魁蚶(Scapharca broughtonii)半乳糖凝集素(SbGal)基因cDNA的克隆及表達(dá)分析*
2015-03-22 00:58:47鄭利兵劉志鴻周麗青孫秀俊田吉騰楊愛國鄭言鑫
海洋與湖沼 2015年5期
鄭利兵 吳 彪 劉志鴻① 周麗青 孫秀俊田吉騰 楊愛國 鄭言鑫
(1. 農(nóng)業(yè)部海洋漁業(yè)可持續(xù)發(fā)展重點(diǎn)實(shí)驗(yàn)室 中國水產(chǎn)科學(xué)研究院黃海水產(chǎn)研究所 青島 266071;2. 上海海洋大學(xué)水產(chǎn)與生命學(xué)院 上海 201306)
凝集素(lectin)是生物體內(nèi)廣泛存在的一類能使細(xì)胞發(fā)生凝集的糖類結(jié)合蛋白。作為軟體動(dòng)物體液免疫的一個(gè)重要免疫因子, 凝集素在貝類免疫防御中起到異體成分的識(shí)別、調(diào)理、凝集等作用, 是構(gòu)成機(jī)體防御體系的重要組成部分(翟玉梅等, 1998; Weisset al, 2000; 孫虎山等, 2001; 胥煒等, 2005)。
半乳糖凝集素(galectin)是動(dòng)物凝集素家族的成員之一(Gerardoet al, 2009), 可特異性識(shí)別糖鏈末端的 β-半乳糖苷(賀雪明等, 2012), 為非膜整合性的可溶性蛋白(Barondes, 1984; Drickameret al, 1985)。在貝類等無脊椎動(dòng)物發(fā)生感染時(shí), 半乳糖凝集素通過其標(biāo)志性的糖識(shí)別結(jié)構(gòu)域(Carbohydrate Recognition Domain, CRD)特異性結(jié)合半乳糖苷來參與對(duì)病害的免疫反應(yīng)(Dumicet al, 2006; 陳騁, 2013)。例如, 菲律賓蛤仔(Ruditapes philippinarum)的半乳糖凝集素可結(jié)合派琴蟲(Perkinsus)和弧菌(Vibrio)表面的半乳糖和 N-乙酰半乳糖胺而進(jìn)行病原菌的識(shí)別(Kimet al,2008); 海鰻的原型galectin參與細(xì)胞對(duì)寄生蟲的包囊作用(Nakamuraet al, 2012); 大西洋鱈魚galectin-1能凝集革蘭氏陽性和陰性菌, 參與其免疫防御過程(Rajanet al, 2013)。因此, 半乳糖凝集素作為模式識(shí)別受體(Pattern Recognition Molecular, PRR)發(fā)揮作用的功能得到了廣泛研究。
Galectin 家族成員眾多, 在一種動(dòng)物中可能含有不同的類型, 即使在同一種細(xì)胞中也可能會(huì)表達(dá)出幾種不同的galectin (Colnotet al, 1996)。近年來,對(duì)軟體動(dòng)物半乳糖凝集素的研究中發(fā)現(xiàn)一種獨(dú)特的quadruple-galectin (Tasumiet al, 2007; Songet al, 2010;Zhanget al, 2011), 其對(duì)半乳糖凝集素家族的結(jié)構(gòu)是一個(gè)新的補(bǔ)充。但有關(guān)該類型galectin的報(bào)道還很少, 第一個(gè)被鑒定的 quadruple-galectin是來自于美洲牡蠣(Crassostrea viginica), 它可以識(shí)別并吞噬原生動(dòng)物鞭孢簇蟲(Perkinsus marinus)(Tasumiet al, 2007); 之后被鑒定的 quadruple-galectin還有 PoGal (Zhanget al,2011)、AiGal1 (Songet al, 2010)和 AiGal2 (Songet al,2011), 經(jīng)過病原微生物的免疫刺激, 這些galectin的表達(dá)量都有明顯的上調(diào)。然而, 目前尚未有魁蚶(Scapharca broughtonii)半乳糖凝集素的相關(guān)報(bào)道。
