長江水系中華絨螯蟹野生和養殖群體選育子一代養殖性能和性腺發育的比較*
2015-03-08 05:27:24吳旭干龍曉文趙恒亮姜曉東成永旭
海洋與湖沼 2015年4期
何 杰 吳旭干 龍曉文 劉 青 趙恒亮姜曉東 成永旭①
(1. 上海海洋大學 水產種質資源發掘與利用教育部重點實驗室 上海 201306;2. 上海海洋大學 上海高校知識服務平臺水產動物遺傳育種中心 上海 201306)
中華絨螯蟹(Eriocheir sinensis)俗稱河蟹、大閘蟹,隨著20世紀80年代人工育苗技術取得突破以來,中華絨螯蟹養殖規模迅速擴大,全國絕大多數省份都已經開展人工增養殖,2012年全國養殖總產量達714400t (王武等,2013)。目前,我國中華絨螯蟹養殖主要集中于長江流域,因此長江水系中華絨螯蟹的種源質量對于中華絨螯蟹養殖產業具有極其重要的作用(Suiet al,2011; 張世勇等,2013)。但是,在中華絨螯蟹人工繁育過程中,許多育苗場為了降低親本費用,通常采用小規格親本(雌: 60—100 g/只,雄:80—150 g/只)進行育苗,加上不同水系間盲目引種等原因,致使長江水系中華絨螯蟹養殖群體種質退化嚴重,一齡性早熟率越來越高,商品蟹規格越來越小(Suiet al,2011; 王武等,2013)。因此,開展長江水系中華絨螯蟹優異種質資源的挖掘利用和良種選育勢在必行(朱清順等,2012; 張世勇等,2013)。
近 10年來,江蘇淡水水產研究所通過群體選育的方法已選育出生長速度較快的長江水系中華絨螯蟹良種(朱清順等,2012; 鄧燕飛等,2013),這在一定程度上推動了中華絨螯蟹養殖產業的可持續發展,但迄今為止,中華絨螯蟹的良種培育尚缺乏對性腺成熟時間、抗病力和營養品質等經濟性狀的足夠關注和研究。由于性腺和肝胰腺是中華絨螯蟹重要的可食部分,因此中華絨螯蟹商品蟹主要為性腺發育成熟或基本成熟的成蟹(Wuet al,2011),長江水系中華絨螯蟹性腺發育成熟或接近成熟的時間為 10—12月,過早或過晚捕撈均會影響其食用價值,因此,其銷售及食用時間主要集中于10月底至12月初(滕煒鳴等,2008)。中華絨螯蟹銷售時間集中,使得商品蟹銷售市場競爭過度,價格波動較大,這不僅使得中華絨螯蟹養殖的經濟風險較大,且不利于其均衡上市,這給中華絨螯蟹養殖產業的健康發展帶來了一定的負面影響。因此,不同性腺成熟時間的中華絨螯蟹品種培育顯得十分重要和迫切。有研究表明,通過遺傳育種可以培育動植物的早熟和晚熟品種,在一定程度上實現均衡上市,提高產品的市場競爭力(韋鳳英等,2002;李小紅等,2009; 謝宗周等,2011; 冀曉昊等,2014),這為中華絨螯蟹的良種培育提供了思路和依據。
研究表明,長江水系野生扣蟹在池塘養成過程中不僅商品蟹規格大和成活率高,且具有晚熟特性;而長江水系經過多代池塘養殖的人工繁殖群體通常具有二齡早熟特性(成蟹養殖過程中生殖蛻殼和性腺發育時間提前),這暗示有可能通過遺傳育種手段培育中華絨螯蟹二齡早熟和晚熟品系(Heet al,2014)。因此,有必要研究長江水系中華絨螯蟹野生和養殖群體選育子代是否仍然能保持二齡早熟和晚熟特性。鑒于此,本文系統研究了長江水系野生和養殖群體選育子一代(G1)及非選育群體在扣蟹和成蟹階段的養殖性能和性腺發育情況,旨在評估兩個選育群體子一代在池塘養殖條件下的綜合養殖性能和選育進展,為進一步開展中華絨螯蟹種質資源評價和良種選育提供科學依據和實踐參考。
1 材料與方法
1.1 選育群體幼體來源
