斜帶石斑魚(Epinephelus coioides)兩種神經肽NPB的發現及表達分析*
2015-03-08 05:27:34李偉萍宇肖李水生林浩然
海洋與湖沼 2015年4期
李偉萍 胡 哲 李 玉 王 翔 黃 文 王 宇肖 玲 李水生 張 勇 林浩然,①
(1. 海南大學海洋學院 海口 570228;2. 中山大學水生經濟動物研究所暨廣東省水生經濟動物良種繁育重點實驗室 廣州 510275)
魚類的攝食和生長受許多神經內分泌因子的調節,如食欲素(Orexin)、促腎上腺皮質素釋放因子(corticotropin releasing factor,CRF)、神經肽 Y (Neuropeptide Y,NPY)、神經肽 W (Neuropeptide W,NPW)和神經肽 B(Neuropeptide B,NPB)等(傅小鎖等,1993; 肖東等,2000; 譚雪梅等,2004; 李昀等,2009; 雷治海等,2010)。在2002—2003年間,NPB作為G蛋白偶聯受體GPR7 (Neuropeptide B/W receptor-1,NPBWR1)和GPR8 (Neuropeptide B/W receptor-2,NPBWR2)的內源性配體從哺乳動物的下丘腦提取物中被分離鑒定出來(Fujiiet al,2002; Shimomuraet al,2002;Brézillonet al,2003; Tanakaet al,2003)。隨著研究的不斷深入,NPB被認為是與調節攝食,能量平衡,睡眠,神經內分泌功能以及調節炎癥性疼痛等功能相關(Ishiiet al,2003; Aikawaet al,2008; Hirashimaet al,2011; Skrzypskiet al,2012; Watanabeet al,2012)。
NPB廣泛分布在整個腦組織的不同部位如海馬組織、下丘腦室旁核、動眼神經副核和中腦導水管周圍灰質等(Tanakaet al,2003; Jacksonet al,2006),特別是在下丘腦中,均能檢測到了NPB與GPR7 mRNA的高度表達(Schulzet al,2007),此外,在哺乳動物中的一些外周組織,也檢測到 NPB mRNA的表達(Brézillonet al,2003)。
在硬骨魚類的研究中,NPB已從斑馬魚(Tanakaet al,2003)、鮭魚(Leonget al,2010)和羅非魚(Yanget al,2014)等魚類中克隆得到,并發現NPB與攝食調控活動相關。由于魚類作為脊椎動物中種類最多的一個群體,其進化的原始性、物種的多樣性以及生活環境的復雜性,其攝食調控機制也復雜而多樣性。斜帶石斑魚(Epinephelus coioides)是我國南方重要的經濟魚類,研究其生長與攝食調控機制具有重要的應用價值。
本文利用生物信息學分析和基因克隆技術在斜帶石斑魚中克隆得到了兩種 NPB基因 cDNA序列,分析了 NPB的信號肽及成熟肽結構及其在不同組織器官中的分布表達模式,并進一步研究了其在攝食調控下的表達模式,證明了這兩種 NPB均參與了斜帶石斑魚攝食調控。
1 材料與方法
1.1 樣品采集
1.1.1 克隆和組織分布的樣品采集 本實驗所用斜帶石斑魚來自廣東省大亞灣水產試驗中心,均為健康的1—2齡魚,體重為720—770g,并以25—30°C循環過濾海水飼養。取樣前,先用冰塊冷凍麻醉1—2 min,然后取出其不同分區的腦組織和外周組織,并放入液氮中進行速凍處理,而后轉存放于–80°C超低溫冰箱備用。取樣時所用的所有解剖器械(如剪刀、鑷子、錫箔紙等)均經過180°C 4h以上的烘烤滅菌,存放樣品的1.5 mL離心管為進口離心管(不含有RNA酶)。所有的動物實驗均按照中山大學動物研究倫理委員會對進行動物實驗時所做的規章制度進行操作。
1.1.2 饑餓實驗的樣品采集 實驗所用斜帶石斑魚來自廣東省大亞灣水產試驗中心,體重 32—60 g,體長 12—15 cm的仔魚。將仔魚(n=150)馴養于室內海水池中,并把短期饑餓的魚分成 6組,馴養(每天9:00定點投喂)兩周后開始進行短期(7d)饑餓實驗:實驗第 3天取饑餓組和對照組(投喂組)樣品,并將其中3組恢復投喂; 實驗第7天取饑餓組、對照組(投喂組)和饑餓復投組樣品。取下的下丘腦組織于凍存管中,液氮速凍保存帶回實驗室,并將其轉存到–80°C超低溫冰箱備用。實驗所用的解剖器具和玻璃器皿等均經過 180°C烘烤5h用以滅活RNA酶。
