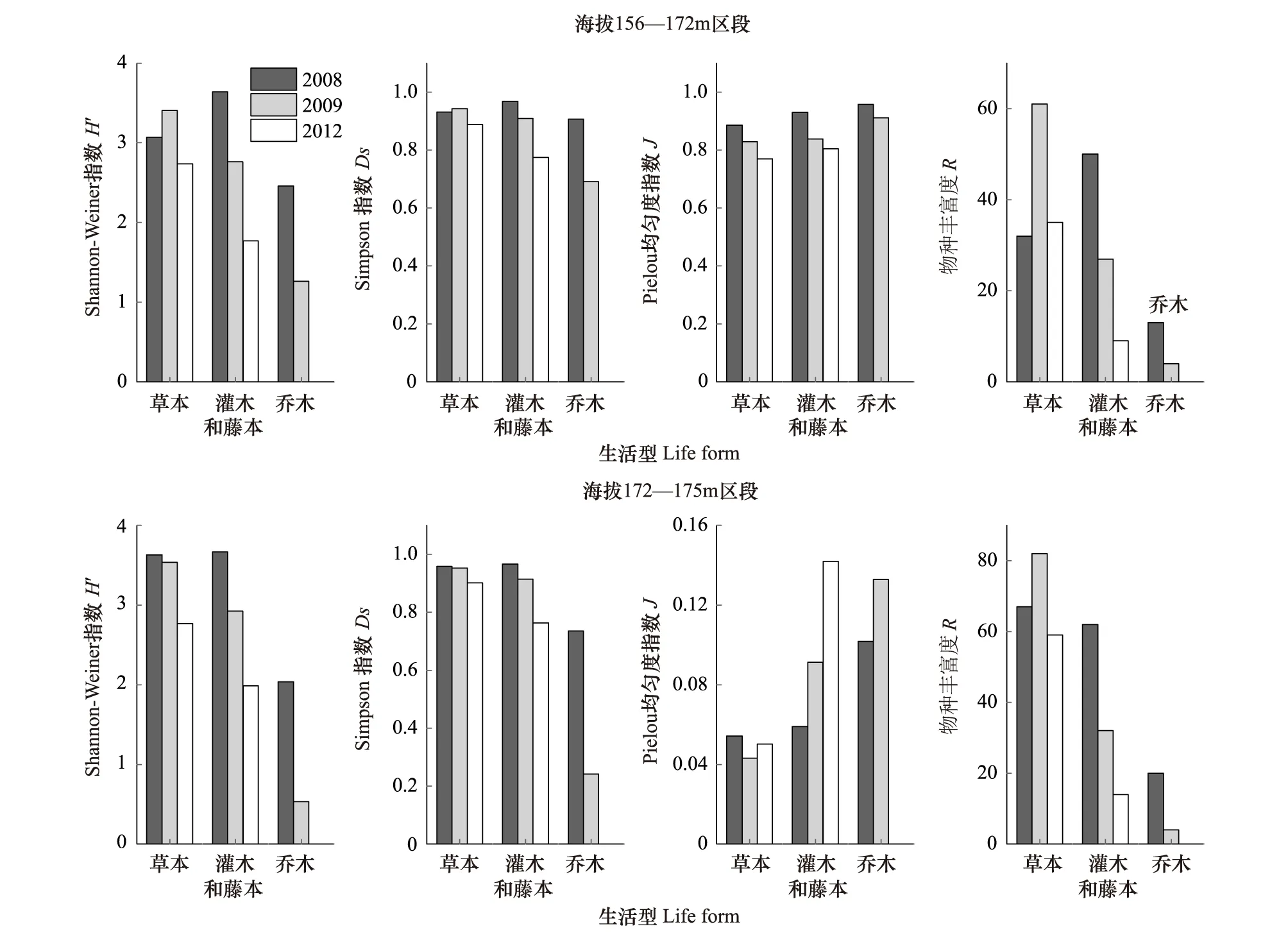
圖3 2008—2012年三峽水庫消落帶植被的物種多樣性變化Fig.3 Changes in species diversity of plant community in Hydro-fluctuation Belt of the Three Gorges Reservoir(2008—2012)
3 結論與討論
3.1 植物的生態適應性與消落帶水陸生境變化
消落帶原生植物在經歷多次水位漲落后大幅度減少。其中,三峽水庫水位變動的時機、持續時間、幅度、頻率、變化率改變了消落帶的物理環境和資源養分的分布[9],以及不同生態適應型的植物對變化環境的響應程度的差異是其發生變化的主要原因[19]。植物生態適應性是在漫長的歷史演變過程中通過不斷進化而形成的相對固定的一種特性,并在其結構和生理等方面產生了一系列適應性特化[20]。水生植物只所以能夠適應缺氧環境,其原因在于其根、莖、葉之間已形成了一整套通氣系統。也有一些水生植物能夠貯存自身呼吸釋放的CO2和光合作用釋放的O2,來滿足自身光合和呼吸代謝的需要[21]。而大多數陸生植物缺乏這種組織結構和功能,長期深水淹沒下植物死亡的主要原因是體內缺氧而導致的窒息死亡。一般認為,植物對漸變的環境,可以通過自身的形態變異或生理過程調節來增加其與環境的適合度[22],但這種“變異”和“調節”需經歷一個漸變過程,對消落帶原生植物而言,首次經歷水庫水位漲落完全是一種瞬時、偶發的極端環境變化。在這種突變的環境條件下,植物的“變異”和“調節”能力是極為有限的。因此,植物對首次經歷水庫水位漲落的反應更為強烈。經歷水庫水位漲落后消落帶上出現的“新”植物,并非是植物與環境長期協同進化意義上的“新種”。 其中,有些植物種可能來自鄰近區域,而更多的可能是在經歷水庫水位漲落前以種子庫的形式在消落帶存在的植物種。
經過多次水位漲落后成為消落帶優勢種的生活史對策大多具有r對策種群特征。主要表現是營養生長期相對較短,植物個體較小,初次結實的時間到來較早,能產生大量種子,種子的體形小,可借助風力傳播,并迅速占領由于種種原因形成的空曠地[20]等。同時,與Baker提出的理想雜草(the ideal weed)特征非常吻合。 比如能不連續的發芽(自我控制)且種子能在較長時間內保持生命力;幼苗生長迅速;開花前無性生長期短;只要條件允許則能維持連續的種子生產;能在廣泛的環境中結實,對氣候和土壤變異具有較強的忍耐性(可塑性常常高),對長距離和短距離傳播都有特殊的適應方式;如果是多年生植物,則具有可進行無性繁殖的根莖或根狀莖[23]。