野生中華絨螯蟹成蟹于2010年11—12月捕自長江江蘇揚中至丹徒河段(32°11′8.57′N,119°50′1.44′E至 32°06′35.80′N,119°29′14.12′E),從中挑選規格較大、背甲額齒尖銳、疣突明顯、體質健壯的成蟹作為野生群體選育親本; 池塘養殖群體成蟹購自上海崇明雨池水產養殖合作社,從該合作社的 4hm2池塘共產出的約30000只蟹中挑選2400余只(留種率為8%)規格相對較大,且背甲額齒尖銳、疣突明顯的健康個體作為養殖群體選育親本,對照組購自江蘇興化水產養殖個體戶,挑選規格接近、體質健壯的個體作為親本參與繁殖(三群體親本數量及規格見表1)。12月中旬將所有親本運往遼寧盤錦光合蟹業有限公司如東育苗基地分池交配和育苗,分別獲得野生選育 G1代(野選G1)、養殖選育G1代(養選G1)和對照組幼體。5月中旬將培育出的三群體大眼幼體運往上海海洋大學崇明基地分池養殖至豆蟹,期間投喂配合飼料,待7月上旬,從三群體中分別挑選規格相近、十足健全的豆蟹備用于扣蟹階段養殖性能比較[野選G1、養選G1、對照組豆蟹平均體重分別為(0.88±0.13)g、(0.86±0.11)g 和(0.87±0.15)g]。
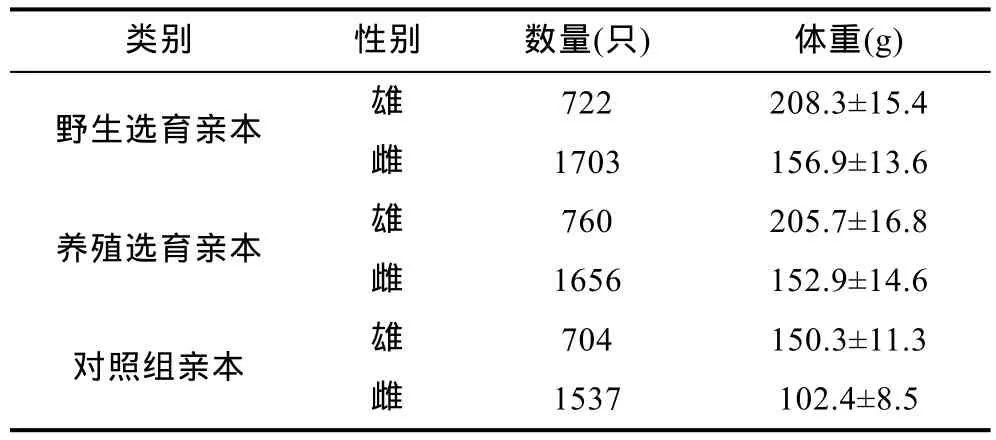
表1 野生選育、養殖選育及對照組參與繁殖親本的數量及規格Tab.1 The number and size of the adult E. sinensis which participate in the breeding
1.2 扣蟹實驗池塘及養殖管理
扣蟹實驗池塘為 12個等大的養殖圍隔,單個圍隔呈“回”字形,總底面積約60m2(長×寬=7.8m×7.8m),圍隔中間為一水坑,長×寬×深=6m×4m×0.7m 供其高溫期棲息,水坑四周為平臺,平臺上種植水稻,每圍隔四周設置雙層防逃網,防逃網上口設置 25cm高的防逃塑料板,每個圍隔分別放養對應的豆蟹 5000只(雌雄各半),每群體各設 4個重復圍隔,各圍隔之間水流相通,水質基本一致。各圍隔內分別種植一定量的水花生供扣蟹隱蔽,同時有利于降低夏季圍隔內的水溫,圍隔內水花生分布和數量基本相似。養殖期間每日下午5: 30投喂一次扣蟹配合飼料(由浙江欣欣飼料有限公司生產),投喂量約占蟹體重的 1%—6%,具體根據水溫和攝食情況靈活調整,每隔15d左右用聚維酮碘對池塘水體消毒一次,根據水質情況適當換水,確保水質良好。此外,定期梳理水花生,使水花生密度適中,防止瘋長造成扣蟹缺氧和水質惡化。
1.3 成蟹實驗池塘及養殖管理
2012年2月,在同一基地選擇兩口池塘進行成蟹養殖實驗。首先采用塑料網片將該池塘隔成12個面積相等的圍隔(長×寬=35m×6m),每個圍隔均采用雙層網片防逃,圍網底部采用石籠連接后埋入土下30cm左右,網頂部兩側縫上35cm高的塑料防逃板防止不同圍隔間的河蟹混雜。2月中旬使用生石灰對實驗池塘進行消毒和清塘,2周后在各圍隔內種植伊樂藻供河蟹隱蔽,伊樂藻株距為 1.5m,行距為 2m。3月 20日左右,從三群體扣蟹實驗池塘中挑選規格相近的扣蟹[野生群體的雌雄平均體重分別為(7.86±2.15)g和(7.39±1.90)g,養殖群體的雌雄平均體重分別為(7.68±1.66)g和(7.23±1.70)g,對照組群體的雌雄平均體重為(7.41±1.69)g和(7.25±1.85)g]。用 30mg/L的聚維酮碘溶液浸泡消毒0.5h后,隨機放養于12個圍隔,每群體重復 4個圍隔,每圍隔雌雄各放養 100只扣蟹。整個養殖期間合理控制水位和水草數量,確保水草生長良好,密度適宜,當池塘水溫升高至12°C以上時開始投喂人工配合飼料,每天一次,通過檢查食臺上的殘餌情況調整投喂量; 6月下旬起在夜間對各圍隔水體進行底部微孔增氧,定期檢測水質,每周換水 1—2次,保證水質維持在適宜中華絨螯蟹生長的正常范圍內。
1.4 數據采集
1.4.1 扣蟹養殖階段生長性能、存活率、產量和飼料系數 2011年7月份起,每月15日前后采樣測量蟹的生長情況,用操網隨機從每個圍隔中各采樣150—200只個體,雌雄各占一半左右,用毛巾輕輕擦拭蟹體表水分后,采用電子天平精確稱重(精確到0.01 g),據此計算增重率和特定生長率:

