流感病毒多表位核酸疫苗CTB-Eg的構(gòu)建與細(xì)胞轉(zhuǎn)染研究*
2015-02-25 03:05:35寇靜遠(yuǎn)潘興李婉宜鄺玉周琳琳楊靖石新麗陸順順黃筱鈞鄧昭敏秦臻王保寧李明遠(yuǎn)
西部醫(yī)學(xué) 2015年2期
寇靜遠(yuǎn) 潘興 李婉宜 鄺玉 周琳琳 楊靖 石新麗 陸順順黃筱鈞 鄧昭敏 秦臻 王保寧 李明遠(yuǎn)
(1.四川大學(xué)華西基礎(chǔ)醫(yī)學(xué)與法醫(yī)學(xué)院微生物教研室, 四川 成都 610041;2.湖北醫(yī)藥學(xué)院微生物學(xué)教研室, 湖北 十堰 442000)
流感病毒多表位核酸疫苗CTB-Eg的構(gòu)建與細(xì)胞轉(zhuǎn)染研究*
寇靜遠(yuǎn)1潘興1李婉宜1鄺玉1周琳琳1楊靖2石新麗1陸順順1黃筱鈞1鄧昭敏1秦臻1王保寧1李明遠(yuǎn)1
(1.四川大學(xué)華西基礎(chǔ)醫(yī)學(xué)與法醫(yī)學(xué)院微生物教研室, 四川 成都 610041;2.湖北醫(yī)藥學(xué)院微生物學(xué)教研室, 湖北 十堰 442000)
目的 設(shè)計(jì)并構(gòu)建含分子內(nèi)佐劑的流感病毒多表位核酸疫苗CTB-Eg,初步研究其在HEK293T細(xì)胞中的轉(zhuǎn)染效率。方法 通過生物信息學(xué)軟件設(shè)計(jì)并合成了含分子內(nèi)佐劑的流感病毒多表位基因CTB-Eg,將其插入真核載體pEGFP-C2中,獲得了重組質(zhì)粒pEGFP-C2/CTB-Eg;用脂質(zhì)體法轉(zhuǎn)染HEK293T細(xì)胞,通過熒光顯微鏡觀察和PCR法檢測(cè)其轉(zhuǎn)染效率。結(jié)果 成功構(gòu)建了真核表達(dá)質(zhì)粒pEGFP-C2/CTB-Eg,且該重組質(zhì)粒能在HEK293T細(xì)胞中瞬時(shí)轉(zhuǎn)染, 轉(zhuǎn)染效率在50%~70%之間。結(jié)論 本研究成功設(shè)計(jì)、構(gòu)建了CTB-Eg融合基因,其在HEK293T細(xì)胞中轉(zhuǎn)染效率較高,為流感病毒多表位核酸疫苗CTB-Eg在小鼠體內(nèi)的抗病毒作用研究奠定了堅(jiān)實(shí)基礎(chǔ)。
流感病毒; 核酸疫苗; 多表位基因; 霍亂毒素B亞單位
季節(jié)性流感至少影響著全球15% 的人口,每年約有50 萬人死于流感[1~4]。同其他病毒性疾病一樣,流感的防治尚無特別有效的方法,接種疫苗被認(rèn)為是預(yù)防流感發(fā)生與傳播的最佳方法。目前使用的流感疫苗主要是以血凝素(Hem- agglutinin,HA)和神經(jīng)氨酸酶(Neuraminidase,NA)作為靶抗原。然而HA基因很容易發(fā)生變異,針對(duì)特定流感病毒株的疫苗不能對(duì)其他的病毒株產(chǎn)生相應(yīng)的保護(hù)性。