魁蚶屬于雙殼綱, 蚶目, 具有適應(yīng)性強(qiáng)、生態(tài)修復(fù)效果顯著的特點(diǎn), 但近年來資源量銳減, 深入挖掘魁蚶免疫因子對(duì)其產(chǎn)業(yè)的發(fā)展具有重要的理論和現(xiàn)實(shí)意義。目前, 本課題組已在魁蚶血細(xì)胞分類及免疫功能研究(周麗青等, 2013, 2014)及種質(zhì)資源和遺傳多樣性分析(吳彪等, 2010, 2012; 梁超等, 2011; 田吉騰, 2012; 周麗青等, 2012)等方面進(jìn)行了研究, 發(fā)現(xiàn)鰻弧菌(Vibrio anguillarum)能夠啟動(dòng)魁蚶的免疫防御功能。為進(jìn)一步明確魁蚶半乳糖凝集素的特征和免疫作用, 本研究在魁蚶轉(zhuǎn)錄組文庫中篩選到的半乳糖凝集素序列的基礎(chǔ)上, 經(jīng)過 RACE 擴(kuò)增得到 cDNA全長序列, 并開展基因表達(dá)組織分析和鰻弧菌刺激響應(yīng)規(guī)律的研究, 以期為魁蚶分子育種及抗病選育提供依據(jù), 為認(rèn)識(shí)魁蚶半乳糖凝集素和對(duì)該分子的進(jìn)化、結(jié)構(gòu)和功能的研究提供資料。
1 材料與方法
1.1 實(shí)驗(yàn)材料與來源
實(shí)驗(yàn)用魁蚶購于青島臺(tái)東水產(chǎn)品市場(chǎng), 殼長約55mm左右, 個(gè)體健康, 于20°C充氣海水中暫養(yǎng)一周,每天換水兩次。期間投喂硅藻, 實(shí)驗(yàn)處理前兩天停止投喂。鰻弧菌為本實(shí)驗(yàn)室保存的菌種。
1.2 魁蚶半乳糖凝集素基因的獲得
1.2.1 總 RNA提取 異硫氰酸胍法提取總 RNA:采用Solution D (4mol/L異硫氰酸胍, 17mmol/L月桂酰肌苷酸鈉, 25mmol/L檸檬酸鈉)與β-巰基乙醇裂解組織, 然后相同體積氯仿/異戊醇(24︰1)和水飽和酚抽提蛋白質(zhì), 異丙醇與醋酸鈉沉淀核酸, 75%乙醇洗滌核酸后干燥, 用 DNase(Progema)來去除基因組DNA, 具體步驟參照董迎輝(2012)。1.0%瓊脂糖檢測(cè)RNA 完整性, 核酸分析儀(Eppendorf)檢測(cè)其純度和濃度。
1.2.2 全長 cDNA克隆及測(cè)序 根據(jù)獲得的魁蚶半乳糖凝集素基因的部分序列, Primer Premier 5.0設(shè)計(jì) 3'RACE和 5'RACE特異性引物, 利用 SMART?RACE Amplication Kit和Advantage 2 PCR Kit用于3'端和5'端的擴(kuò)增。3'RACE 擴(kuò)增所用引物為SbGal-3F和通用引物UPM, 5'RACE擴(kuò)增所用引物為SbGal-5R和通用引物UPM(表1)。具體步驟如下: 擴(kuò)增體系為50μL, 內(nèi)含 3'(5')RACE cDNA模板 2.5μL, PCR-Grade water 34.5μL, 10× Advantage2 PCR buffer 5.0μL,dNTP Mix(10mmol/L)1.0μL, UPM 5.0μL, SbGal-3F(SbGal-5R) 1.0μL; 反應(yīng)程序?yàn)? 94 °C 30 s, 68 °C 30 s,72 °C 2 min, 25個(gè)循環(huán)。PCR產(chǎn)物經(jīng)1.2%瓊脂糖凝膠電泳檢測(cè)、切膠回收, 回收產(chǎn)物分別與 PMD18-T連接, 轉(zhuǎn)化大腸桿菌Top10感受態(tài)細(xì)胞, 并挑陽性克隆進(jìn)行測(cè)序。