其中,Wt指第t月蟹的平均重量,W(t–1)指第t–1 月蟹的平均重量,D為采樣的間隔時間。
扣蟹生長實驗于11月15日停止,排水干塘后捕捉所有的蟹,雌雄分開,分別統計存活率、早熟率、產量以及總體飼料系數,其中飼料系數=消耗飼料總重/(最終產出蟹總重–最初放養扣蟹重)。
1.4.2 成蟹養殖階段生長性能、性腺發育速度、存活率、產量和飼料系數 自2012年4月份到10月份,每月20日左右從各養殖圍隔內徒手或用地籠隨機抓取 30—40只個體,逐個測量體重,計算各月的增重率和特定生長率。為了比較三群體扣蟹在成蟹養殖階段的性腺發育速度,在 8—10月份的體重測量時,逐個記錄完成生殖蛻殼完成情況,計算各月的生殖蛻殼完成率,并且從各圍隔內隨機取出 6只蟹(雌雄各半),首先擦去蟹體表水分,精確稱量后(精確到0.01 g)進行活體解剖,取出全部肝胰腺和性腺,稱重(精確到 0.001g)各組織后按照如下公式,分別計算肝胰腺指數(Hepatosomatic index,HSI,%)和性腺指數(Gonadosomatic index,GSI,%):