因此,研發(fā)能夠誘導(dǎo)廣譜免疫反應(yīng)的通用流感疫苗已經(jīng)成為流感疫苗研究的重要趨勢(shì)[5、6]。本研究將前期設(shè)計(jì)合成的新型流感病毒多表位基因Eg[7]與分子內(nèi)佐劑CTB串聯(lián),連接至真核表達(dá)載體pEGFP-C2中,構(gòu)建了流感病毒多表位核酸疫苗CTB-Eg,并對(duì)其在真核細(xì)胞HEK293T中的轉(zhuǎn)染效率做了初步研究,為流感病毒多表位核酸疫苗CTB-Eg在小鼠體內(nèi)的抗病毒作用研究奠定了堅(jiān)實(shí)基礎(chǔ)。
1 材料與方法
1.1 實(shí)驗(yàn)材料 大腸桿菌DH5α菌株由本教研室保存;真核表達(dá)質(zhì)粒pEGFP-C2購自ATCC(America Type Culture Collection);人胚腎HEK293T(Human embryonic kidney 293)細(xì)胞源自美國ATCC;轉(zhuǎn)染試劑Lipofectamine TM 2000 、膠回收試劑盒和質(zhì)粒提取試劑盒均購自Invitrogen公司;PCR試劑盒(SinoBio 2× Taq Master Mix)、Trizol試劑、T4連接酶、限制性內(nèi)切酶EcoR I和Hind III購自TAKARA公司;青霉素和卡那霉素為北京百泰克生物技術(shù)有限公司;DMEM、RPMI-1640細(xì)胞培養(yǎng)基購自Gibco公司,新生小牛血清購自中美合資蘭州民海生物工程有限公司;96孔培養(yǎng)板、24孔培養(yǎng)板和6孔培養(yǎng)板為Corning 公司產(chǎn)品。
1.2 實(shí)驗(yàn)方法
1.2.1 CTB-Eg的設(shè)計(jì)與合成 為了增加流感病毒多表位核酸疫苗的抗原性及靶向性,選擇兼具神經(jīng)節(jié)苷脂結(jié)合活性及粘膜佐劑活性的霍亂腸毒素B亞單位(CTB)作為分子內(nèi)佐劑,重新設(shè)計(jì)并合成了分子內(nèi)佐劑-流感病毒多表位基因融合疫苗CTB-Eg,并通過ProPred、SWISS-MODEL等生物信息學(xué)軟件,對(duì)CTB-Eg蛋白進(jìn)行了3維建模。目的片段Eg、CTB及CTB-Eg 均由華大基因合成。合成的puc57/CTB、puc57/Eg、puc57/ CTB-Eg質(zhì)粒通過EcoR Ⅰ和Hind Ⅲ酶切位點(diǎn)分別進(jìn)行單、雙酶切鑒定,并回收雙酶切產(chǎn)物。
1.2.2 重組真核表達(dá)質(zhì)粒pEGFP-C2/CTB-Eg的構(gòu)建 pEGFP-C2質(zhì)粒經(jīng)EcoR Ⅰ和Hind Ⅲ雙酶切膠回收后,在T4 DNA連接酶的作用下,分別與目的片段CTB、Eg和CTB-Eg連接,載體與目的片段摩爾比為1∶10。連接產(chǎn)物轉(zhuǎn)化感受態(tài)大腸桿菌DH5α,在含有卡那霉素的LB平板上篩選克隆。挑取陽性克隆,搖菌擴(kuò)增,DNA抽提試劑盒提取質(zhì)粒,并進(jìn)行單、雙酶切鑒定和基因測(cè)序鑒定。