1.2 方法和處理
1.2.1 引物設計 根據已有的斜帶石斑魚轉錄組數據(未公開)和 GenBank上已上傳的 NPB基因序列的比對分析,找出了兩個與 NPB相似的基因序列,并暫命名為NPB1和NPB2。利用引物設計軟件primer premier 5.0設計斜帶石斑魚NPB克隆和RT-PCR的特異性引物。具體引物序列如表1所示。

表1 PCR引物序列Tab.1 Sequences of the Primers used for PCR
1.2.2 總 RNA提取及 cDNA模版的制備 參照Trizol?reagent(由美國Invitrogen公司提供)說明書及操作步驟,將總 RNA提取出來,并通過反轉錄技術制備 cDNA模版以備基因克隆(polymerase chain reaction,PCR)和實時定量PCR (real time-polymerase chain reaction,RT-PCR)使用。
1.2.3 NPB基因的克隆與分析 以腦組織反轉錄產物為模板,利用NPB1和NPB2的簡并引物(NPB-F和 NPB-R)進行聚合酶鏈式反應(PCR)來克隆和驗證NPB1和NPB2基因。通過瓊脂糖凝膠電泳后將PCR產物的目的條帶進行切膠回收,用膠回收試劑盒E.Z.N.A?Plasmid Extraction Kit (Omega Bio-Tek公司,USA)進行分離純化,并將膠回收產物進行連接和轉化以及菌株培養鑒定,最后送至廣州英駿公司進行測序。根據測序結果利用 DNAssist2.2和BioEdit軟件將 NPB翻譯成相應的氨基酸序列,通過使用Clustal W1.83 和MEGA 4.0軟件將NPB的蛋白序列進行多重比對和進化樹分析,并在在線預測網站SignalP 4.1 server (http://www.cbs.dtu.dk/services/SignalP/)和SMART (http://smart.embl-heidelberg.de/)對其NPB進行信號肽的預測分析等。
1.2.4 實時定量 PCR及數據分析 根據以上的實驗方法,提取總RNA制備定量cDNA模版以及提取標準質粒 DNA制備標準曲線,以備 RT-PCR使用。選取β-actin作為內參基因和選用 SYBR?Green Realtime PCR Master Mix plus kit (TOYOBO公司,Japan)試劑盒進行 RT-PCR試驗。使用儀器為 ABI 7900HT sequence detection system。具體反應程序為:首先95°C預變性2min; 然后 95°C變性15s,55°C退火15s,72°C延伸30s,共40個循環; 最后在84°C收集熒光。對溶解曲線分析,以確定PCR產物質量,用Excel對實驗數據進行初步處理,再利用 Graphpad Prism5.0軟件進一步處理和分析數據。
2 結果與分析
2.1 斜帶石斑魚NPB1和NPB2基因序列的鑒定與分析
在本研究中,作者在對斜帶石斑魚 NPB基因克隆分析的基礎上,發現了兩個NPB基因(分別命名為NPB1和NPB2),其開放閱讀框(ORF)分別為393bp和387bp,各自編碼的蛋白質序列為 130個氨基酸(aa)和128aa,如圖1a所示。在NPB1和NPB2的氨基酸序列的N-末端均有一個包含了25aa的預測信號肽,而緊隨其預測信號肽之后則出現了各自的包含 29aa成熟肽,以及兩個由氨基酸殘基(RR)構成的蛋白質水解切割位點(Arg49-Arg50和Arg55-Arg56),并且都具有一個相對較長的C-末端,如圖1b所示。
如表 2所示,根據不同物種的 NPB的氨基酸序列比對表可知,斜帶石斑魚NPB1的氨基酸序列與尼羅羅非魚和斑馬宮麗魚的同源性最高高達 92%; 其次是劍魚、青 鳉和斑馬魚等,同源性高達 60%以上;而與哺乳類的同源性則比較低,如人類的52%,小家鼠的48%和大鼠45%。而斜帶石斑魚NPB2與鮭魚的同源性最高,為61%; 與尼羅羅非魚、青 鳉和斑馬魚等都在 50%以上; 與哺乳動物的相似性則在 45%左右,其中與人類的相似性為48%,較NPB1與人類的同源性(52%)略低。

