四川羚牛的家域與忠誠度
2015-03-11 06:47:31官天培葛寶明諶利民游章強唐中海宋延齡
生態學報 2015年6期
官天培,葛寶明,諶利民,游章強,唐中海, 劉 昊,宋延齡
1 綿陽師范學院生態安全與保護四川省重點實驗室, 綿陽 621000 2 鹽城師范學院江蘇省灘涂生物資源與環境保護重點建設實驗室, 鹽城 224002 3 四川唐家河國家級自然保護區,青川 628109 4 中國科學院動物研究所動物生態學與保護生物學重實驗室, 北京 100101
四川羚牛的家域與忠誠度
官天培1,葛寶明2,諶利民3,游章強1,唐中海1, 劉 昊1,宋延齡4,*
1 綿陽師范學院生態安全與保護四川省重點實驗室, 綿陽 621000 2 鹽城師范學院江蘇省灘涂生物資源與環境保護重點建設實驗室, 鹽城 224002 3 四川唐家河國家級自然保護區,青川 628109 4 中國科學院動物研究所動物生態學與保護生物學重實驗室, 北京 100101
野生動物傾向回到或留在一個特定范圍或者與原有區域完全重疊的行為被稱為棲息地忠誠。利用GPS無線電頸圈對5只四川羚牛的家域及家域的季節和年度忠誠度進行了研究和分析(2006—2009年)。結果顯示:四川羚牛年均家域面積為(MCP/FKE)(15.01±2.92)km2/(9.02±1.85) km2,但個體間及年際波動較大;季節間家域面積差異顯著, 個體家域的季節變化體現出較一致的變化模式,最大季節家域主要集中于春季和夏季。年際間季節家域忠誠度最高的是秋季和夏季,冬季家域年際忠誠度最低,春季家域忠誠度也相對較低。單因素方差分析顯示季節間質心距離總體差異不顯著,與家域重疊算法獲得的忠誠度結論基本一致。
最小凸多邊形;固定核域法;有蹄類;四川羚牛;家域
家域(home range)是動物生態學中的常見概念及基本生態習性信息,是指動物完成取食、交配和育幼等行為所利用的區域[1]。一般而言,家域的最小面積取決于個體生存和繁殖的最低成本,但它必須提供足夠的生存資源。家域面積除受限于食物、水源和隱蔽條件等環境因子,其穩定性也同時受到種群密度和捕食風險等生態因子的影響[2-5]。因此,研究野生動物家域是探討資源的空間分布狀況與動物生活史過程和生存策略的基礎。
許多動物會進行季節性遷移且在遷移完成后傾向于回到相對固定的棲息地內或附近,將這種行為傾向稱為棲息地忠誠[6]。棲息地忠誠行為廣泛存在于昆蟲[6]、魚類[7]、兩棲類[8-9]、鳥類[10-12]和哺乳動物[13-14]等眾多不同動物類群。然而,忠誠度的研究卻存在諸多挑戰和不確定性:其一,文獻中出現的忠誠度定義因作者和研究物種而異,有的是指繁殖地[14],有的是指領域或家域[15-16],有的指的是具體的巢址[10];其二,研究棲息地忠誠度需要對個體進行標識,并獲取動物在數輪遷移或遷徙過程中活動的位置信息,實踐中很難獲得這樣的數據;其三,界定個體在特定時間尺度(如季節)的活動邊界往往存在主觀因素。因此,將一個相對完整的周期作為忠誠度評價的時間尺度可能是較適合的,例如年際家域的忠誠度。在本研究中,棲息地忠誠度是指家域尺度的忠誠度。
羚牛(Budorcastaxicolor)是分布在青藏高原東段的珍稀有蹄類,具有明顯的季節性垂直遷移習性[17-18]。由于棲息地的喪失和偷盜獵泛濫,羚牛被IUCN列為易危(UV),被我國政府作為Ⅰ級重點保護動物[19]。宋延齡等對秦嶺羚牛季節性家域進行了估算并對家域變化原因進行初步推斷,認為食物資源的多度與分布可能導致家域季節變化的主要因素[20]。四川羚牛,即羚牛四川亞種,被認為是目前現存羚牛四個亞種中種群最大,分布最廣的[19]。然而至今未見關于羚牛家域忠誠度的相關研究和四川羚牛家域的報道。為探索以羚牛為代表的大型山地有蹄類的生存策略與資源時空分布關系提供基礎,本文嘗試回答以下問題:(1)四川羚牛家域的季節及年際變化特征;(2)四川羚牛對家域是否忠誠,其強度如何?