1.2.3 HEK293T細(xì)胞的培養(yǎng)和轉(zhuǎn)染 HEK293T細(xì)胞株于37℃、5%的CO2條件下, 在含10%胎牛血清、100 μg/ml青霉素及100 U/ml鏈霉素的DMEM高糖培養(yǎng)基中進(jìn)行培養(yǎng)。將生長良好的細(xì)胞用上述完全培養(yǎng)基調(diào)至2×105~5×105/ml后按2 ml/孔分裝至6孔板中,待細(xì)胞貼壁生長至大約70%覆蓋密度時(shí)進(jìn)行質(zhì)粒DNA轉(zhuǎn)染。轉(zhuǎn)染采用Lipofectamine TM 2000試劑,按pEGFP-C2、pEGFP-C2/CTB、pEGFP-C2/Eg、pEGFP-C2/CTB-Eg質(zhì)粒DNA 0.2 g/孔、轉(zhuǎn)染試劑1 μl/孔對(duì)細(xì)胞進(jìn)行轉(zhuǎn)染,同時(shí)設(shè)立不進(jìn)行轉(zhuǎn)染的細(xì)胞對(duì)照孔。轉(zhuǎn)染細(xì)胞培養(yǎng)48 h后,在倒置熒光顯微鏡下,利用pEGFP-C2質(zhì)粒自帶的EGFP標(biāo)簽檢測(cè)重組質(zhì)粒的轉(zhuǎn)染效率。
1.2.4 PCR法檢測(cè)重組質(zhì)粒在HEK293T細(xì)胞中的轉(zhuǎn)染情況 分別設(shè)計(jì)CTB、Eg、CTB-Eg引物各一對(duì),引物序列見表1。轉(zhuǎn)染細(xì)胞培養(yǎng)48小時(shí)后,利用普通PCR法檢測(cè)重組質(zhì)粒在HEK293T細(xì)胞中的轉(zhuǎn)染情況。

表1 PCR擴(kuò)增CTB、Eg、CTB-Eg 3者的引物序列
2 結(jié)果
2.1 CTB-Eg結(jié)構(gòu)示意圖及其3級(jí)結(jié)構(gòu) 在前期構(gòu)建的流感病毒多表位基因Eg[7]的N末端添加已去除信號(hào)肽的CTB序列,GGGGS為二者的連接序列,進(jìn)而構(gòu)建了流感病毒多表位核酸疫苗CTB-Eg,見圖1。CTB-Eg三級(jí)結(jié)構(gòu)顯示Linker序列GGGGS、AAY有效地維持了各個(gè)抗原的空間結(jié)構(gòu),避免了抗原表位間的相互干擾,見圖2。
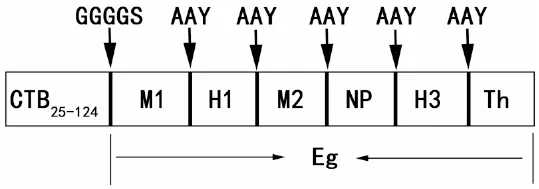
圖1 CTB-Eg構(gòu)建示意圖
Fig. 1 Schematic representation of the designed multi-epitope vaccine CTB-Eg
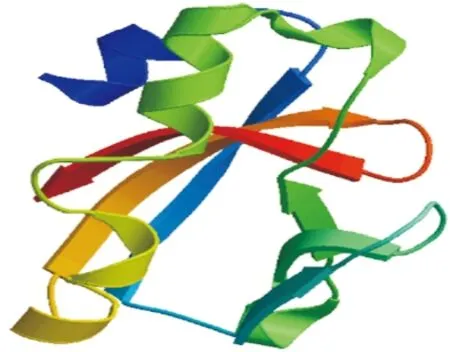
圖2 SWISS-MODEL預(yù)測(cè)CTB-Eg的3級(jí)結(jié)構(gòu)
Fig. 2 Three dimensional (3D) structure information of CTB-Eg.