表2 斜帶石斑魚NPB1和NPB2蛋白質前體氨基酸序列與其它脊椎動物的同源性百分比Tab.2 Pairwise comparison in homology for E. coioides NPB1 and NPB2 protein sequence with other species NPB protein sequences
如圖2所示,根據斜帶石斑魚與某些典型的哺乳動物和魚類的 NPB的氨基酸序列所構建的進化樹系統分析表明,NPB前體蛋白斜帶石斑魚的 NPB1和NPB2基因共同聚集在硬骨魚類的一個大分支上而與哺乳類分開。其中NPB1又與尼羅羅非魚和斑馬宮麗魚關系較近, 與劍尾魚和青 鳉關系次之,而與哺乳類關系較遠,與鳥類關系最遠; NPB2則與鮭魚關系相近,與斑馬魚關系次之,同樣與哺乳類和鳥類關系較遠。
2.2 NPB在斜帶石斑魚中的組織分布表達模式
如圖 3所示,通過逆轉錄 PCR和實時熒光定量PCR技術在斜帶石斑魚的不同器官組織中對NPB1和NPB2的mRNA的表達水平進行了空間表達模式的檢測,所有的數據都表明了,NPB1和NPB2基因在腦組織中廣泛表達,特別是在下丘腦中的表達極其豐富。其中而NPB1其它腦區如端腦、中腦、延腦和垂體的表達量較低,特別是嗅球和小腦的表達量在腦區中是最低的; 而NPB2則在嗅球和中腦的表達量是最低的。NPB1和NPB2在其它外周組織器官中也有少量的表達,如NPB1在性腺、鰓和肌肉中的表達量相對較高一點,而NPB2則在性腺和心臟中的表達量相對較高,其它組織如鰓、脾臟、肌肉、胃、腸和肝臟中的表達量都相對較低一些。由此可見,NPB2與NPB1在外周組織中的表達模式是有些微差異的,即 NPB2在外周組織中的表達更為廣泛; 但是兩者都在下丘腦中擁有更為豐富的分布表達。
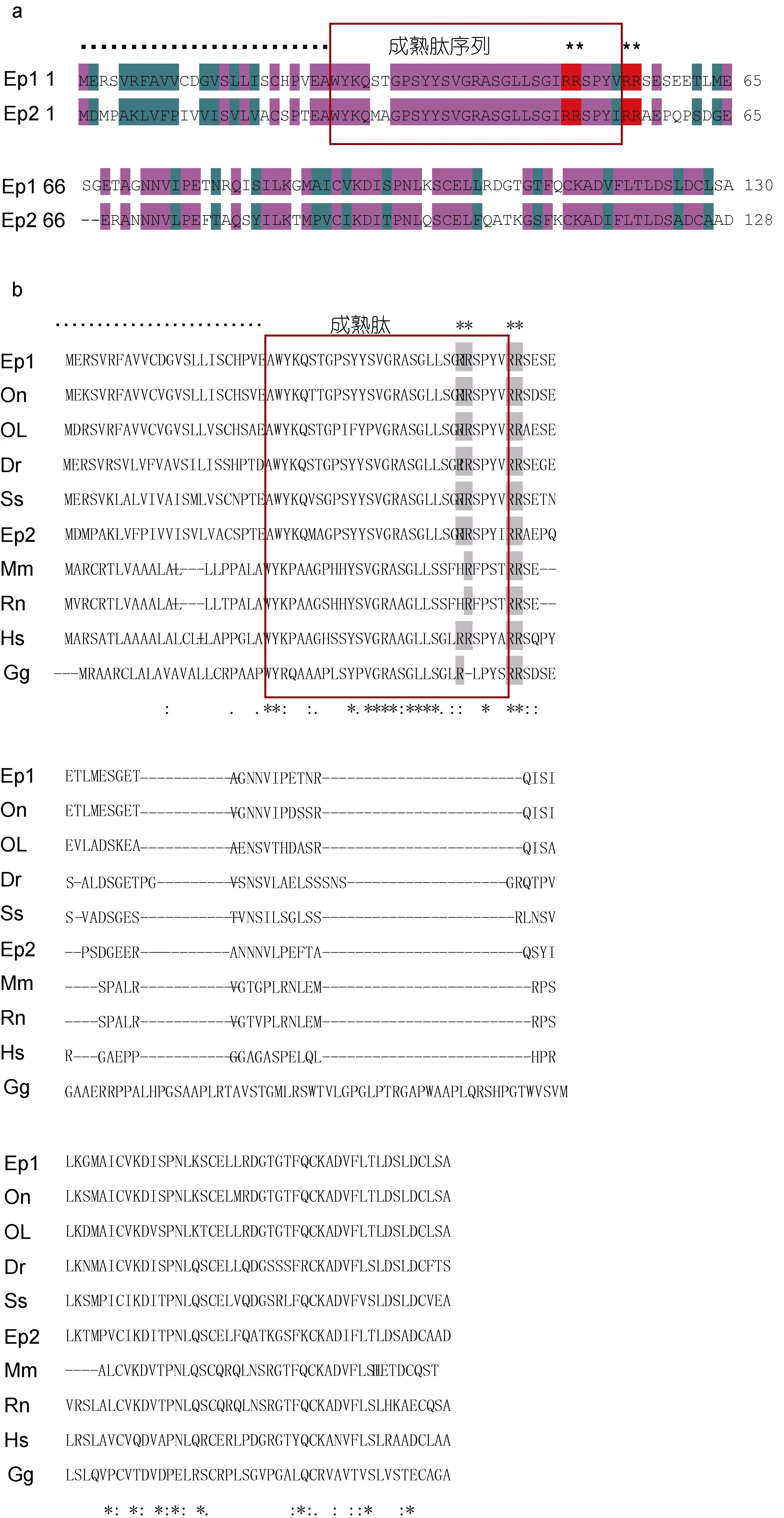
圖1 a. 斜帶石斑魚NPB1和NPB2蛋白質前體氨基酸序列比對,b. 典型脊椎動物NPB蛋白質前體氨基酸序列比對Fig.1 Alignment of the deduced amino acid sequence of NPB (neuropeptide B) protein sequencesa: NPB1 and NPB2 of E. coioides,b. Alignment of E. coioides NPBs protein sequences with other species’小黑點“?”表示預測的信號肽序列; 方框表示成熟肽序列; 上星號“*”表示由兩個精氨酸RR氨基酸殘基組成的蛋白質水解切割位點。下星號“*”表示一致的氨基酸; 冒號“:”表示高度保守度的氨基酸; 小圓點“.”表示低保守度的氨基酸。各物種的NPB縮寫詞如下: Ep1和Ep2,斜帶石斑魚(Epinephelus coioides) NPB1和NPB2; On,尼羅羅非魚(Oreochromis niloticus,GenBank accession No. XP_003455810.1);Ol,青鳉(Oryzias latipes,No. XP_004066211.1); Dr,斑馬魚(Danio rerio,No. NP_001120841.1); Ss,鮭魚(Salmo salar,No. ACI66524.1);Mm,小家鼠(Mus musculus,No. XP_006532795.1); Rn,大鼠(Rattus norvegicus,No. NP_695205.1); Hs,人類(Homo sapiens,No.NP_683694.1)Gg,雞(Gallus gallus,No. XP_001231800.2)