毛馬唐、狗尾草、鬼針草、碎米莎草等優勢種多屬于田間和坡地常見的一年生草本。毛馬唐自然生長多見于路旁、田野。喜濕、喜光, 種子耐受水淹的能力較強;狗尾草多生于林邊、山坡、路邊和荒蕪的園地及荒野,對干燥生境有較強的適應能力;鬼針草多生長在路邊荒地、山坡及田間,碎米莎草多生于田間、山坡、路旁陰濕處,為秋熟農田的主要雜草。這些植物都具有生活周期短,能在消落帶成陸期獲得成熟的種子,且種子細小,容易隨風和水體四處擴散。鬼針草的瘦果上還具倒刺毛,能沾鉤在動物體表進行種子傳播。這些植物的種子都能夠以休眠的形式越冬,并在適宜溫度和水分條件滿足時快速萌發完成早期定居過程,進而提高其在植物中的競爭力[24-25];而當萌發條件不具備時,還能夠推遲萌發來分散其在不可預測環境中的風險性[26-27],增加幼苗在更適宜的環境條件下出現和生長的機會[28]。狗牙根為多年生草本,具有能夠克隆繁殖的匍匐狀地下根莖。據野外調查,經歷水深2—7 m,淹水100d左右和伏旱季節后,狗牙根的種群密度和生物量不但沒有降低,反而比對照還有所提高;在經歷水深22—27 m,淹水250d左右的海拔區段,其種群的增殖能力還能維持在較高水平[29]。鹽膚木和算盤子為在消落帶適應性較強的灌木樹種。這兩種植物自然分布在山坡、溝谷雜木疏林和灌叢中。鹽膚木的萌芽更新能力較強。種皮堅硬,表面具蠟質和油質,在生產上用未經處理的種子播種一般難以發芽,需要進行層積催芽處理。湖北算盤子,多生于山坡灌叢中,耐干旱瘠薄,果期6—9 月,與消落帶成陸期一致。目前對這兩種木本植物的適應機制還缺乏深入研究,初步分析認為,除與其耐干旱瘠薄,能夠在消落帶成陸期結實以及特殊的種子結構有關外,長時間淹水對于消除其種子萌發障礙可能還具有一定的促進作用。
3.2 物種多樣性和植物生活型組成的時空變化分析
消落帶海拔156—172 m區段的植物群落經歷了4 次水庫水位漲落,而海拔172—175 m區段僅經歷了2次水庫水位漲落;海拔156—172 m區段每年持續水淹時間為105—243 d,淹水深度在3—19 m,而海拔172—175 m區段每年持續水淹時間約30d左右,淹水深度在0—3 m。雖然不同海拔區段的植物受到水庫水位漲落干擾的強度和頻次不同,但物種多樣性變化總體呈下降的趨勢是一致的。這與王強等在三峽水庫蓄水初期研究消落帶植物物種多樣性空間分布格局的結論相吻合[9]。一般認為,一個較成熟的群落往往具有較高的物種多樣性、較高的均勻度和較低的生態優勢度。如果植物群落朝正向演替(或正向波動),生態系統的物質和能量可得到進一步累積,生物多樣性會相應提高;相反,若植物群落朝逆向演替(或負向波動),生物多樣性勢必下降[30]。根據本研究結果分析認為,目前,三峽庫區消落帶植物群落仍處在逆向演替的早期階段,群落的物種組成和結構仍處在不穩定狀態。