2.2 CTB、Eg、CTB-Eg目的片段和真核載體pEGFP-C2的鑒定 puc57/CTB、puc57/Eg、puc57/ CTB-Eg及pEGFP-C2質(zhì)粒分別經(jīng)EcoR I和Hind III進(jìn)行單、雙酶切,經(jīng)1.5%的瓊脂糖凝膠電泳,可分別在312 bp、264 bp、579 bp和4700 bp處出現(xiàn)明顯條帶,見圖3,與目的片段及載體pEGFP-C2片段大小一致。
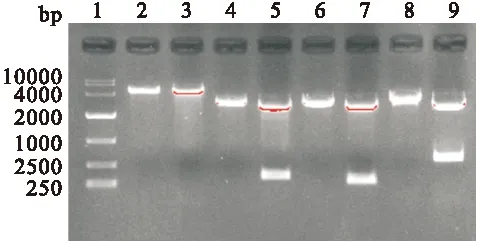
圖3 1.5%瓊脂糖凝膠電泳檢測(cè)目的基因片段大小
Fig.3 Restriction analysis of recombinant plasmids
1: DNA Maker;2、3:pEGFP-C2載體單、雙酶切 ;4、5:puc57/CTB單、雙酶切;6、7:puc57/Eg單、雙酶切;8、9:puc57/CTB-Eg單、雙酶切
2.3 重組質(zhì)粒的酶切鑒定及測(cè)序鑒定 重組質(zhì)粒pEGFP-C2/CTB、pEGFP-C2/Eg、 pEGFP-C2/CTB-Eg 分別進(jìn)行EcoR I、Hind III單、雙酶切鑒定,1.5%瓊脂糖凝膠電泳顯示獲得了312 bp、264 bp、579 bp的插入片段,見圖4。測(cè)序結(jié)果同樣證實(shí)所獲目的片段與設(shè)計(jì)的基因序列完全一致,無堿基突變。
2.4 熒光顯微鏡觀察重組質(zhì)粒轉(zhuǎn)染HEK293T細(xì)胞 重組質(zhì)粒pEGFP-C2/CTB、pEGFP-C2/Eg、 pEGFP-C2/CTB-Eg轉(zhuǎn)染HEK293T細(xì)胞48 h后,熒光顯微鏡觀察EGFP綠色熒光信號(hào)。結(jié)果可見重組質(zhì)粒轉(zhuǎn)染HEK293T細(xì)胞的轉(zhuǎn)染效率達(dá)50%~70%,對(duì)照孔未見熒光,見圖5。
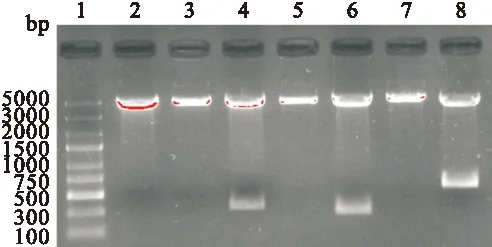
圖4 重組真核表達(dá)質(zhì)粒EcoR I和Hind III單、雙酶切分析
Fig. 4 Restriction analysis of recombinant plasmids.
1: DNA Maker;2:pEGFP-C2載體雙酶切 ;3、4:pEGFP-C2/CTB單、雙酶切;5、6: pEGFP-C2/Eg單、雙酶切;7、8:pEGFP-C2/ CTB-Eg單、雙酶切
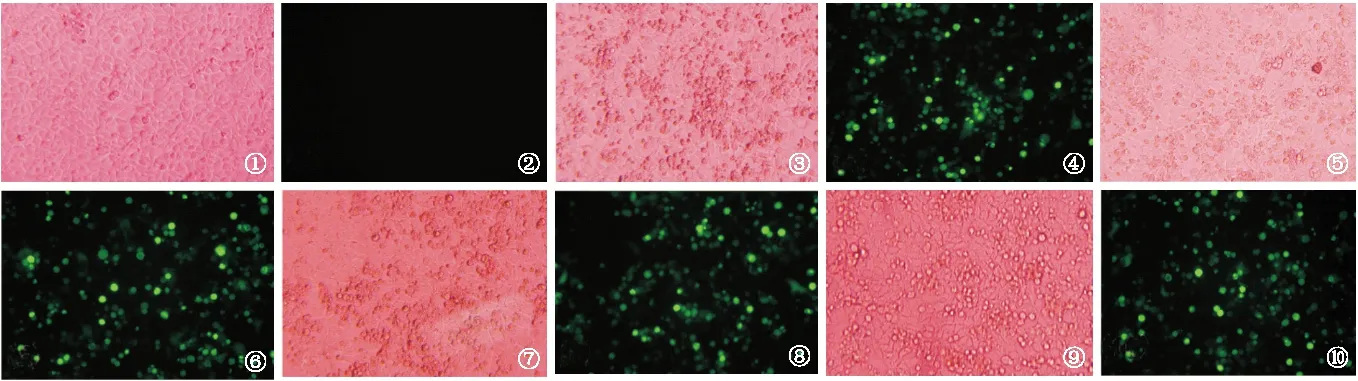
圖5 熒光顯微鏡檢測(cè)各組重組質(zhì)粒在HEK293T細(xì)胞中的轉(zhuǎn)染效率(×200)
Fig.5 Fluorescence microscope detection of the transfection effiencyof recombinant plasmids.