圖2 斜帶石斑魚NPB1和NPB2與其它物種的進化樹分析Fig.2 Phylogenetic tree illustrating the relationship between E. coioides NPB1 and NPB2 with those of other species NPB斜帶石斑魚(Epinephelus coioides) NPB1和NPB2; 尼羅羅非魚(Oreochromis niloticus,GenBank accession No. XP_003455810.1); 斑馬宮麗魚(Maylandia zebra,No. XP_004539074.1); 劍尾魚(Xiphophorus maculatus,No. XP_005805134.1); 青鳉(Oryzias latipes,No.XP_004066211.1); 斑點雀鱔(Lepisosteus oculatus,No. XP_006635208.1); 矛尾魚(Latimeria chalumnae,No. XP_005989170.1); 斑馬魚(Danio rerio,No. NP_001120841.1); 鮭魚(Salmo salar,No. ACI66524.1); 非洲爪蟾蜍(Xenopus (Silurana) tropicalis,No. XP_002937351.1);雪貂(Mustela putorius furo,No. XP_004749308.1); 雞(Gallus gallus,No. XP_001231800.2); 小家鼠(Mus musculus,No. XP_006532795.1);大鼠(Rattus norvegicus,No. NP_695205.1); 人類(Homo sapiens,No. NP_683694.1)
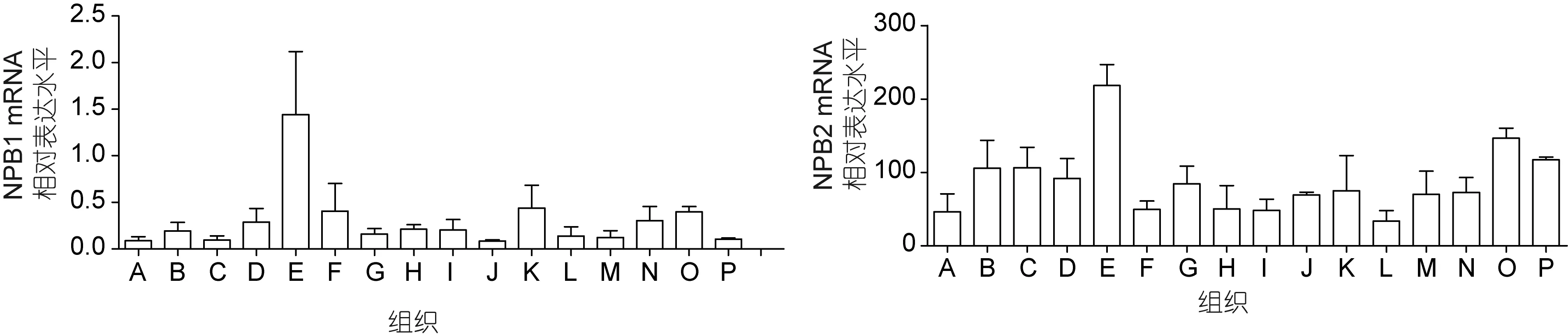
圖3 斜帶石斑魚NPB1和NPB2在不同組織中的表達模式Fig.3 Tissue distribution of NPB1 and NPB2 of E. coioides所有數值用平均值±標準差(mean±SEM)來表示。各個字母所代表的組織器官如下: A. 嗅球; B. 端腦; C. 小腦; D. 垂體; E. 下丘腦; F.中腦; G. 延腦; H. 肝臟; I. 胃; J. 腸道; K. 肌肉; L. 腎臟; M. 脾臟; N. 鰓; O. 性腺; P. 心臟
2.3 饑餓試驗中NPB在下丘腦表達模式
如圖4所示,在饑餓試驗的研究中發現,斜帶石斑魚在禁食3d和7d后,其NPB1和NPB2的mRNA相對表達量與投喂組的相比較,都顯著發生了下調;復投喂后,NPB1和NPB2的mRNA表達恢復,與對照組水平相當。
3 討論
基于 GenBank數據庫和實驗室研究組已有的石斑魚轉錄組數據的比對分析結果,作者在斜帶石斑魚中克隆得到了兩種編碼神經肽 NPB的亞型基因NPB1和 NPB2,其在 N-末端的信號肽和成熟肽結構序列,與尼羅羅非魚、斑馬魚、小鼠和人類等均在N-末端擁有的信號肽和成熟肽結構相似。除了哺乳動物,NPB在斑馬魚、非洲爪蟾和羅非魚中也已被克隆出來(Tanakaet al,2003; Yanget al,2014),NPB是由不同的肽前體衍生的,即前體NPB (Hirashimaet al,2011),成熟的神經肽NPB則由29個氨基酸組成,并且它在 N-末端的色氨酸殘基翻譯后被溴化修飾(Fujiiet al,2002),這些數據表明,斜帶石斑魚NPB的蛋白質的一級結構與其它脊椎動物相一致。同源性分析表明,斜帶石斑魚NPB1和NPB2均與其它魚類NPB的同源性相對較高,與哺乳類和雞的同源性則相對比較低; 其中與哺乳類和雞相比較,NPB1的同源性則比NPB2略高。而根據斜帶石斑魚與某些典型的哺乳動物和魚類的 NPB的氨基酸序列所構建的進化樹系統分析數據也證實了這一結果,NPB1和NPB2基因共同聚集在硬骨魚類的一個大分支上而與哺乳類和鳥類分開。由此可見在,斜帶石斑魚的NPB1的氨基酸序列在結構上是比較保守的,而NPB2的保守性則比較低。此外,斜帶石斑魚NPB1和NPB2的氨基酸序列的相似性較高,達59%,并且都有兩個潛在的由精氨酸殘基(RR)組成的蛋白質水解切割位點(Arg49-Arg50和Arg55-Arg56),與人類和尼羅羅非魚相似(Fujiiet al,2002; Yanget al,2014),這意味著斜帶石斑魚這兩種亞型 NPB的成熟肽可能各自有兩種肽蛋白質異構體NPB23和NPB29參與其生理調控功能。