1.2.3 序列分析 將篩選到的序列及推測(cè)的氨基酸序列與GenBank數(shù)據(jù)庫中的已知序列進(jìn)行blast比對(duì); DNAStar軟件對(duì)cDNA序列進(jìn)行開放閱讀框搜索及編碼氨基酸預(yù)測(cè); 利用 ProtParam tool (http://web.expasy.org/protparam/)對(duì)半乳糖凝集素分子量(Mw)及等電點(diǎn)pI進(jìn)行預(yù)測(cè); SignalIP 4.0 Server (http://www.cbs.dtu.dk/services/SignalP/)查找半乳糖凝集素信號(hào)肽; TargetP 1.1 (http: //www.cbs.dtu.dk/services/TargetP/)對(duì)半乳糖凝集素進(jìn)行亞細(xì)胞定位; TMHMM Server v.2.0 (http: //www.cbs.dtu.dk/services/TMHMM/)預(yù)測(cè)蛋白質(zhì)跨膜區(qū)域; SMART (http: //www.expasy.ch/SMART)進(jìn)行蛋白質(zhì)結(jié)構(gòu)域分析。用 NCBI的Blastp (http: //blast.ncbi.nlm.nih.gov/)對(duì)序列進(jìn)行同源檢索, 取不同分類物種以及不同種類半乳糖凝集素氨基酸序列, 用DNA man8.0軟件與魁蚶鐵蛋白氨基酸序列進(jìn)行同源性比對(duì)分析, 在此基礎(chǔ)上用 MEGA 5.0構(gòu)建系統(tǒng)發(fā)育樹。
1.3 半乳糖凝集素組織分布及對(duì)細(xì)菌刺激的響應(yīng)模式分析
1.3.1 鰻弧菌刺激實(shí)驗(yàn) 挑選健康的暫養(yǎng)魁蚶并隨機(jī)分為對(duì)照組和處理組進(jìn)行鰻弧菌刺激實(shí)驗(yàn)。鰻弧菌經(jīng) 2216E培養(yǎng)基擴(kuò)大培養(yǎng)后離心取沉淀, 用 PBS調(diào)整濃度至OD600=0.4 (1OD=5×108bacteria/mL)。微量注射器向處理組個(gè)體前閉殼肌處注射50μL菌懸液后放回原養(yǎng)殖槽內(nèi), 分別在0h、4h、8h、16h、24h、32h、64h隨機(jī)選取 3個(gè)個(gè)體活體取血(4°C,800r/min15min收集血細(xì)胞沉淀)、外套膜、鰓、閉殼肌、肝胰腺、斧足, 用于總 RNA的提取。對(duì)照組注射同劑量的滅菌PBS(0.1 mol/L, pH 7.4)代替鰻弧菌。
1.3.2 實(shí)時(shí)熒光定量 PCR(qRT-PCR) 運(yùn)用qRT-PCR檢測(cè)半乳糖凝集素轉(zhuǎn)錄本的組織分布情況及對(duì)鰻弧菌刺激的反應(yīng)模式。根據(jù)獲得的半乳糖凝集素 cDNA序列, 用 Primer Premier 5.0軟件設(shè)計(jì)qRT-PCR引物 qSbGal-F、qSbGal-R(引物信息見表 1),選取β-actin基因作為內(nèi)參基因(Liet al, 2012), 反應(yīng)在ABI7500熒光定量PCR儀上完成。利用PrimeScriptTMRT reagent Kit (TaKaRa)試劑盒進(jìn)行反轉(zhuǎn)錄合成模板cDNA, 反應(yīng)體積及反應(yīng)條件按照說明書進(jìn)行。