不同調查時期、不同海拔區段消落帶植物生活型組成總的變化趨勢是:喬木、灌木和藤本減少,草本增加。這與前人預測和調查的結果相一致[1, 6]。植物生活型是不同植物對相同生境趨同適應的外在表現[31]。植物生活型對水陸生境變化的響應策略,決定了植物群落的物種組成和替代變化趨勢,也是消落帶植物群落在水陸生境變化下演替的基礎。在三峽水庫反復周期性水位漲落影響下,草本植物(包括一年生和多年生植物)生活型取代喬木、灌木和藤本生活型是三峽庫區消落帶植物生活型組成變化的必然趨勢。
3.3 奉節以東巫山和秭歸段與其他區域消落帶植物群落動態特征的比較
三峽水庫奉節以東巫山和秭歸段庫岸山高坡陡,森林資源豐富,且植被的自然度較高,而中部和西南部地區的庫岸地勢較平緩,以低山丘陵為主,因開發較早,海拔600m以下大多被開墾為農耕地,自然植被已遭到嚴重破壞。對消落帶植物群落的動態而言,水文條件變化是影響其最主導的因素,而植物區系、地形、土壤、人為干擾等也具有重要影響。然而以往的研究往往規避或未能充分考慮這些因素[10]。王強等于2008—2010年監測位于奉節西部的開縣典型消落帶植被的結果表明,在坡度15°的水田和旱地撂荒地60m×30m的樣地上,經歷水庫水位漲落影響后,不同調查年份的優勢植物有蒼耳(Xanthiumsibiricum)、雙穗雀稗(Paspalumpaspaloides)狗牙根、狗尾草等[9];王業春等在庫區西南部的忠縣,選擇3 個地質地貌和土地利用歷史相似的近自然消落帶(土地利用歷史為農用耕地),對不同水位高程(海拔160m和170m)植物群落和土壤特征的研究結果表明,不同水位高程的植被物種組成完全相同,且蓋度、生物量、生物多樣性指數均沒有顯著差異,只是群落優勢種存在差異[10]。與本研究結果相比,奉節以東巫山和秭歸段消落帶的植物組成和優勢種以及群落結構的變化等都與上述研究區域有所不同。另外,奉節以東巫山和秭歸段消落帶的坡度較大,土壤沖刷嚴重。土壤是植物生存和分布的基礎條件,土壤流失速度快,消落帶植物群落變化的速度也必然會相應加快。
3.4 消落帶植被動態研究方法分析
獲取經歷水庫水位漲落前消落帶植物群落組成和結構的初始資料后,對經歷水庫水位漲落影響后植物的“消長”動態定位“跟蹤”,再根據不同植物多年的“消長”變化趨勢,確定用于消落帶植被恢復的備選植物,較之“以空間代替時間”和一次性調查得出的結果具有一定的可靠性;同時,通過對不同植物和群落結構變化的長期監測,即可獲得消落帶植物群落演替的確鑿證據。三峽水庫運行初期是消落帶植物群落種類組成和結構快速變化時期,在此時期,定位監測具有其它方法不可替代的重要意義。