圖4 食物剝奪試驗中斜帶石斑魚NPB1和NPB2在下丘腦的表達模式Fig.4 Expression pattern of the E. coioides NPB1 and NPB2 in food deprivation fed: 正常投喂組; unfed: 饑餓處理組; refed: 饑餓處理后復投喂組。NPB mRNA相對表達水平用平均值±標準差(mean±SEM)表示(n=10);*表示與正常投喂組差異顯著(P<0.05)
組織分布的研究表明,斜帶石斑魚 NPB1和NPB2基因在腦組織中廣泛表達,特別是在下丘腦中的表達極其豐富,這一結果與其它脊椎動物中的NPB表達分布情況相一致(Fujiiet al,2002; Brézillonet al,2003; Tanakaet al,2003; Yanget al,2014)。在哺乳動物中,NPB及其受體GPR7在腦組織區域廣泛表達,特別是在下丘腦中表達豐富(Hocholet al,2006;Schulzet al,2007; Hondoet al,2008; Sakurai,2013),如 NPB廣泛分布在整個腦組織的不同部位如海馬組織、下丘腦室旁核、動眼神經副核和中腦導水管周圍灰質等(Tanakaet al,2003; Jacksonet al,2006)。在大鼠中,NPB-免疫反應的陽性體細胞被發現于下丘腦內的許多區域,并且在下丘腦的這些區域也檢測到GPR7 mRNA的高濃度表達,這些區域包括下丘腦腹內側核、背內側下丘腦核、弓狀核、視前核和從腹側到未定帶(Schulzet al,2007)。而在硬骨魚類中,也分別在斑馬魚(Tanakaet al,2003)、鮭魚(Leonget al,2010)和尼羅羅非魚(Yanget al,2014)等的下丘腦中檢測到了 NPB的豐富表達。此外,在哺乳動物中的一些外周組織,如在人類的腎臟、子宮、卵巢、睪丸和胎盤中,在鼠科動物的胃、脊髓、睪丸、肝和腎等組織中也檢測到 NPB mRNA 的表達(Brézillonet al,2003)。與此相似,在斜帶石斑魚的外周組如性腺、心臟和鰓等織器官中也檢測到了 NPB mRNA的表達,特別是 NPB2在外周組織中的表達更為廣泛。由于NPB1和NPB2在斜帶石斑魚中樞神經系統的廣泛分布狀和在各種外周組織的表達模式,暗示著這兩種神經肽在斜帶石斑魚中可能與許多生理活動的調節有關。
由于下丘腦是許多復雜的生理活動的調節中樞,而就NPB和GPR7在下丘腦的分布情況推測其與攝食調控和能量消耗有關(Hondoet al,2008; Civelliet al,2008),而前人的研究也表明NPB/GPR7與攝食行為調節有關(Tanakaet al,2003; Samsonet al,2004;Johanssonet al,2008)。在本研究中,斜帶石斑魚在禁食3d和7d后,其NPB1和NPB2的mRNA相對表達量與正常投喂組相比都具有顯著的下調表達變化;而復投喂后,NPB1和NPB2恢復上調表達并與正常投喂組無明顯差異。這一結果表明NPB1和NPB2均參與了斜帶石斑魚的攝食調控。尼羅羅非魚在禁食7d和14d后,NPB在下丘腦的表達沒有明顯的變化,而在復投喂之后出現了明顯的上調表達(Yanget al,2014)。這些結果表明,在硬骨魚類中,NPB也參與了攝食調節。
李 昀,肖云虹,張 磊,2009. 促腎上腺皮質激素釋放因子及其受體的研究. 山東醫藥,49(48): 110—111
肖 東,林浩然,2000. 魚類攝食和生長的神經內分泌調控途徑研究進展. 中國水產科學 7(3): 102—106
傅小鎖,張景艷,1993. 神經肽 Y 及其作用. 生理科學進展,24(2): 152—155
雷治海,蘇 娟,賈曉慶等,2010. 神經肽W和神經肽B的研究進展. 畜牧與獸醫,42(8): 94—99
譚雪梅,陳代文,羊云飛,2004. 增食欲素及其生理功能. 中國飼料,(14): 2—7