①、②:空白組;③、④:pEGFP-C2組;⑤、⑥:pEGFP-C2/CTB組;⑦、⑧:pEGFP-C2/Eg組;⑨、⑩:pEGFP-C2/CTB-Eg組
2.5 PCR法鑒定重組質(zhì)粒轉(zhuǎn)染HEK293T 重組質(zhì)粒轉(zhuǎn)染HEK293T細(xì)胞48小時(shí)后,利用普通PCR法檢測(cè)重組質(zhì)粒在293T細(xì)胞中的轉(zhuǎn)染情況。經(jīng)1.5%的瓊脂糖凝膠電泳,可分別在312 bp、264 bp和579 bp處出現(xiàn)條帶,與目的片段大小一致,見圖6。
3 討論
流感病毒獨(dú)特的基因結(jié)構(gòu)導(dǎo)致其極易發(fā)生變異,而新病毒株的出現(xiàn)意味著那些接種過傳統(tǒng)流感疫苗的人群對(duì)新出現(xiàn)的流行株同樣敏感。因此,流感疫苗的研究焦點(diǎn)是研發(fā)對(duì)不同亞型流感病毒都具有交叉保護(hù)作用的通用型長效疫苗,而多表位基因疫苗的構(gòu)想有望實(shí)現(xiàn)這一目標(biāo)。
多表位核酸疫苗可在宿主細(xì)胞系統(tǒng)中轉(zhuǎn)錄并表達(dá)多個(gè)不同的保護(hù)性抗原表位,從多個(gè)方面誘導(dǎo)機(jī)體
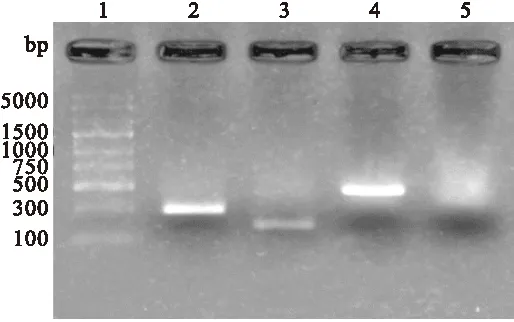
圖6 PCR檢測(cè)各組質(zhì)粒在HEK293T細(xì)胞中的轉(zhuǎn)染情況
Fig. 6 PCR analysis of the transfection of recombinant plasmids.
1:DNA Maker; 2: pEGFP-C2/CTB組;3: pEGFP-C2/Eg組;4: pEGFP-C2/CTB-Eg組; 5: pEGFP-C2載體組
的免疫應(yīng)答,是一種具有廣譜免疫反應(yīng)的通用疫苗。且DNA疫苗在體內(nèi)有長期持久的免疫原性,其表達(dá)蛋白的結(jié)構(gòu)類似于正常真核細(xì)胞的表達(dá)結(jié)構(gòu)[8]。DNA編碼的相關(guān)蛋白作為內(nèi)源性抗原經(jīng)由MHC-Ⅰ類途徑加工處理后,表達(dá)于抗原提成細(xì)胞(antigen presenting cell, APC)表面,進(jìn)一步刺激細(xì)胞毒性T細(xì)胞的活化。抗原肽也可作為分泌性蛋白,通過B細(xì)胞表面抗原受體直接識(shí)別結(jié)合進(jìn)而攝取抗原,并經(jīng)MHC-Ⅱ類途徑加工處理后形成的抗原肽,以抗原肽-MHC-Ⅱ類分子復(fù)合物的形式轉(zhuǎn)運(yùn)到細(xì)胞表面,供CD4+ Th細(xì)胞識(shí)別,從而啟動(dòng)特異性體液免疫應(yīng)答,同時(shí)產(chǎn)生記憶B細(xì)胞參與二次免疫,保護(hù)宿主免受相關(guān)微生物的攻擊[9]。
Graeme等研究表明NP和M2的表達(dá)可顯著降低流感病毒在小鼠之間的傳播[10]。Donnelly等構(gòu)建了含有不同流感病毒株HA、M1和NP的融合DNA疫苗,該疫苗可以有效的提高小鼠的免疫保護(hù)率[11]。還有研究證實(shí),含有HA基因的DNA疫苗加上NP或M基因,被免疫的小鼠可以抵抗不同亞型的流感病毒[12,13],其中M1或NP基因能提高疫苗的保護(hù)性并使免疫小鼠抵抗不同亞型流感病毒的攻擊,而M2基因?qū)γ庖咝∈笕狈τ行У谋Wo(hù)作用[14]。流感病毒感染人體后,只有1%~2%的人會(huì)產(chǎn)生抗M2e抗體,該抗體可能通過與感染細(xì)胞結(jié)合,介導(dǎo)CD+8 T細(xì)胞的殺細(xì)胞過程[15]。本研究從HA蛋白選擇了2個(gè)B細(xì)胞表位,分別在NP、M1、M2蛋白選擇了3個(gè)T細(xì)胞表位。既保留HA表位良好的抗原性及誘導(dǎo)中和抗體的能力,又可能通過高度保守的M、NP蛋白克服HA高度變異所導(dǎo)致的疫苗交叉保護(hù)性弱的問題。為防止DNA疫苗在體內(nèi)非特異性降解而導(dǎo)致的效率減弱及各表位間相互抑制的問題,課題中我們將所選取的5個(gè)表位通過不同排列方式串聯(lián)組合,通過生物信息學(xué)分析評(píng)估,選取最佳組合方式,加上適當(dāng)?shù)拈g隔序列設(shè)計(jì)成流感通用多表位基因疫苗Eg。