1 研究地點和研究方法
1.1 研究地點
本研究開展于四川省唐家河國家級自然保護區(以下簡稱唐家河,東經104°36′—104°56′,北緯32°32′—32°41′)。唐家河地處岷山山系摩天嶺南麓, 面積400km2,最低海拔1100m,最高海拔3864m,屬于低山丘陵向高山峽谷過渡的地帶,復雜崎嶇是唐家河地形的重要特征。唐家河成立于1978年,是以大熊貓(Ailuropodamelanoleuca)、川金絲猴(Rhinopithecusroxellana)、羚牛等珍稀野生動物及其棲息地為主要保護對象的自然保護區。保護區內的所有居民于1990年前已全部遷至區外,人為干擾微弱。植被類型隨海拔梯度變化明顯,從低海拔至高海拔依次是河谷灌叢及灌木林,落葉闊葉林,針闊混交林,針葉林以及高山草甸。區內主要分布有3種箭竹,分別是缺苞箭竹(Fargesiadenudata)、糙花箭竹(Fargesiascabrida)和青川箭竹(Fargesiarufa),主要集中分布于1800—2600m的林下。區內屬于溫帶氣候,根據保護區內氣象記錄(海拔1700m),最低溫發生在1月份(-11℃), 最高溫發生在7月(30℃)。每年的6月至9月為雨季,年降水量達1100mm。
1.2 羚牛的捕捉及數據下載方法
從2006年10月至2009年4月,在獲得國家林業局批準后,由專業獸醫及麻醉師為9只不同群體的成年羚牛個體佩戴了帶有GPS模塊的無線電頸圈(Lotek GPS 4400 M,Canada,重量不及羚牛體重的1%)。整個捕捉過程符合國家的相關法律規定并由保護區全程配合。每個頸圈都在每天固定的時間開啟4次(24:00、7:00、12:00、19:00)、每次15分鐘的自動定位程序。從佩戴頸圈之日算起,每隔3個月,用手持無線下載終端激活頸圈,將所有詳細定位信息,包括時間、日期、經緯度、海拔、衛星數量及位置精度衰減因子(Position Dilution of Precision,PDOP)等傳輸到便攜式計算機中。
1.3 家域及忠誠度的計算方法
對季節家域忠誠度的分析,首先計算各季節的家域面積。本研究中,按照當地的氣候條件、同時也考慮數據量的均衡性,對季節進行劃分。冬季為每年的12月至次年2月,春季3月至5月,夏季6月至8月,秋季9月至11月。按照以下標準選擇用于計算家域面積的數據:(1)GPS定位數據的精度均需達到3D水平,同時要滿足PDOP(位置精度衰減因子)小于10的條件[17];(2)一天內只使用1個GPS定位點計算每只羚牛的家域面積。
進行家域估計時,一方面使用最小凸多邊形法(Minimum Convex Polygon, MCP)計算家域,可以與以往的研究進行比較;另一方面使用固定核域法(Fixed Kernel Estimation, FKE)彌補MCP法的不足。家域計算是在ArcGIS 9.2中進行,使用了Hawth tools for ArcGIS9.2工具包,獲得MCP和FKE家域面積。家域忠誠度的衡量依據是家域的重疊比例(重疊面積占前一年際/季節的家域比例),先通過ArcGIS中的overlap工具形成的重疊區域的多邊形,再獲得該多邊形的面積。此外,還嘗試用家域多邊形質心間的距離來評價羚牛家域忠誠度。在進行數據分析時,首先檢驗數據是否符合正態分布,然后依據檢驗結果分別選擇單因素方差分析(One way ANOVA)或K個獨立樣本檢驗進行相關的分析和檢驗(如檢驗季節間家域是否存在差異或檢驗不同季節的家域忠誠度是否存在差異),數據以平均值 ± 標準誤(Mean±S.E)來表示。