Aikawa S,Ishii M,Yanagisawa Met al,2008. Effect of neuropeptide B on feeding behavior is influenced by endogenous corticotropin-releasing factor activities.Regulatory Peptides,151(1—3): 147—152
Brézillon S,Lannoy V,Franssen J-Det al,2003. Identification of natural ligands for the orphan G protein-coupled receptors GPR7 and GPR8. Journal of Biology Chemistry,278(2):776—783
Civelli O,Zhou Q-Y,2008. Orphan G protein-coupled receptors and novel neuropeptides. Berlin Heidelberg: Springer,1—256
Fujii R,Yoshida H,Fukusumi Set al,2002. Identification of a neuropeptide modified with bromine as an endogenous ligand for GPR7. Journal of Biological Chemistry,277(37):34010—34016
Hirashima N,Tsunematsu T,Ichiki Ket al,2011. Neuropeptide B induces slow wave sleep in mice. Department of Molecular Pharmacology,University of Tsukuba,34(1): 31—37
Hochol A,Belloni A S,Rucinski Met al,2006. Expression of neuropeptides B and W and their receptors in endocrine glands of the rat. International Journal of Molecular Medicine,18(6): 1101—1106
Hondo M,Ishii M,Sakurai T,2008. The Npb/NPW neuropeptide system and its role in regulating energy homeostasis,pain,and emotion. Results and Problems in Cell Differentiation,46: 239—256
Ishii M,Fei H,Friedman J M,2003. Targeted disruption of GPR7,the endogenous receptor for neuropeptides B and W,leads to metabolic defects and adult-onset obesity. Proceedings of the National Academy of Science,100(18): 10540—10545
Jackson V R,Lin S H,Wang Z Wet al,2006. A study of the rat neuropeptide B/neuropeptide W system using in situ techniques. Journal of Comparative Neurology,497(3):367—383
Johansson A,Fredriksson R,Winnergren Set al,2008. The relative impact of chronic food restriction and acute food deprivation on plasma hormone levels and hypothalamic neuropeptide expression. Peptides,29(9): 1588—1595