反轉(zhuǎn)錄產(chǎn)物置于–20°C保存?zhèn)溆谩RT-PCR擴(kuò)增體系為20μL, 包括: SYBR Premix Ex TaqTMⅡ(2×), 10μL;PCR Forward Primer(10μmol/L), 0.8μL; PCR Reverse Primer (10μmol/L), 0.8μL; ROX Reference DyeⅡ(50×),0.4μL; cDNA 模板, 2.0μL; DEPC 水, 6.0μL。反應(yīng)程序?yàn)?95°C 30 s; 95°C 5 s, 60°C 34 s, 40 個(gè)循環(huán), 95°C 15 s,60°C 1 min, 95°C 15s。實(shí)驗(yàn)設(shè)置 3 個(gè)平行組。1.3.3 數(shù)據(jù)分析 軟件獲得每個(gè)反應(yīng)的 CT, 采用2-DDCT方法(Livaket al, 2001)進(jìn)行相對(duì)定量分析。根據(jù)每個(gè)樣品的 3個(gè)平行實(shí)驗(yàn)所得的數(shù)據(jù)計(jì)算相對(duì)表達(dá)量平均值和標(biāo)準(zhǔn)差。

表1 實(shí)驗(yàn)所用引物序列Tab.1 The primers used in the experiment
2 結(jié)果
2.1 SbGal基因cDNA序列分析
3'RACE和5'RACE擴(kuò)增產(chǎn)物分別測(cè)序后與轉(zhuǎn)錄組文庫中篩選出的序列進(jìn)行拼接, 獲得魁蚶半乳糖凝集素全長cDNA序列, 命名為SbGal(GenBank登錄號(hào)為KP123598)。SbGal基因cDNA全長3120bp, 包含一個(gè)213 bp的5'-端非翻譯區(qū)(5'-UTR), 1342 bp的3'-UTR和1565 bp的開放閱讀框(ORF); 其起始密碼子為ATG, 終止密碼子為TGA, Poly(A)序列上游有4個(gè)規(guī)則的加尾信號(hào)AATAAA, 很明顯, 該cDNA具有完整的蛋白質(zhì)編碼區(qū)(圖1)。
氨基酸序列分析可知,SbGal基因編碼一個(gè)含有554個(gè)氨基酸殘基組成的蛋白質(zhì), 預(yù)測(cè)的分子量大小為63.3158kDa, 理論等電點(diǎn)為4.99。結(jié)構(gòu)特征分析結(jié)果表明: 該氨基酸序列無信號(hào)肽, 也非線粒體靶向肽,無明顯的跨膜區(qū)域; 該蛋白具有多個(gè)功能化位點(diǎn), 第186位NETC和319位NGTD存在潛在糖基化位點(diǎn);在137、218、278、279、382處有脾酪氨酸激酶磷酸化位點(diǎn), 279、382、388、526bp處有可激活胰島素受體激酶磷酸化位點(diǎn), 225位有共濟(jì)失調(diào)毛細(xì)血管擴(kuò)張突變激酶磷酸化位點(diǎn), 374位有蛋白激酶C磷酸化位點(diǎn), 218位有表皮生長因子受體磷酸化位點(diǎn); 結(jié)構(gòu)域的預(yù)測(cè)分析結(jié)果表明SbGal有四個(gè)糖識(shí)別結(jié)構(gòu)域(CRD), 且每個(gè)結(jié)構(gòu)域通過一含有 6個(gè)氨基酸的短肽鏈連接在一起, 每個(gè)CRD都含有保守的糖結(jié)合位點(diǎn),第一、第三和第四 CRD均含有 132個(gè)氨基酸殘基(12—143, 286—417, 424—555), 第二CRD含有130個(gè)氨基酸殘基(150—279)。綜上分析結(jié)果表明, 本實(shí)驗(yàn)成功獲得了魁蚶半乳糖凝集素基因全長cDNA。