2 研究結果
2.1 不同時間尺度的家域
羚牛年家域面積的個體變異度較高。依據工作超過12個月的頸圈定位數據(n=5),羚牛平均家域為(15.01±2.92)km2/(9.02±1.85) km2(MCP/90%FKE),最大家域達31.61km2/21.02 km2,最小家域僅為3.71km2/1.84 km2(表1)。不論采用MCP法還是FKE法進行家域估計,個體年家域面積未表現出明顯的變化模式,但FKE家域的年際波動較MCP家域波動幅度小。
K-S檢驗顯示,兩種估算方法獲得的季節家域面積均符合正態分布。檢驗結果顯示,羚牛各季節間的家域面積存在顯著差異(MCP:F=3.008,df=3,P=0.041; FKE: F=3.101, df=3,P=0.038)。對MCP家域進行成對檢驗,結果顯示羚牛的春季家域((7.02±1.49)km2,n=9)顯著大于秋季((3.97±0.63)km2,n=14,P=0.044)和冬季的家域((3.64±0.62)km2,n=14,P=0.026),夏季家域((6.79±1.63) km2,n=9),雖然大于秋季但沒有顯著差別(P=0.061);夏季家域面積則顯著大于冬季的家域(P=0.037)。春季家域與夏季家域也無顯著差異(P=0.89)。對FKE家域進行成對檢驗,同樣顯示冬季家域(3.81±0.41,n=11)顯著小于春季家域(9.41±2.22,n=9,P=0.012),也小于夏季家域(8.01±2.08)且統計檢驗接近顯著性水平(P=0.056);冬季家域與秋季家域(4.82±1.01,n=12)無顯著差異。春季家域與夏季家域沒有顯著差異,但顯著大于秋季家域(P=0.034);其他季節間家域差異不顯著。
2.2 家域忠誠度
2.2.1 MCP法
相同季節的家域在年際間重疊度超過36%,家域的空間位置在年際間是穩定的(圖1)。秋季和夏季家域的年際間忠誠度最高,分別為(56.27±8.29)%(n=13)和(55.75±21.82)%(n=4);冬季家域年際忠誠度最低((36.34±6.82)%,n=13),春季家域忠誠度也相對較低((41.85±20.72)%,n=4)。雖然年際的家域忠誠度存在高低之分,但K個獨立樣本的非參數檢驗顯示季節家域忠誠度并無顯著差異。
2.2.2 FKE法
年際家域忠誠度較高,重疊比例為(72.54±7.99)%(n=8),編號682個體的家域重疊最高(94.9%),最低的是編號808的個體(27.21%)。頸圈工作時間超過3a的兩只羚牛(編號1534)和(編號1532)的相鄰年度家域重疊比例分別大于70%和85%。年際相同季節的家域重疊比例最小的僅8%,最大的達100%,均值為(61.3±5)%(n=24)。按季節重疊比例由小到大的次序依次是冬季((51.41±10.35)%,n=7)、春季((58.61±12.99)%,n=4)、秋季((64.42±8.68)%,n=9)和夏季((72.64±12.25)%,n=4)。然而,One way ANOVA檢驗結果顯示季節家域忠誠度在不同季節間并沒有顯著差異。