固定樣地監測的不足在于很難反映較大地理空間尺度上消落帶植物群落的宏觀變化趨勢,而且受人力、物力的限制,很難做到大范圍的定位觀測。如何將固定樣地監測與遙感數據的宏觀監測相結合揭示較大地理空間尺度上消落帶植物群落的動態變化規律還有待深入研究。
致謝:野外調查得到秭歸縣和巫山縣林業局的大力支持,劉正宇研究員、劉濤老師,周火明副教授、程瑞梅研究員、金江群、巴哈爾古麗、康義、王曉榮、王祥福等碩士研究生參加了部分年度調查,特此致謝。
[1] 胡波, 張平倉, 任紅玉, 羅慧敏, 岑奕. 三峽庫區消落帶植被生態學特征分析. 長江科學院院報, 2010, 27(11):81- 85.
[2] 熊俊, 袁喜, 梅朋森, 張麗萍, 許文年, 黃應平. 三峽庫區消落帶環境治理和生態恢復的研究現狀與進展. 三峽大學學報(自然科學版), 2011, 33(2):23- 28.
[3] 郭泉水, 康義, 洪明, 金江群, 朱妮妮, 聶必紅, 王佐慶. 三峽庫區消落帶陸生植被對首次水陸生境變化的響應. 林業科學, 2013, 49(5):1- 9.
[4] 黃世能, 王伯蓀. 熱帶次生林群落動態研究:回顧與展望. 世界林業研究, 2000, 13(6):7- 13.
[5] 熊平生, 謝世友, 謝金寧. 初探三峽水庫濕地面臨的問題及其對策. 國土與自然資源研究, 2004, (4):62- 63.
[6] 王勇, 劉義飛, 劉松柏, 黃宏文. 三峽庫區消漲帶植被重建. 植物學通報, 2005, 22(5):513- 522.
[7] 白寶偉, 王海洋, 李先源, 馮義龍, 智麗. 三峽庫區淹沒區與自然消落區現存植被的比較. 西南農業大學學報(自然科學版), 2005, 27(5):684- 691.
[8] 盧志軍, 李連發, 黃漢東, 陶敏, 張全發, 江明喜. 三峽水庫蓄水對消漲帶植被的初步影響. 武漢植物學研究, 2010, 28(3):303- 314.
[9] 王強, 袁興中, 劉紅, 張躍偉, 陳忠禮, 李波. 三峽水庫初期蓄水對消落帶植被及物種多樣性的影響. 自然資源學報, 2011, 26(10):1680- 1693.
[10] 王業春, 雷波, 張晟. 三峽庫區消落帶不同水位高程植被和土壤特征差異. 湖泊科學, 2012, 24(2):206- 212.
[11] 肖文發, 周志翔, 黃志霖, 王鵬程, 吳昌廣. 長江三峽庫區森林景觀格局與景觀恢復研究. 北京:科學出版社, 2012.