式中,WH指肝胰腺重,WG指性腺重,W指體重。
10月 20日起,在岸上徒手抓捕或用地籠捕撈,分別統計每個圍隔的雌雄個體成活率、產量,同時計算成蟹養殖階段的飼料系數,計算方法同上。
1.4.3 成蟹平均規格和規格分級 根據最終每個圍隔中捕撈的所有成蟹分別計算兩群體雌雄成蟹的平均體重。參照 He 等(2014)的方法,對最終捕獲的所有成蟹按體重進行規格分級,雄蟹分為7級: ≥250g,225—249.9g,200—224.9g,175—199.9g,150—174.9g,125—149.9g和<125g; 雌蟹分為 5級: ≥150g,125—149.9g,100—124.9g和 <75g,分別統計三群體養成的成蟹各規格等級所占比例。
1.5 數據分析
所有數據采用平均值±標準差表示。采用SPSS13.0軟件對實驗數據進行統計分析,用 Levene方法進行方差齊次性檢測,當不滿足使用齊性方差時進行反正弦或平方根處理,用T檢驗(Independent samplest-test)檢查三群體各指標間的差異性,采用 Sigmaplot10.0軟件繪圖,取P<0.05為差異顯著。
2 結果
2.1 扣蟹階段的養殖性能
2.1.1 生長性能 野選 G1、養選 G1及對照組在扣蟹養殖階段的體重增長趨勢見圖 1,8月份三群體的體重無顯著差異,自9月份起野選G1的體重始終低于對照組,其中9月份顯著低于對照組,而養選G1在 10月份起的體重略高于對照組并顯著高于野選G1(P<0.05); 隨著養殖時間的延長,三群體扣蟹的增重率(WGR)整體上均呈下降趨勢(圖2),僅在8—9月間,野選 G1顯著低于對照組,其它時間段內三群體間的WGR并無顯著差異(P>0.05); 三群體的SGR變化與WGR的變化趨勢類似(圖3),總體上,整個扣蟹養殖周期(7—11月)的SGR大小順序為養選G1>對照組>野選G1。
由圖4可見,無論是雄體還是雌體,兩選育群體G1的早熟率均低于對照組,其中,野選G1的雄體及總體早熟率均顯著低于對照組(P<0.05); 與對照組相比,野選 G1的雄體、雌體及總體早熟率分別降低36.64%、35.35%和36.89%,養選G1的早熟率分別降低 14.31%、14.06%和 15.25%; 另外,三群體的雌體早熟率(11.95%—18.86%)均明顯高于雄體(5.01%—7.75%),表現出明顯的性別差異。
2.1.2 養殖規格、成活率、產量及餌料系數 從最終扣蟹規格看,三群體最終正常扣蟹與早熟蟹的平均體重均無顯著差異(表2,P>0.05); 但是,無論是雄體還是雌體,野選 G1和養選 G1的成活率均高于對照組(表3),相比之下,野選G1雄體、雌體和總體成活率分別比對照組高出 11.48%,12.47%和 12.09%,養選G1雄體、雌體和總體分別高出對照組8.94%、8.05%和8.61%。由于上述提及的對照組早熟率較高,故對照組早熟蟹的產量顯著高于野選 G1,而正常扣蟹的產量顯著低于兩個選育群體(P<0.05),三群體的正常扣蟹和早熟蟹總產量差異不顯著,均在3200kg/hm2左右; 三群體在扣蟹養殖階段的飼料系數順序為: 野選 G1(1.96)>對照組(1.83)>養選 G1(1.67),但三者無顯著差異(P>0.05)。