FKE年際家域的多邊形質心間距離為(0.47±0.19)km(n=6)。各季節年際間質心距離沒有顯著差異(df=3,F=1.71,P=0.202, One way ANOVA),但質心距離均值由小到大的順序是秋季((0.43±0.13)km,n=7)、夏季((0.75±0.28)km,n=4)、冬季((0.81±0.21)km,n=6)和春季((0.98±0.16)km,n=4)。
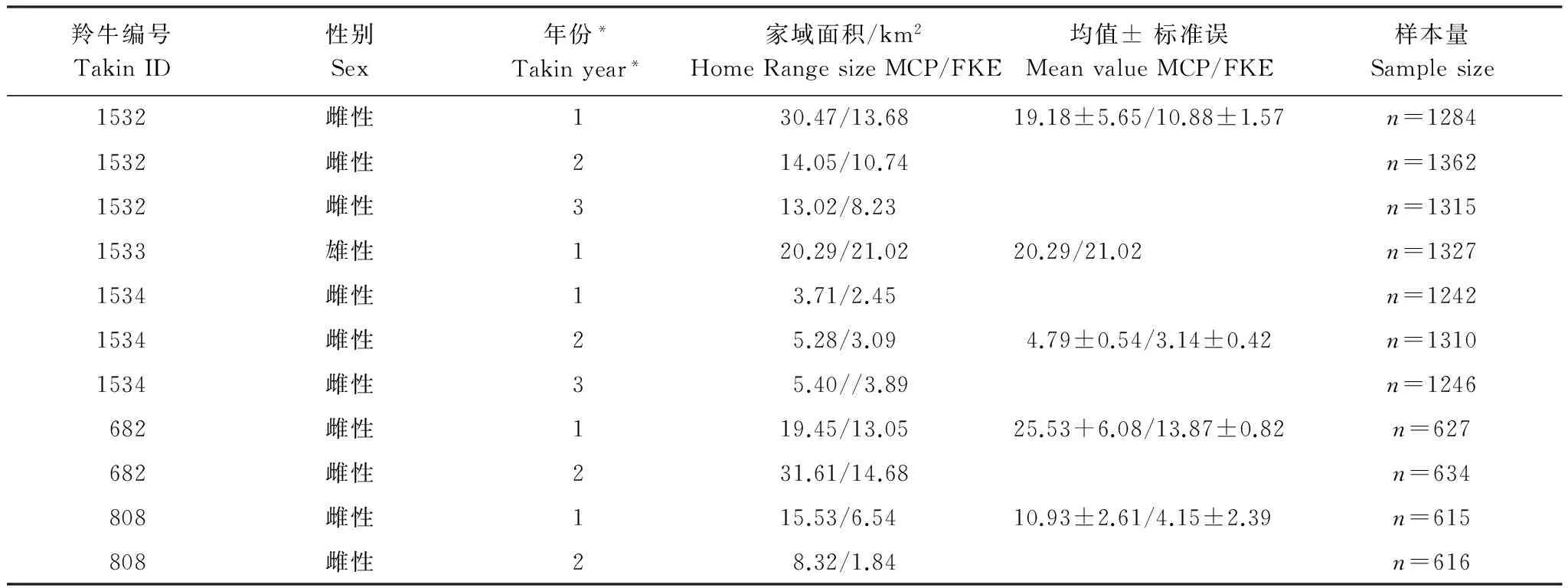
表1 唐家河自然保護區羚牛家域面積及年際變化(MCP/FKE)Table 1 Takin Home range size variation among years in Tangjiahe nature reserve (MCP/FKE)
*此處年份指從羚牛捕捉佩戴項圈起往后連續的12個月; MCP 和FKE 分別指最小凸多邊形法和固定核域法
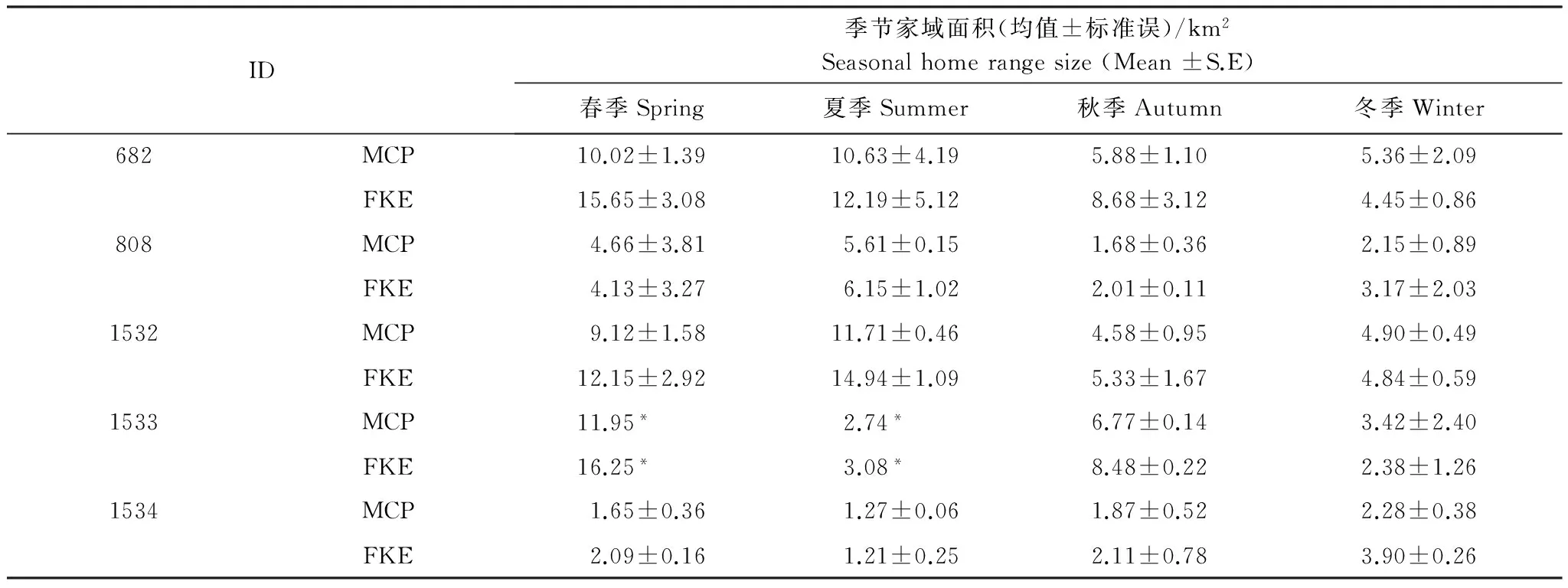
表2 唐家河自然保護區羚牛季節性家域面積(MCP/FKE)Table 2 Takin seasonal home range size in Tangjiahe nature reserve (MCP/FKE)
*表示雄性羚牛(編號1533)在春季和夏季的家域均只有1個樣本,即項圈的整個工作期內僅包括了1個春季和1個夏季;MCP:最小多邊形法Minimum convex polygon,FKE:固定核域法Fixed kernel estimation

圖1 唐家河自然保護區羚牛年家域多邊形(最小凸多邊形)Fig.1 Polygons of takin annual home range in Tangjiahe nature reserve (MCP)
3 討論
四川羚牛家域的季節變化顯著,其波動特征與秦嶺亞種相似。然而,四川亞種的家域面積(MCP法)不論是年家域還是季節家域均遠小于秦嶺亞種[20](年家域15.01km2vs 56.8km2;春季7.02 km2vs. 26.9 km2;夏季6.79 km2vs. 19.5 km2;秋季3.97 km2vs. 22.1 km2;冬季3.64 km2vs. 11.1 km2)。羚牛亞種間家域差異如此巨大,存在以下可能:1)由于兩亞種研究方法的不同,即數據獲取方式的不同引起的差異(無線電三角定位vs. GPS自動定位);2)秦嶺亞種研究區域的種群密度要遠小于唐家河區域的四川羚牛種群密度;3)秦嶺亞種羚牛體型顯著大于四川亞種個體;4)秦嶺區域羚牛的適宜棲息地斑塊分散。
家域概念最早由Burt提出,經Kernohan的總結后表述為:在一定的時間范圍內, 特定的動物出現概率的區域范圍[1,21]。早期的家域研究著重描述動物的活動范圍,后期逐漸強調對不同區域的利用強度[22]。最小凸多邊形法(MCP)是使用最早、最廣泛的家域估算方法,但由于該方法存在異常點干擾及不能反映家域內部利用強度的差異,其合理性受到了較多質疑[21,23]。相比較,由于概率模型可以反映動物對不同空間的利用強度,核域法是目前最受青睞的估算方法[21,23]。本研究使用最小凸多邊形法及核域法估算家域是基于研究結果既可與往研究比較,也可發現羚牛對不同空間利用強度的差異的考慮。此外,GPS頸圈應用使定位數據質量的穩定性得以保障,而數據精度也成為目前約束無線電定位法廣泛應用主要因素,因為有研究證實其定位誤差可能達500m[24]。因此,定位手段的差異也可能是兩亞種家域差異較大的主要原因之一。