2.2 SbGal基因同源性分析
同源性在線搜索結(jié)果顯示:SbGal編碼的氨基酸序列與其它雙殼類具有較高的相似性, 例如, 與美洲牡蠣Crassostrea viginica的相似性為70%, 合浦珠母貝Pinctada fucata為 65%, 海灣扇貝Argopectehs irradiaas為46%; 與腹足類的相似性相對(duì)較低, 與光滑雙臍螺Biompalaria glabrata兩個(gè)半乳糖凝集素氨基酸的相似性分別是30%、29%; 魁蚶半乳糖凝集素的四個(gè) CRD彼此的相似性在 36%—41%之間, 且CDR1與CDR4的相似性最大為41%。

圖1 SbGal核苷酸序列及其編碼氨基酸序列Fig.1 Nucleotide and deduced amino acid sequence of SbGal陰影部分為SbGal的四個(gè)結(jié)構(gòu)域, 加粗帶下劃線的為糖結(jié)合位點(diǎn), 雙下劃線分別代表起始密碼子和終止密碼子, 帶框的為加尾信號(hào), 波浪線表示多聚尾
DNAMAN8.0軟件對(duì)SbGal基因編碼的氨基酸序列與海灣扇貝、美洲牡蠣和合浦珠母貝的氨基酸序列進(jìn)行比對(duì)(圖2), 在SbGal四個(gè)CRD上都有保守的糖結(jié)合位點(diǎn), 不同物種的同一種 galectin在糖結(jié)合位點(diǎn)上都很保守, 在SbGal的第一CRD中, 這些保守序列分別是His57, Asn59, Arg61, Asn70, Trp76, Glu79和Arg81,在其它三個(gè)CRD中這些氨基酸殘基也同樣存在。
在 GenBank上搜索選取了具有代表性物種的半乳糖凝集素序列, 利用Mega5.0構(gòu)建NJ系統(tǒng)進(jìn)化樹如圖3, 整個(gè)系統(tǒng)發(fā)育樹分成4大支, 原型galectin分成原型1和2兩個(gè)分支, quadruple-galectin居于這兩原型分支間, 且與原型 galectin 1首先聚在一起; 在quadruple-galectin分支中,S. broughtonii首先與C.virginica聚在一起后再與P. fucata聚類。

圖2 不同物種Galectin氨基酸序列比對(duì)Fig.2 Multiple sequence alignment by DNAman 8.0框線內(nèi)為半乳糖凝集素的四個(gè)糖識(shí)別結(jié)構(gòu)域, 用方框標(biāo)出, 糖結(jié)合位點(diǎn)用三角標(biāo)出, 各物種GenBank登錄號(hào)為: 海灣扇貝A. irradians(ACS72240, FJ469998); 美洲牡蠣C. virginica (ABG75998); 合浦珠母貝P. fucata (ACO36044)
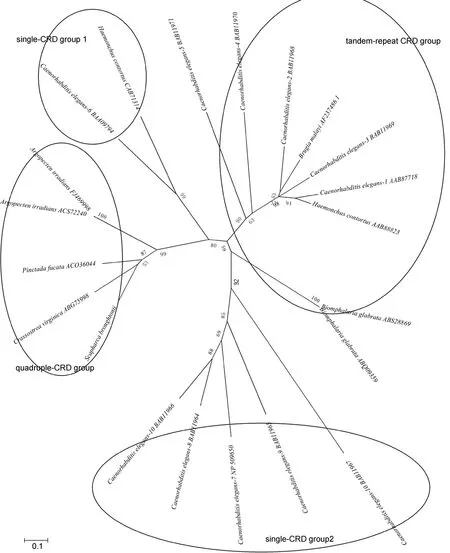
圖3 半乳糖凝集素的NJ系統(tǒng)進(jìn)化樹Fig.3 Phylogenetic tree of CRDs from 21 galectins
2.3 SbGal基因組織表達(dá)差異分析
以β-actin為內(nèi)參基因檢測(cè)了SbGalmRNA在健康個(gè)體不同組織中的表達(dá)差異性(圖 4), 結(jié)果表明:SbGalmRNA在斧足、鰓、外套膜、肝胰腺、閉殼肌、血細(xì)胞中都有表達(dá), 但表達(dá)量存在明顯的差異。以肝胰腺的表達(dá)量為參考,SbGal在血細(xì)胞中的表達(dá)量最高, 為肝胰腺的87.05倍(P<0.01), 其次是在斧足當(dāng)中,為72.86倍(P<0.01)。