圖1 野選G1、養選G1在扣蟹階段的體重變化Fig.1 The changes of monthly body weight (WG) of the first generation (G1) of selective-breeding based on wild and cultured E. sinensis populations during the juvenile culture stage

圖2 野選G1、養選G1在扣蟹階段的月增重率Fig.2 Mean monthly weight gain (WGR) of the first generation(G1) of selective-breeding based on wild and cultured E. sinensis populations during the juvenile culture stage

圖3 野選G1、養選G1在扣蟹階段的月特定生長率Fig.3 Mean monthly specific growth rate (SGR) of the first generation (G1) of selective-breeding based on wild and culturedE. sinensis populations during the juvenile culture stage

圖4 野選G1、養選G1在扣蟹階段的早熟率Fig.4 The percentage of precocious of the first generation (G1)of selective-breeding based on wild and cultured E. sinensis populations during the juvenile culture stage
2.2 成蟹養殖階段的養殖性能
2.2.1 生長性能 三群體中華絨螯蟹扣蟹在成蟹養殖階段的體重變化見圖5,就雄體而言,9月份之前,養選G1生長較快,體重較高,自9月份起,野選G1體重迅速增加,在10月份,野選G1養成的雄蟹體重顯著高于對照組(P<0.05),而養選G1介于野選G1與對照組之間; 就雌蟹而言,8月份之前,兩選育群體體重均低于對照組,自9月份以后,兩選育群體的體重均高于對照組,但差異不顯著(P>0.05); 從最終養成成蟹的平均體重看,兩選育群體均高于對照組,但僅有野選G1雌蟹顯著高于對照組(P<0.05),野選G1的雄體、雌體、總體規格分別比對照組提高14.19%、10.56%和 12.38%,養選 G1分別比對照組提高10.59%、10.11%和10.40%; 隨著養殖時間的延長,三群體雌雄蟹的WGR均持續降低(圖6),其中在9—10月期間增重率極低,僅 4%左右,并且無論是雄體還是雌體,三群體的WGR僅在8—9月期間出現顯著差異,即在 8—9月野選 G1的 WGR顯著高于對照組(P<0.05); 三群體的SGR與WGR均表現出相似的變化規律(圖 7),但是無論是雄體還是雌體,整個養殖期間(3—10月)兩選育群體的SGR與對照組相比均無顯著差異(P>0.05)。

表2 野選G1、養選G1在扣蟹養殖階段產出的正常扣蟹和早熟蟹的規格比較Tab.2 Mean body weight of normal crab and precocious crab of the first generation (G1) of selective-breeding based on wild and cultured E. sinensis populations during the juvenile culture stage

表3 野選、養選G1在扣蟹養殖階段的成活率、產量和飼料系數的比較Tab.3 Survival,yield and feed conversion rate (FCR) of the first generation (G1) of selective-breeding based on wild and cultured E.sinensis populations during the juvenile culture stage