曾治高等的研究區域是以佛坪自然保護區為核心[18],其主要特征就是周邊適宜棲息地逐漸減少迫使羚牛向佛坪集中(1.29—1.56只/km2,)[25],且種群密度應處于繼續增大的狀態。唐家河羚牛種群密度的報道僅見于20世紀80年代末(1.2—1.3只/km2)[26],至今未見數據更新,因此無有力證據完全排除由于兩個研究區域羚牛種群密度差異導致家域差異的可能。秦嶺亞種與四川亞種外形的差異主要體現在毛色而非體重,兩亞種成體重量主要在300—400kg范圍內[27],因此體重的差異也無法很好地解釋這一現象。雖然唐家河所在緯度比佛坪自然保護區低(32°32′—32°41′vs.33°33′—33°46′),但是兩個區域的氣候差異并不明顯(唐家河vs.佛坪,年均溫12℃ vs.11.5℃;最冷月均溫-1.2℃ vs.0.3℃;最熱月均溫19.7℃ vs.21.9℃,年日照時數1337.6h vs.1726.5h)[17, 20],不能得到唐家河的食物資源優于佛坪的結論并據此分析家域的差異。至于唐家河與佛坪自然保護區羚牛適宜棲息地分布是否存在差異還需要棲息地利用及評價方面的研究結果支持,但目前并無此類結果可供參考。因此,不能排除適宜棲息地分布特征引起的兩個亞種家域差異的可能。
家域的位置和面積波動是動物對環境變化的應答,是生存策略的具體化。在食物質量高且分布集中的區域,動物不需要進行大范圍覓食,因此家域也較小(如夏季家域),反之家域面積則較大(冬季家域)[28]。羚牛食物(質與量)的季節波動顯著[29-30]。本研究僅兩只雌性羚牛夏季家域和冬季家域變化能夠符合這一假說,說明影響動物家域的因素比已知的復雜。宋延齡等對秦嶺羚牛的家域研究認為羚牛季節家域的很大一部分是由遷移造成的[31],即其中的有很大部分屬于動物路過而非長期選擇利用的生境。因為統一的季節劃分標準并不一定適用于所有個體,例如某些個體的夏季家域可能是包含了其遷移過程中的棲息地,而有些個體的夏季家域則是已經完成遷移后相對穩定的家域。本研究中的5只羚牛,其中4只羚牛的夏季家域大于春季或近似于春季家域,且均為雌性。相比較,雄性羚牛的夏季家域僅為春季家域的1/4左右。可能的解釋是,雄性個體的遷移完成的比雌性羚牛更早。然而,至今尚無研究支持羚牛的遷移模式存在性間差異這一論點,但這種現象被證實在狍(Capreoluscapreolus)的遷移模式中是存在的[32]。此外,一些研究認為身體狀況和是否帶幼崽也對有蹄類的家域忠誠度和棲息地選擇有顯著的效應[33,29]。Mysterud曾提出動物在夏季增大活動范圍而獲取的能量效率要高于冬季,但不論冬季或夏季,動物的體重與其所占有的家域是顯著正相關的觀點[4]。羚牛平均體重在250—500kg,其冬季家域面積不足4km2,而生活在北美州,體重約60—130kg的白尾鹿(Odocoileusvirginianus)的冬季家域面積達10—12km2[31],可見體重和家域面積的關系不能作為唯一判斷標準,還應參考食物資源的質量、數量以及空間分布特征。因此,羚牛的個體差異包括繁殖狀態[34]、體形差異[22]及年齡差異[35]等可能是羚牛家域波動個體間差異較大的主要原因。