[12] Whittaker R J, Willis K J, Field R. Scale and species richness:towards a general hierarchical theory of species diversity. Journal of Biogeography, 2001, 28(4):453- 470.
[13] Wills K J, Whittaker R J. Species diversity-scale matters. Science, 2002, 295(5558):1245- 1248.
[14] 黃忠良, 彭少麟, 易俗. 影響季風常綠闊葉林幼苗定居的主要因素. 熱帶亞熱帶植物學報, 2001, 9(2):123- 128.
[15] 中國長江三峽集團公司水情信息網, 宜昌. http://www.ctg.com.cn/inc/sqsk.php.
[16] 中國植被編輯委員會. 中國植被. 北京:科學出版社, 1980.
[17] 賀金生, 陳偉烈, 李凌浩. 中國中亞熱帶東部常綠闊葉林主要類型的群落多樣性特征. 植物生態學報, 1998, 22(4):304- 311.
[18] 方精云, 王襄平, 沈澤昊, 唐志堯, 賀金生, 于丹, 江源, 王志恒, 鄭成洋, 朱江玲, 郭兆迪. 植物群落清查的主要內容、方法和技術規范. 生物多樣性, 2009, 17(6):533- 548.
[19] 叢靜, 尹華群, 盧慧, 宿秀江, 楊敬元, 李迪強, 張于光. 神農架保護區典型植被的物種多樣性和環境解釋. 林業科學, 2013, 49(5):30- 35.
[20] 李俊清. 森林生態學. 北京:高等教育出版社, 2010.
[21] Mittler R. Oxidative stress, antioxidants and stress tolerance. Trends in Plant Science, 2002, 7(9):405- 410.
[22] 常杰, 葛瀅. 生態學. 杭州:浙江大學出版社, 2001.
[23] Baker H G, Stebbins G L. The Genetics of Colonizing Species. New York:Academic Press, 1965.
[24] Grime JP, Mason G, Curtis AV, Rodman J, Band S R. A comparative study of germination characteristics in a local Flora. Journal of Ecology, 1981, 69(3):1017- 1059.
[25] Bohumil M. Germination requirements of invasive and non-invasiveAtriplexspecies:a comparative study. Flora-Morphology, Distribution, Functional Ecology of Plants, 2003, 198(1):45- 54.
[26] Venable D L, Brown J S. The selective interactions of dispersal, dormancy and seed size as adaptations for reducing risk in variable environments. The American Naturalist, 1988, 131(3):360- 384.
[27] Philippi T. Bet-hedging germination of desert annuals:variation among populations and maternal effects inLepidiumlasiocarpum. The American Naturalist, 1993, 142(3):488- 507.
[28] Kevin J R, Andrew R D. Seed aging, delayed germination and reduced competitive ability inBromustectorum. Plant Ecology, 2001, 155(2):237- 243.
[29] 洪明, 郭泉水, 聶必紅, 康義, 裴順祥, 金江群, 王祥福. 三峽庫區消落帶狗牙根種群對水陸生境變化的響應. 應用生態學報, 2011, 22(11):2829- 2835.
[30] 李瑞. 植被動態研究進展及展望. 中國水土保持科學, 2012, 10(2):115- 120.
[31] 蔣有緒, 郭泉水, 馬娟. 中國森林群落分類及其群落學特征. 北京:科學出版社, 1998.