圖5 野選G1、養選G1在成蟹養殖階段的體重變化Fig.5 The changes of monthly body weight of the first generation (G1) of selective-breeding based on wild and cultured E. sinensis populations during the adult culture stage
2.2.2 性腺發育 三群體在 8月份起均陸續開始進行生殖蛻殼,圖8所示,在8月20日采樣時發現,野選 G1雌雄體的生殖蛻殼完成率均略低于對照組,而養選 G1雌雄體生殖蛻殼完成率均略高于對照組,到9月20日左右,養選G1及對照組的雌雄體生殖蛻殼完成率已經達到100%,野選G1中仍有部分的雌體未完成生殖蛻殼,生殖蛻殼完成率為 92%,整體上,野選 G1的生殖蛻殼稍滯后于對照組,養選 G1的生殖蛻殼稍提前于對照組,但差異不明顯; 生殖蛻殼完成后雄體和雌體的性腺發育速度加快,性腺指數逐漸提高,肝胰腺指數逐漸下降(表 4),無論是雌體還是雄體,野選 G1的性腺指數始終略低于對照組,養選 G1始終略高于對照組,這說明養選 G1性腺發育速度快于對照組,野選 G1慢于對照組; 對同一群體的雌雄體而言,雄體性腺發育時間明顯早于雌體,其在8月份性腺指數已在1%左右,而此時雌體僅為0.3%左右,此后雌體性腺指數增加速度顯著快于雄體。
2.2.3 成活率、產量及餌料系數 從表5可見,無論是雄體還是雌體,兩選育群體在成蟹養殖階段的成活率也均高于對照組5%左右(野選G1雄體、雌體提高及總體分別提高14.21%、12.77%和 12.27%; 養選G1分別提高9.64%、8.41%和9.30%),再加上兩選育群體的平均體重較高,以致兩選育群體的雌雄體產量均高于對照組,其中雄體及總體產量顯著高于對照組(P<0.05); 兩選育群體的飼料系數與對照組較為接近,均無顯著差異(P>0.05)。

圖6 野選G1、養選G1在成蟹養殖階段的月增重率Fig.6 Mean monthly body weight gain rate (WGR) of the first generation (G1) of selective-breeding based on wild and cultured E.sinensis populations during the adult culture stage

圖7 野選G1、養選G1在成蟹養殖階段的月特定生長率Fig.7 Mean monthly specific growth rate (SGR) of the first generation (G1) of selective-breeding based on wild and cultured E.sinensis populations during the adult culture stage

圖8 野選G1、養選G1在成蟹養殖階段8、9月份生殖蛻殼完成率比較Fig.8 The percentage of crabs that finished puberty molting during the August and September samplings of the first generation (G1) of selective-breeding based on wild and cultured E. sinensis populations during the adult culture stage

表4 野選G1、養選G1在成蟹階段肝胰腺指數及性腺指數的比較(%)Tab.4 Mean monthly variation in the hepatosomatic index (HSI) and gonadosomatic index (GSI) of the first generation (G1) of selective-breeding based on wild and cultured E. sinensis populations during the adult culture stage (%)

表5 野選G1、養選G1在成蟹養殖階段的成活率、產量和飼料系數的比較Tab.5 Survival,yield and feed conversion rate (FCR) of the first generation (G1) of selective-breeding based on wild and cultured E.sinensis populations during the adult culture stage
2.2.4 最終平均規格和規格分布 最終野選 G1、養選 G1和對照組養成雄體的平均體重分別為(186.318±19.46)g、(191.37±22.84)g和(165.40±17.68)g;雌體的平均體重分別為(122.45±15.73)g、(121.62±13.47)g和(112.90±13.96)g。圖9顯示,三群體最終養成成蟹的各規格百分比均呈正態分布,雄體主要集中于 175—199.9g和 150—174.9g兩個體重范圍,雌體主要集中于125—149.9g和100—124.9g。并且,無論是雄體還是雌體,兩選育群體的大規格蟹(雄體>175g,雌體>125g)比例均高于對照組; 由于不同圍隔間的差異較大,三群體間各種規格蟹的比例均無顯著差異(P>0.05)。
3 討論
先前的研究表明,野生捕撈和池塘養殖中華絨螯蟹扣蟹在相同養殖環境下進行成蟹養殖時,最終池塘養殖群體生殖蛻殼提前、性腺成熟時間較早,二齡早熟現象明顯,而野生群體具有明顯的二齡晚熟現象,但最終成蟹規格大(Heet al,2014),這給選育二齡早熟和晚熟品系培育提供了基礎材料,因此,本課題組以長江水系野生和池塘養殖群體為選育基礎群,以降低一齡早熟率、提前或延后二齡成熟期、提高養成規格為主要選育目標,采用繼代選育法組建保種性選育核心群,通過第一代的選育,進行定向對比養殖實驗,結果顯示,野選和養選 G1代在扣蟹養殖階段的一齡早熟率比對照組分別降低 36.89%和15.25%,成活率分別提高12.09%和8.61%,在成蟹養殖階段,兩選育群體的成活率分別比對照組提高12.27%和 9.30%,最終成蟹規格分別比對照組提高12.38%和10.40%,并且野選G1的生殖蛻殼和性成熟時間略有推遲,養選 G1的生殖蛻殼時間略有提前;中華絨螯蟹“長江1號”快速生長品系選育時,G1代的平均規格比未選育組提高6.64%(鄧燕飛等,2013),這暗示不同的選育基礎群體和不同選育策略均會影響其選育效果和遺傳進展。此外,不同物種選育到同一世代的選育效果可能也會有較大差異,如三疣梭子蟹(Portunus trituberculatus)快速生長品系“黃選1號”選育到 G1代規格提高了 2.91%,成活率提高 8.66%(李健等,2013); 凡納濱對蝦(Litopenaeus vannamei)的生長和抗 TSV(Taura syndrome virus)品系選育時G1代的生長速度提高 21.2%,成活率提高了18.4%(Argueet al,2002)。此外,中華絨螯蟹的一齡早熟率與一齡扣蟹養殖期間的成活率直接相關,在相同的養殖條件下,成活率高的群體或家系,則早熟率低; 反之亦然(何正侃等,1999),因此,本研究中兩選育群體一齡早熟率低是由遺傳決定還是由于兩選育群體在一齡扣蟹養殖期間的成活率高所致,這有待進一步研究。本研究中兩選育群體G1代最終養成規格大于未選育組(對照組),這可能是由于采用大規格親本蟹選育所達到的良好效果。總體上,本研究中野選和養選G1代在降低一齡早熟率、提高成活率和成蟹規格方面取得了較好的選育效果,但在二齡早熟和晚熟方面效果不夠顯著,這可能是由于本研究中的選育方法造成的。