圖4 SbGal在不同組織中的表達(dá)分布Fig.4 Distribution of SbGal gene in different tissues of S.broughtonii
2.4 SbGal在鰻弧菌感染后的表達(dá)模式
鰻弧菌感染后,SbGal在斧足、鰓、外套膜、肝胰腺、閉殼肌、血細(xì)胞 6種組織中的表達(dá)模式如圖5, 結(jié)果表明: 鰻弧菌感染后, 魁蚶 6種組織中的SbGal的表達(dá)量都明顯上調(diào)。感染4h時(shí), 在各組織中的表達(dá)量開始明顯高于對(duì)照組, 并且在肝胰腺和閉殼肌中, 其表達(dá)量達(dá)到最大值, 分別為對(duì)照組的1430.48倍(P<0.01)、1038.23倍(P<0.01); 在斧足和鰓中的表達(dá)量于感染后 8h達(dá)到最高值, 分別為對(duì)照組的128.21倍(P<0.01)、1502.70倍(P<0.01); 在外套膜和血細(xì)胞中的表達(dá)量于感染后16h達(dá)到最高值, 分別為對(duì)照組的 361.56倍(P<0.01)、354.87倍(P<0.01)。隨后, 各組織中的表達(dá)量開始回落, 且在鰓和閉殼肌中其表達(dá)量的回落表現(xiàn)出明顯的瞬時(shí)趨勢(shì);SbGal在肝胰腺中的表達(dá)量于感染后 32h出現(xiàn)二次表達(dá)高峰,為對(duì)照組的261.72倍(P<0.01)。總之, 經(jīng)鰻弧菌感染后,SbGal表現(xiàn)出顯著的時(shí)間依賴性, 除血細(xì)胞外還表現(xiàn)出較顯著地瞬時(shí)表達(dá)趨勢(shì), 總體表現(xiàn)為先升高后降低的趨勢(shì)。
3 討論
特異性結(jié)合 β-半乳糖苷的半乳糖凝集素是動(dòng)物凝集素的一種代表類型。半乳糖凝集素作為 PRR參與免疫防御, 病原微生物的刺激會(huì)調(diào)節(jié)半乳糖凝集素在生物體內(nèi)的表達(dá), 進(jìn)而加強(qiáng)免疫應(yīng)答過程(Vasta,2009)。近來, 有報(bào)道稱半乳糖凝集素結(jié)合在細(xì)胞表面和體內(nèi)潛在病原微生物細(xì)胞外基質(zhì)的多糖上, 在先天免疫過程中以識(shí)別效應(yīng)器的角色發(fā)揮作用(Vasta,2009)。目前為止, 對(duì)無脊椎動(dòng)物galectin的研究指出,quadruple-galectin僅在軟體動(dòng)物中發(fā)現(xiàn), 如AiGal1、PfGal、AiGal2已被成功鑒定并做結(jié)構(gòu)功能分析。本研究通過構(gòu)建魁蚶高通量測(cè)序轉(zhuǎn)錄組文庫并利用RACE技術(shù)獲得了魁蚶 galectin基因全長 cDNA; 檢測(cè)了其 mRNA在魁蚶中各組織的分布情況和在外源微生物刺激下表達(dá)量的變化, 對(duì)探討半乳糖凝集素在固有免疫防御中發(fā)揮的作用具有重要意義。
現(xiàn)在已鑒定分析的 galectin無信號(hào)肽, 通過非經(jīng)典的分泌方式到達(dá)細(xì)胞外(Almkvistet al, 2002)。SbGal編碼氨基酸序列的N端也不存在信號(hào)肽, 推測(cè)SbGal也是通過非經(jīng)典分泌方式到達(dá)細(xì)胞外, 但還需要進(jìn)一步的實(shí)驗(yàn)驗(yàn)證。SbGalcDNA的3'-UTR存在多個(gè)加尾信號(hào)的現(xiàn)象可能與脊椎動(dòng)物galectin一樣存在多種剪切方式可產(chǎn)生多個(gè)轉(zhuǎn)錄變異體, 皺紋盤鮑galectin基因cDNA的3'-UTR存在3處多聚腺苷酸加尾位點(diǎn)(鄭明剛, 2007)。