霍亂毒素(cholera toxin,CT)是最常用的免疫佐劑之一,由霍亂弧菌(Vibrio cholerae)分泌,分子量大小為84 kD,具有很強(qiáng)的免疫原性和粘膜免疫佐劑性。CT的結(jié)構(gòu)為一個(gè)A亞單位、五個(gè)B亞單位。CTB分子量大小為11.6 kD,五分子的CTB通過鹽鍵和氫建聚集成56 kD的大亞基,呈圓筒狀,與真核細(xì)胞表面的神經(jīng)節(jié)苷脂GM1結(jié)合,可使連接的抗原與粘膜作用,進(jìn)而引起一系列的生化反應(yīng),產(chǎn)生更強(qiáng)的免疫效果[16]。CTB可促進(jìn)抗原通過粘膜屏障,加強(qiáng)抗原被樹突狀細(xì)胞和其他抗原提呈細(xì)胞的提呈作用,增強(qiáng)抑制性T細(xì)胞分泌TGF-β。重組CTB去除了CTA的毒性,保留了霍亂毒素的免疫佐劑性,從而使機(jī)體產(chǎn)生較強(qiáng)的系統(tǒng)免疫應(yīng)答和局部黏膜免疫應(yīng)答[17,18]。因此,在多表位核酸疫苗的設(shè)計(jì)中,我們?cè)谄銷末端添加霍亂毒素B亞單位(cholera toxin B subunit, CTB)作為分子內(nèi)佐劑,期望能增強(qiáng)疫苗的免疫效果。
4 結(jié)論
本實(shí)驗(yàn)將流感多表位基因Eg與分子內(nèi)佐劑CTB串聯(lián),連接至真核表達(dá)載體pEGFP-C2中,構(gòu)建了流感病毒多表位核酸疫苗CTB-Eg,并對(duì)其在真核細(xì)胞HEK293T中的轉(zhuǎn)染情況做了初步研究,為下一步流感病毒多表位核酸疫苗CTB-Eg在小鼠體內(nèi)的抗流感作用研究奠定了實(shí)驗(yàn)基礎(chǔ)。
[1]Meng S, L.Z., Xu L,etal. Intranasal immunization with recombinant HA and mast cell activator C48/80 elicits protective immunity against 2009 pandemic H1N1 influenza in mice[J]. PLoS One, 2011, 6(5): e19863.
[2]Ferraris O, E.V., Bouscambert M,etal. H1N1 influenza A virus neuraminidase modulates infectivity in mice[J]. Antiviral Res, 2012, 93(3): 374-380.
[3]Imai M, Watanabe T, Hatta M,etal. Experimental adaptation of an influenza H5 HA confers respiratory droplet transmission to a reassortant H5 HA/H1N1 virus in ferrets[J]. Nature, 2012, 486(7403): 420-428.
[4]Seasonal influenza: World Health Organization, Copenhagen, Denmark (2009). (www.euro.who.int/influenza/2008618_1).
[5]Girard MP, Cherian T, Pervikov Y,etal. A review of vaccine research and development: human acute respiratory infections[J]. Vaccine, 2005, 23 (50): 5708-5724.
[6]陳則.通用流感疫苗進(jìn)展研究[J]. 湖南師范大學(xué)學(xué)報(bào)(醫(yī)學(xué)版),2012, 9 (3):1-5.
[7]邵京京,豐鋒,張強(qiáng),等.流感病毒多表位基因盒的設(shè)計(jì)與預(yù)測(cè)[J].華西藥學(xué)雜志,2012,27(005):486-489.