家域忠誠度高有利于提高個體的適合度,熟悉和適應一個區域將提高動物的生存機會和繁殖成功率,如更快地找到覓食地和逃生路線[6]。在較大的空間(時間)尺度如分布區(年度),每輪遷移周期的棲息地忠誠度可能接近100%,但是在更小的空間或時間尺度(如家域和季節),這種重疊度可能會在0和100%之間波動[18]。因此,同一物種對棲息地的忠誠度的高低,可以在一定程度上反映棲息地質量的優劣和穩定程度。例如,對馬鹿(Cervuselaphus)和駝鹿(Alcesamericanus)的研究就提出人類干擾應作為影響有蹄類對棲息地忠誠度的重要因子[36-37]。唐家河自然保護區的羚牛年際間的家域忠誠度,在食物較豐盛的季節(夏季和秋季)較高,達到64% 以上,且總體家域重疊度超過72%,這顯示了該保護區適宜羚牛生存且質量穩定。因此,對棲息地忠誠度研究的意義在于了解影響物種分布的關鍵棲息地因子,進而通過監測棲息地關鍵因子的波動和動物的行為反饋,一方面能夠探討動物行為策略形成的機制,另一方面也能應用于物種的保護和管理。雖然研究期間(2006—2009)唐家河自然保護區的人為干擾強度較低,不具備探討人為干擾對羚牛家域忠誠度的產生的影響及動物可能做出響應的條件。但是,目前唐家河自然保護區已經開展了生態旅游(自2011年),隨著開發的力度增加,人為活動給羚牛帶來的干擾可能日益顯現。因此,非常有必要在生態旅游起步階段開展人為活動對羚牛行為影響的監測工作,為有效地提升保護區對羚牛及其它野生動物保護和管理,提供數據支持。
由于本研究的樣本量較少,而且僅局限于家域行為和忠誠度的描述,缺乏相關生態數據的分析探討。未來的研究,需要聯系環境的屬性和變化以及不同策略間的生態后果與家域及忠誠度變化,探究影響羚牛及有蹄類的行為策略的深層原因。
[1] Burt W H. Territoriality and home range concepts as applied to mammals. Journal of Mammalogy, 1943, 24(3): 346-352.
[2] Mattisson J, Sand H, Wabakken P, Gervasi V, Liberg O, Linnell J D C, Rauset G R, Pedersen H C. Home range size variation in a recovering wolf population: evaluating the effect of environmental, demographic, and social factors. Oecologia, 2013, 173(3): 813-825.
[3] Milton, K, May M L. Body weight, diet and home range area in primates. Nature, 1976, 259(5543): 459-462.
[4] Mysterud A, Pérez-Barbería F J, Gordon I J. The effect of season, sex and feeding style on home range area versus body mass scaling in temperate ruminants. Oecologia, 2001, 127(1): 30-39.