圖9 野選、養選G1最終養成成蟹的規格分布Fig.9 Harvest size distribution of different body weight ranges from the first generation (G1) of selective-breeding based on wild and cultured E. sinensis populations adult E. sinensis
早熟和晚熟可能是生物體的一種病理狀態,也可能是農業生產中具有重要經濟價值的數量性狀,一些物種在進化或選育過程中形成了性早熟或晚熟的特性并能在群體中穩定遺傳,從而改變其成熟期,這對優化農產品的品種結構、延長市場供應期具有重要的作用(儲明星等,2009; 馮濤等,2009; 謝宗周等,2011; 冀曉昊等,2014)。性早熟和性晚熟在自然界許多魚類和甲殼動物上廣泛存在(Amanoet al,1995;Basavarajuet al,2002; Begtashiet al,2004),可能有著較為復雜的原因,連續家養多代的魚類養殖群體通常存在性早熟現象(Navarro-Martinet al,2011;Crespoet al,2013)。中華絨螯蟹的性早熟現象較為復雜,分為一齡扣蟹早熟和二齡成蟹早熟兩種不同的早熟現象,一齡扣蟹早熟通常出現于9月下旬后,這些幼蟹養殖 3—4個月便進行生殖蛻殼,從而啟動性腺快速發育,一齡早熟蟹體重在 15—35g,經濟價值和食用價值較低,這嚴重危害一齡扣蟹養殖階段的養殖性能和經濟效益(Wuet al,2011); 二齡早熟通常出現在成蟹養殖階段,一些成蟹在 7—8月便完成生殖蛻殼和啟動性腺發育,早于正常二齡蟹生殖蛻殼時間(8—9月),二齡早熟蟹體重通常小于二齡晚熟蟹,但其性腺成熟時間早于二齡正常成熟個體。由于部分二齡早熟蟹的出現,可以提前供應市場,有利于均衡上市,降低中華絨螯蟹上市時間過分集中的現象。因此,降低中華絨螯蟹一齡早熟率,提前或延遲二齡蟹成熟時間是今后中華絨螯蟹良種培育的重要目標,這有助于中華絨螯蟹養殖產業的健康可持續發展。
大量研究表明中華絨螯蟹性成熟時間由內因和外因共同決定,內因是其在長期進化過程中形成的自身遺傳特點(李晨虹等,2002; 張世勇等,2013)、種質混雜(李應森等,2001)、性別(Heet al,2014)等; 外因包括溫度(張列士等,2001)、光照(Liet al,2011a)、鹽度(魏薇等,2007)、營養(陳再忠等,2003; Wuet al,2011)和養殖密度(Liet al,2007)等。本研究中養選G1成熟時間提前、性腺發育提早,而野選 G1的二齡成熟時間延后,性腺發育推遲,與野生、養殖扣蟹原代(G0)在池塘內養殖所得結果類似(Heet al,2014),這說明,經過多年人工養殖的中華絨螯蟹養殖性狀已經發生了一些變化; 同時也暗示通過遺傳選育調控中華絨螯蟹性腺成熟及上市時間是可行的。經過多代家養的水產動物性腺成熟時間提前,從而生長時間變短和成體偏規格偏小,這可能與其表觀遺傳有關,特別是性腺發育相關功能基因的 DNA甲基化(Navarro-Martinet al,2011; Crespoet al,2013),有關中華絨螯蟹一齡和二齡性早熟的分子機制有待進一步深入研究。本研究中兩選育群體 G1一齡早熟率較低,二齡早熟和晚熟效果不明顯,其可能原因是:(1) 本研究中選用大規格野生和池塘養殖親本進行人工育苗,大規格親本及祖先在扣蟹養殖過程中可能早熟較低,從而造成其子代在扣蟹養殖階段的低早熟率(張德隆等,2001; Liet al,2011b); (2) 本研究中的二齡早熟和晚熟親本的選擇方法有待進一步提高,如: 親本選擇過程中,未能將二齡早熟和晚熟性狀與大規格親本兩類性狀有效結合進行良種選育。鑒于此,本課題組在G1代選育的基礎上,進一步優化良種選育的技術路線,系統研究中華絨螯蟹性成熟的第二性征變化,篩選快速評價指標來有效判別中華絨螯蟹生殖蛻殼和性腺發育階段,有利于良種選育過程為挑選個體較大的二齡早熟(7月下旬成熟)和晚熟(10月上旬成熟)個體進行定向選育,提高選育效果。
王 武,王成輝,馬旭洲,2013. 河蟹生態養殖. 北京: 中國農業出版社,59—84
王成輝,李思發,李晨虹等,2001. 池塘放養中華絨螯蟹長江種群與遼河種群性早熟出現差異的觀察與分析. 湖泊科學,13(1): 57—62
韋鳳英,廖玉英,羅蒙和等,2002. 銀香麻雞和霞煙雞早熟品系選育與應用研究. 中國家禽,24(18): 12—14
鄧燕飛,夏愛軍,潘建林等,2013. 中華絨螯蟹“長江1號”的選育. 水產養殖,34(4): 43—47
馮 濤,狄 冉,儲明星等,2009. 哺乳動物性晚熟相關基因的研究進展. 中國畜牧獸醫,36(2): 86—90
成永旭,王 武,吳嘉敏等,2000. 蝦蟹類幼體的脂類需求及脂類與發育的關系. 中國水產科學,7(4): 104—107
朱清順,柏如發,鄧燕飛,2012. 中華絨螯蟹‘長江一號’生長性能比較研究. 農學學報,2(1): 53—57
劉 磊,李 健,劉 萍等,2012. 三疣梭子蟹微衛星標記與生長相關性狀的相關性分析. 水產學報,36(7): 1034—1041
李 莉,許 飛,張國范,2011. 水產動物基因資源和分子育種的研究與應用. 中國農業科技導報,13(5): 102—110
李 健,劉 萍,高保全等,2013. 三疣梭子蟹新品種“黃選 1號”的選育. 漁業科學進展,34(5): 51—57
李小紅,王業建,謝運河等,2009. 極早熟優質高產春大豆湘春豆26的選育. 大豆科技,(5): 63,65
李應森,李思發,王江玲等,2001. 長江和遼河水系中華絨螯蟹湖泊放養生長性能的比較. 水產科學,20(5): 1—3
李曉暉,許志強,潘建林等,2010. 中華絨螯蟹人工選育群體的遺傳多樣性. 中國水產科學,17(2): 236—242
李晨虹,李思發,邢益于等,2002. 池養長江蟹、遼河蟹生長性能及其遺傳環境交互作用分析. 水生生物學報,26(4):335—341
李鴻鳴,孫效文,2002. 應用大規模家系選育技術促進遼寧海水養殖業的可持續發展. 沈陽農業大學學報(社會科學版),4(1): 7—10
吳 滟,付春鵬,蔣速飛等,2011. 中華絨螯蟹微衛星標記與生長性狀相關性的初步分析. 水生生物學報,35(2):197—202
何正侃,印 駿,朱雅珠,1999. 密度、營養與河蟹蟹種生長及性早熟之間的相關關系. 水產科技情報,26(2): 73—76
張世勇,傅洪拓,喬 慧等,2013. 中華絨螯蟹遺傳育種研究進展. 中國農學通報,9(20): 39—45
張列士,徐琴英,2001. 自然及養殖水體河蟹性成熟和性早熟的研究. 水產科技情報,28(3): 106—111
張德隆,杜曉燕,趙金利等,2001. 河蟹性早熟成因及控制方法的研究. 淡水漁業,31(4): 36—39
陳再忠,成永旭,王 武,2003. 早熟期間中華絨螯蟹肝胰腺指數、肝脂含量及脂肪酸組成的變化. 水產學報,27(1):57—61
郭正霞,辛秀娟,肖君華,2004. 性早熟相關基因的研究進展.國外醫學兒科學分冊,31(6): 300—302
盛志廉,陳瑤生,1999. 數量遺傳學. 北京: 科學出版社,191