[8]Alarcon JB, Waine GW, McManus DP. DNA vaccines: technology and application as anti-parasite and anti-microbial agents[J]. Adv Parasitol, 1999, 42: 343-410.
[9]Kishwar Hayat Khan. DNA vaccine: roles against diseases[J]. Germs, 2013, (3)1: 27-35.
[10] Graeme E. Price, Chia-Yun Lo,etal. Mucosal Immunization with a Candidate Universal Influenza Vaccine Reduces Virus Transmission in a Mouse Model[J]. Journal of Virology, 2014, 88(11): 6019-6030.
[11] Donnelly JJ, Friedman A, Ulmer JB,etal. Further protection against antigenic drift of influenza virus in a ferret model by DNA vaccination[J]. Vaccine, 1997, 15(8): 865-868.
[12] Chen Q, Kuang H, Wang H,etal. Comparing the ability of a series of viral proteinexpressing plasmid DNAs to protect against H5N1 influenza virus[J]. Virus Genes, 2009, 38(1): 30-38.
[13] Chen Z, Yoshikawa T, Kadowaki S,etal. Protection and antibody responses in different strains of mouse immunized with plasmid DNAs encoding influenza virus haemagglutinin, neura-minidase and nucleoprotein [J]. J Gen Virol, 1999, 80(10): 2559-2564.
[14] M. Zheng, J. Luo, Z. Chen. Development of universal influenza vaccines based on influenza virus M and NP genes[J]. Infection, 2014, 42(2): 251-262.
[15] Oxford JS. Towards a universal influenza vaccine: volunteer virus challenge studies in quarantine to speed the development and subsequent licensing[J]. Br J Clin Pharmacol, 2013, 76(2): 210-216.
[16] Hiramatsu Y, Yamamoto M, Satho T,etal. Recombinant fusion protein of cholera toxin B subunit with YVAD secreted by Lactobacillus casei inhibits lipopolysaccharide-induced caspase-1 activation and subsequent IL-1 beta secretion in Caco-2 cells[J]. BMC Biotechnol, 2014, 14(1): 38.
[17] Sun JB, Raghavan S, Sjoling A,etal. Oral tolerance induction with antigen conjugated to cholera toxin B subunit generates both Foxp3+ CD25+ and Foxp3+ CD25+ CD4+ regulatory T cells[J]. Immunol, 2006, 177(11): 7624-7644.
[18] Luci C, Hervouet C, Rousseau D,etal. Dendritic cell-mediated induction of mucosal cytotoxic responses following intravaginal immunization with the nontoxic B subunit of cholera toxin[J]. Immunol, 2006, 176(5): 2749-2757.
Construction and transfection of a multi-epitope DNA vaccine CTB-Eg of influenza
KOU Jing-yuan1, PAN Xing1, LI Wan-yi1,etal
(1.DepartmentofMicrobiology,WestChinaSchoolofPreclinicalandForensicMedicine,SichuanUniversity,Chengdu610041,China; 2.DepartmentofMicrobiology,HubeiUniversityofMedicine,Shiyan442000,China)
Objective To design and construct the eukaryotic expression vector of CTB-Eg fusion gene of influenza virus, and study the transfection efficiency of pEGFP-C2/CTB-Eg in HEK293T cells. Methods CTB-Eg fusion gene containing CTB (cholera toxin B subunit) as the molecule adjuvant was designed and synthesized. The eukaryotic expression vector of pEGFP-C2/CTB-Eg was constructed. The certified recombinant plasmid was transfected in HEK293T cells by Liposomal Transfection Reagent. The transfection efficiency of CTB-Eg gene was detected by fluorescence microscope and PCR. Results Expression vector pEGFP-C2/CTB-Eg was constructed successfully. The recombinant plasmid could be transfected in HEK293T cells and its transfection efficiency could reach to 50%-70%. Conclusion The eukaryotic expression vector of pEGFP-C2/CTB-Eg was constructed successfully and transfected in HEK293T cells.
Influenza virus; DNA vaccine; multi-epitope genes; Cholera toxin B subunit
四川省科技廳支撐計(jì)劃項(xiàng)目(2014SZ0036)
李婉宜,教授,E-mail:hxliwanyi@126.com
R 181.2+4
A
10.3969/j.issn.1672-3511.2015.02.007
2014-07-11; 編輯: 張文秀)