[5] Jerina K. Roads and supplemental feeding affect home-range size of Slovenian red deer more than natural factors. Journal of Mammalogy, 2012, 93(4): 1139-1148.
[6] Ruther J, Thal K, Steiner S. Pheromone communication inNasoniavitripennis: abdominal sex attractant mediates site fidelity of releasing males. Journal of Chemical Ecology, 2011, 37(2): 161-165.
[7] Green B C, Smith D J, Grey J, Underwood G J. High site fidelity and low site connectivity in temperate salt marsh fish populations: a stable isotope approach. Oecologia, 2012, 168(1): 245-255.
[8] Matthews K R, Preisler H K. Site fidelity of the declining amphibianRanasierrae(Sierra Nevada yellow-legged frog). Canadian Journal of Fisheries and Aquatic Sciences, 2010, 67(2): 243-255.
[10] Hoover J P. Multiple effects of brood parasitism reduce the reproductive success of prothonotary warblers,Protonotariacitrea. Animal Behaviour, 2003, 65(5): 923-934.
[11] Hestbeck J B, Nichols J D, Malecki R A. Estimates of movement and site fidelity using mark-resight data of wintering Canada geese. Ecology, 1991, 72(2): 523-533.
[12] Xu J L, Zhang XH, Sun Q H, Zheng G M, Wang Y, Zhang Z W. Home range, daily movements and site fidelity of male Reeves′s pheasantsSyrmaticusreevesiiin the Dabie Mountains, central China. Wildlife Biology, 2009, 15(3): 338-344.
[13] van Beest F M, Vander Wal E, Stronen A V, Paquet P C, Brook R K. Temporal variation in site fidelity: scale-dependent effects of forage abundance and predation risk in a non-migratory large herbivore. Oecologia, 2013, 173(2): 409-420.
[14] Liu Y, Keller I, Heckel G. Breeding site fidelity and winter admixture in a long-distance migrant, the tufted duck (Aythyafuligula). Heredity, 2012, 109(2): 108-116.
[15] Costello C M. Estimates of dispersal and home-range fidelity in American black bears. Journal of Mammalogy, 2010, 91(1): 116-121.
[16] Faille G, Dussault C, Ouellet J P, Fortin D, Courtois R, St-Laurent M H, Dussault C. Range fidelity: The missing link between caribou decline and habitat alteration?. Biological Conservation, 2010, 143(11): 2840-2850.
[17] Guan T P, Ge B M, McShea W J, Li S, Song Y L, Stewart C M. Seasonal migration by a large forest ungulate: a study on takin (Budorcastaxicolor) in Sichuan Province, China. European Journal of Wildlife Research, 2013, 59(1): 81-91.
[18] Zeng Z G, Skidmore A K, Song Y L, Wang T J, Gong H S. Seasonal altitudinal movements of golden takin in the Qinling Mountains of China. Journal of Wildlife Management, 2008, 72(3): 611-617.
[19] 曾治高, 鐘文勤, 宋延齡, 李俊生, 趙雷剛, 鞏會生. 羚牛生態生物學研究現狀. 獸類學報, 2003, 23(2): 161-167.
[20] 宋延齡, 曾治高, 張堅, 王學杰, 鞏會生, 王寬武. 秦嶺羚牛的家域研究. 獸類學報, 2000, 20(4): 241-249.
[21] Kernohan B J, Gitzen R A, Millspaugh J J. Analysis of animal space use and movements // Milspaugh J J, Marzluff J M, eds. Radio tracking and animal populations. New York: Academic, 2001: 125-166.
[22] B?rger L, Franconi N, Ferretti F, Meschi F, De Michele G, Gantz A, Coulson T. An integrated approach to identify spatiotemporal and individual-level determinants of animal home range size. The American Naturalist, 2006, 168(4): 471-485.
[23] Laver P N, Kelly M J. A critical review of home range studies. Journal of Wildlife Management, 2008, 72(1): 290-298.
[24] Rodgers A R, Rempel R S, Abraham K F. A GPS-based telemetry system. Wildlife Society Bulletin, 1996, 24(3): 559-566.
[25] 曾治高, 宋延齡, 鞏會生. 佛坪自然保護區羚牛的種群數量與結構特征. 獸類學報, 1998, 18(4): 241-246.
[26] 葛桃安, 胡錦矗, 江明道, 鄧啟濤. 唐家河自然保護區扭角羚的獸群結構及數量分布. 獸類學報, 1989, 9(4): 262-268.
[27] 吳家炎. 中國羚牛. 北京: 中國林業出版社, 1990.
[28] McCullough D R. Density dependence and life-history strategies of ungulates. Journal of Mammalogy, 1999, 80(4): 1130-1146.