Plant community dynamics in the Hydro-fluctuation Belt of the Three Gorges Reservoir at the Zigui and Wushan Section, East of Fengjie County, China
ZHU Nini1,GUO Quanshui1,*,QIN Aili1,PEI Shunxiang2,MA Fanqiang1,ZHU Li1,3, JIAN Zunji1
1ResearchInstituteofForestEcology,EnvironmentandProtection,ChineseAcademyofForestryKeyLaboratoryofForestEcologyandEnvironmentofStateForestryAdministration,Beijing100091,China2ExperimentalCentreofForestryinNorthChina,ChineseAcademyofForestry,Beijing102300,China3CollegeofAgriculture,HenanUniversityofScienceandTechnology,Luoyang471003,China
The plant community growing in the hydro-fluctuation belt is an important component of reservoir management, playing a significant role in the maintenance of riparian ecosystems and ecological functioning. Obtaining timely and accurate data on plant community dynamics could provide theoretical and practical information for vegetation restoration and reconstruction in the Three Gorges Reservoir. This study collected data over 5 years (2008—2012) for the plant community from four permanent monitoring plots at different elevations in Zigui and Wushan section of the Three Gorges Reservoir (East of Fengjie County). The dynamic characteristics, plant community composition, dominant plants, plant life forms, and species diversity were studied. The results indicated that compared to 2008, the elevation from 156 to 172 m was subject to water level fluctuation on one occasion by 2009 and four occasions by 2012. The number of native plants declined from 147 species in 2008 (belonging to 55 families) to 33 species (belonging to 18 families) by 2009; the rate of reduction for families and species was 67.3%and 77.6%, respectively. The number of native plants declined to 39 species (14 families) by 2012, with the rate of reduction for families and species being 74.5%and 73.5%, respectively. However, 49 (by 2009) and 23 (by 2012) “new” species appeared during the study period, raising the total number of species by 59.8%and 32.9%, respectively, in certain sampled plots. The elevation from 172 to 175 m was subject to water level fluctuations on two occasions by 2012. The number of native plants declined from 91 species (40families) to 20species (13 families). The rate of reduction was 67.5%and 78.0%, respectively. However, 21 “new” species accounted for about 44.7%of all sampled plant species. Based on the comparison of several survey results on the number of “disappeared” and “appeared” plant species, the results indicated that the most sensitive response of the plant community was to the first water level fluctuation. Subsequently, even though the plant community was subject to several water level fluctuations, the rate of change declined. The “growth” dynamics of ecologically adapted plants and the composition of dominant species differed at different altitudes. At an altitude of 156—172 m, the dominant herbaceous species wereBidenspilosa(Asteraceae),Cynodondactylon(Poaceae),Digitariachrysoblephara(Poaceae),Setariaviridis(Poaceae), andCyperusiria(Cyperaceae). The dominant shrub species wereRhuschinensis(Anacardiaceae) andGlochidionpuberum(Euphorbiaceae). At an altitude of 172—175 m,Glochidionwilsonii(Euphorbiaceae),Vitexnegundo(verbenaceae) andParthenocissusquniquefolia(Vitaceae) became dominant shrub species, except forBidenspilosa, andDigitariachrysoblephara. Annual and perennial herbs were the dominant plant life-form in the hydro-fluctuation belt. Species diversity decreased as the number of water level fluctuations increased. Periodic fluctuation in the water level of the Three Gorges Reservoir led to flooding-drying habitat alterations. The adaptability of different ecological plants to changing habitats was the main driving factor of plant community change.
plant community dynamics; plant life form; species diversity; The Three Gorges Reservoir; water-level fluctuation belt
林業十二五科技支撐計劃項目(2011BAD38BO4)
2014- 04- 24; < class="emphasis_bold">網絡出版日期:
日期:2015- 05- 19
10.5846/stxb201404240818
*通訊作者Corresponding author.E-mail:guoqs@caf.ac.cn
朱妮妮,郭泉水,秦愛麗,裴順祥,馬凡強,朱 莉,簡尊吉.三峽水庫奉節以東秭歸和巫山段消落帶植物群落動態特征.生態學報,2015,35(23):7852- 7867.
Zhu N N,Guo Q S,Qin A L,Pei S X,Ma F Q,Zhu L, Jian Z J.Plant community dynamics in the Hydro-fluctuation Belt of the Three Gorges Reservoir at the Zigui and Wushan Section, East of Fengjie County, China.Acta Ecologica Sinica,2015,35(23):7852- 7867.
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
成才之路(2017年21期)2017-07-27 20:52:40
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
安徽農學通報(2017年9期)2017-05-19 23:03:05
現代農業科技(2016年22期)2017-03-24 12:21:57
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
學周刊·下旬刊(2015年10期)2015-07-22 12:36:17