傅洪拓,萬山青,付春鵬等,2010. 青蝦生長性狀相關的微衛星標記篩選,水生生物學報,34(5): 1043—1048
儲明星,馮 濤,狄 冉等,2009. 哺乳動物性早熟相關基因的研究進展. 遺傳,31(1): 13—28
謝宗周,鄧秀新,伊華林等,2011. 晚熟臍橙新品種——倫晚臍橙的選育. 果樹學報,28(4): 733—734
熊良偉,2014. 中華絨螯蟹遺傳連鎖圖譜構建及經濟相關性狀QTL定位. 上海: 上海海洋大學博士學位論文,64—67
冀曉昊,毛志泉,張艷敏等,2014. 晚熟蘋果新品種‘山農紅’.園藝學報,41(7): 1503—1504
魏 薇,吳嘉敏,魏 華,2007. 鹽度對中華絨螯蟹性早熟生理機制的影響. 中國水產科學,14(2): 275—280
Amano M,Hyodo S,Kitamura Set al,1995. Short photoperiod accelerates preoptic and ventral telencephalic salmon GnRH synthesis and precocious maturation in underyearling male masu salmon. General and Comparative Endocrinology,99(1): 22—27
Argue B J,Aree S M,Lotz J Met al,2002. Selective breeding of Pacific white shrimp (Litopenaeus vannamei) for grow th and resistance to Taura Syndrome Virus. Aquaculture,204(3—4): 447— 460
Basavaraju Y,Mair G C,Kumar H M Met al,2002. An evaluation of triploidy as a potential solution to the problem of precocious sexual maturation in common carp,Cyprinus carpio,in Karnataka,India. Aquaculture,204(3—4):407—418
Begtashi I,Rodríguez L,Moles Get al,2004. Long-term exposure to continuous light inhibits precocity in juvenile male European sea bass (Dicentrarchus labrax,L.). I.Morphological aspects. Aquaculture,241(1—4): 539—559
Crespo B,Gómez A,Mazón M Jet al,2013. Isolation and characterization of Ff1 and Gsdf family genes in European sea bass and identification of early gonadal markers of precocious puberty in males. General and Comparative Endocrinology,191: 155—167
He J,Wu X G,Li J Yet al,2014. Comparison of the culture performance and profitability of wild-caught and captive pond-reared Chinese mitten crab (Eriocheir sinensis)juveniles reared in grow-out ponds: Implications for seed selection and genetic selection programs. Aquaculture,434:48—56
Hetzel D J S,Crocos P J,Davis G Pet al,2000. Response to selection and heritability for growth in the Kuruma prawn,Penaeus japonicus. Aquaculture,181(3—4): 215— 223
Li X D,Dong S L,Lei Y Zet al,2007. The effect of stocking density Chinese mitten crabEriocheir sinensison rice and seed yields in rice-crab culture systems. Aquaculture,273(4):487—493
Li X W,Li Z J,Liu J Set al,2011a. Effects of light intensity on molting,growth,precocity,digestive enzyme activity,and chemical composition of juvenile Chinese mitten crabEriocheir sinensis.Aquaculture International,19(2): 301—311
Li X,Li Z,Liu Jet al,2011b. Advances in precocity research of the Chinese mitten crabEriocheir sinensis. Aquaculture Internation,19(2): 251—267
Navarro-Martin L,Vi?as J,Ribas Let al,2011. DNA methylation of the gonadal aromatase (cyp19a) promoter is involved in temperature-dependent sex ratio shifts in the European sea bass. PLoS Genetics,7(12): e1002447
Sui L Y,Wille M,Cheng Y Xet al,2011. Larviculture techniques of Chinese mitten crabEriocheir sinensis.Aquaculture,315(1—2): 16—19
Wu X G,Wang Z K,Cheng Y Xet al,2011. Effects of dietary phospholipids and highly unsaturated fatty acids on the precocity,survival,growth and hepatic lipid composition of juvenile Chinese mitten crab,Eriocheir sinensis(H.Milne-Edwards). Aquaculture Research,42(3): 457—468