[29] 曾治高, 宋延齡, 鐘文勤, 鞏會生, 張堅, 黨高第. 秦嶺羚牛的食性. 動物學雜志, 2001, 36(3): 36-44.
[30] Schaller G B, Teng Q T, Pan W S, Qin Z S, Wang X M, Hu J C, Shen H M. Feeding behavior of Sichuan takin (Budorcastaxicolor). Mammalia, 1986, 50(3): 311-322.
[31] Lesage L, Crête M, Huot J, Dumont A, Ouellet J P. Seasonal home range size and philopatry in two northern white-tailed deer populations. Canadian Journal of Zoology, 2000, 78(11): 1930-1940.
[32] Mysterud A. Seasonal migration pattern and home range of roe deer (Capreoluscapreolus) in an altitudinal gradient in southern Norway. Journal of Zoology, 1999, 247(4): 479-486.
[33] Luccarini S, Mauri L, Ciuti S, Lamberti P, Apollonio M. Red deer (Cervuselaphus) spatial use in the Italian Alps: home range patterns, seasonal migrations, and effects of snow and winter feeding. Ethology Ecology & Evolution, 2006, 18(2): 127-145.
[34] van Beest F M, Rivrud I M, Loe L E, Milner J M, Mysterud A. What determines variation in home range size across spatiotemporal scales in a large browsing herbivore? Journal of Animal Ecology, 2011, 80(4): 771-785.
[35] Sa?d S, Gaillard J M, Widmer O, Débias F, Bourgoin G, Delorme D, Roux C. What shapes intra-specific variation in home range size? A case study of female roe deer. Oikos, 2009, 118(9): 1299-1306.
[36] Tracz B V, LaMontagne J M, Bayne E M, Boutin S. Annual and monthly range fidelity of female boreal woodland caribou in response to petroleum development. Rangifer, 2010, 30(1): 31-44.
[37] Webb S L, Dzialak M R, Harju S M, Hayden Wing L D, Winstead J B. Influence of land development on home range use dynamics of female elk. Wildlife Research, 2011, 38(2): 163-167.
Home range and fidelity of Sichuan takin
GUAN Tianpei1,GE Baoming2,CHEN Limin3,YOU Zhangqiang1,TANG Zhonghai1,LIU Hao1,SONG Yanling4,*
1EcologicalSecurityandProtectionKeyLaboratoryofSichuanProvince,MianyangNormalUniversity,Mianyang621000,China2JiangsuProvincialKeyLaboratoryofCoastalWetlandBioresourcesandEnvironmentalProtection,YanchengTeachersUniversity,Yancheng224002,China3TangjiaheNationalNatureReserve,Qingchuan628109,China4KeyLaboratoryofAnimalEcologyandConservationBiology,InstituteofZoology,ChineseAcademyofSciences,Beijing100101,China
Migrant and resident are observed to return or stay within a specific range of habitat, close to or completely overlap with its original distribution area, known as habitat fidelity. In this study, we estimated takin seasonal and annual home range size and its fidelity between years by utilizing 9 GPS radio collars on adult takin(2006—2009). Results shown annual home range of takin was (MCP/FKE)(15.01±2.92)km2/(9.02±1.85) km2, suggesting individual home range varied among individuals and years. For each individual, we found variation of seasonal home range between years. Most of individuals shown similar pattern of seasonal home range variation, thus the largest home range always found in summer or spring. We extracted the centriods of seasonal / annual home range polygon and calculate its distance between years. We tried the distance between two centriods as one surrogate of home range fidelity. Besides, we considered the overlap ratio of home range between two consecutive years as the most important parameter of home range fidelity.Therefore, we found annual home range fidelity of certain season variation exist, especially for summer and autumn. Both ways of home range fidelity assessment produced similar results.
minimum convex polygon; fixed kernel estimation; ungulate; sichuan takin; home range
國家自然科學青年基金項目 (31300319);綿陽師范學院啟動項目 (QD2012A12)
2013-03-04;
2014-03-04
10.5846/stxb201403040372
*通訊作者Corresponding author.E-mail: songyl@ioz.ac.cn
官天培,葛寶明,諶利民,游章強,唐中海, 劉昊,宋延齡.四川羚牛的家域與忠誠度.生態學報,2015,35(6):1862-1868.Guan T P,Ge B M,Chen L M,You Z Q,Tang Z H,Liu H,Song Y L.Home range and fidelity of Sichuan takin.Acta Ecologica Sinica,2015,35(6):1862-1868.
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