Leong J S,Jantzen S G,Von Schalburg K Ret al,2010.Salmo salarandesox luciusfull-length cDNA sequences reveal changes in evolutionary pressures on a post-tetraploidization genome. Victoria,British Columbia: Biology,Centre for Biomedical Research,University of Victoria,279
Sakurai T,2013. NPBWR1 and NPBWR2: Implications in energy homeostasis,pain,and emotion. Front Endocrinol(Lausanne),4: 23
Samson W K,Baker J R,Samson C Ket al,2004. Central neuropeptide B administration activates stress hormone secretion and stimulates feeding in male rats. Journal of Neuroendocrinology,16(10): 842—849
Schulz S,Stumm R,H?llt V,2007. Immunofluorescent identification of neuropeptide B-containing nerve fibers and terminals in the rat hypothalamus. Neuroscience Letters,411(1): 67—71
Shimomura Y,Harada M,Goto Met al,2002. Identification of neuropeptide W as the endogenous ligand for orphan G-protein-coupled receptors GPR7 and GPR8. The Journal of Biological Chemietry,277(39): 35826—35832
Skrzypski M,Pruszyńska-Oszmalek E,Ruciński Met al,2012.Neuropeptide B and W regulate leptin and resistin secretion,and stimulate lipolysis in isolated rat adipocytes. Regulatory Peptides,176(1—3): 51—56
Tanaka H,Yoshida T,Miyamoto Net al,2003. Characterization of a family of endogenous neuropeptide ligands for the G protein-coupled receptors GPR7 and GPR8. Proceedings of the National Academy Sciences,100(10),6251—6256
Watanabe N,Wada M,Irukayama-Tomobe Yet al,2012. A single nucleotide polymorphism of the neuropeptide B/W receptor-1 gene influences the evaluation of facial expressions. PLoS One,7(4): e35390
Yang L,Sun C Y,Li W S,2014. Neuropeptide B in nile tilapiaOreochromis niloticus: Molecular cloning and its effects on the regulation of food intake and mRNA expression of growth hormone and prolactin. General and Comparative Endocrinology,200(1): 27—34