同源性比對(duì)分析發(fā)現(xiàn)SbGal的四個(gè)結(jié)構(gòu)域高度保守, 且每個(gè)結(jié)構(gòu)域中與糖結(jié)合的氨基酸殘基也都高度保守, 符合半乳糖凝集素家族成員的具有保守糖識(shí)別位點(diǎn)(Kasaiet al, 1996; Leffler, 2001)的結(jié)構(gòu)特征; NJ系統(tǒng)進(jìn)化樹看到SbGal與同屬于 quadruplegalectin的其它物種的galectin聚在一起, 總之, 成功獲得的SbGal也是一種quadruple- galectin。
綜上,SbGal與已知的無脊椎動(dòng)物半乳糖凝集素結(jié)構(gòu)相似, 沒有信號(hào)肽且只含有典型的半乳糖凝集素識(shí)別結(jié)構(gòu)域, 這一點(diǎn)與脊椎動(dòng)物的半乳糖凝集素結(jié)構(gòu)特征相一致(宋小燕, 2009)。特征性序列分析、較高的同源性、保守氨基酸殘基的存在都說明SbGal是半乳糖凝集素家族中的一個(gè)新成員, 其是在軟體動(dòng)物中發(fā)現(xiàn)的又一個(gè)具有四個(gè)明顯結(jié)構(gòu)域的半乳糖凝集素。因此, 本研究不僅填補(bǔ)了魁蚶半乳糖凝集素研究的空白, 而且還豐富了無脊椎動(dòng)物半乳糖凝集素研究的相關(guān)數(shù)據(jù)。
qRT-PCR檢測(cè)發(fā)現(xiàn)SbGal與其它雙殼貝類galectin一樣在健康個(gè)體的不同組織都有表達(dá)(Tasumiet al, 2007; Songet al, 2010, 2011; Zhanget al, 2011),但表達(dá)量存在差異, 說明SbGal基因的表達(dá)不具有組織特異性, 但表達(dá)量具有一定的組織特異性。有研究表明細(xì)菌、病毒、真菌或寄生蟲感染后的牡蠣oyster(Zhanget al, 2011)、文昌魚Branchiostoma lanceolatum(禹艷紅, 2007)和文蛤Meretrix meretrix(Kimet al, 2008)體內(nèi)的半乳糖凝集素基因mRNA水平上調(diào)。經(jīng)過鰻弧菌和藤黃微球菌(Micrococcus luteus)感染后的海灣扇貝, AiGal1 mRNA的表達(dá)量顯著上調(diào),然而, 經(jīng)酵母感染后, 其表達(dá)量基本保持不變(Songet al, 2010); 在正常情況下, 合浦珠母貝半乳糖凝集素 PoGal2持續(xù)表達(dá), 當(dāng)被溶藻弧菌(Vibrio alginolyticus)感染后, 其表達(dá)量也顯著上調(diào)(Zhanget al, 2011)。本研究中鰻弧菌感染魁蚶后,SbGal在各組織的表達(dá)量都有升高, 在肝胰腺中上升幅度最大, 可能SbGal通過肝胰腺發(fā)揮對(duì)病原菌入侵的防御作用較大(鄭明剛, 2007), 或者肝胰腺是軟體動(dòng)物先天免疫中合成蛋白的重要器官(Konget al, 2010; Prokhorovaet al,2010; Zhaoet al, 2010)。鰻弧菌感染后,SbGal前期的表達(dá)是逐步上調(diào)增強(qiáng)的, 再經(jīng)過一定的時(shí)間即恢復(fù)到感染前的水平, 由此可見SbGal的表達(dá)在時(shí)間和空間上都有一定的規(guī)律, 這為進(jìn)一步研究SbGal在魁蚶免疫反應(yīng)中的相關(guān)免疫功能研究提供了資料。
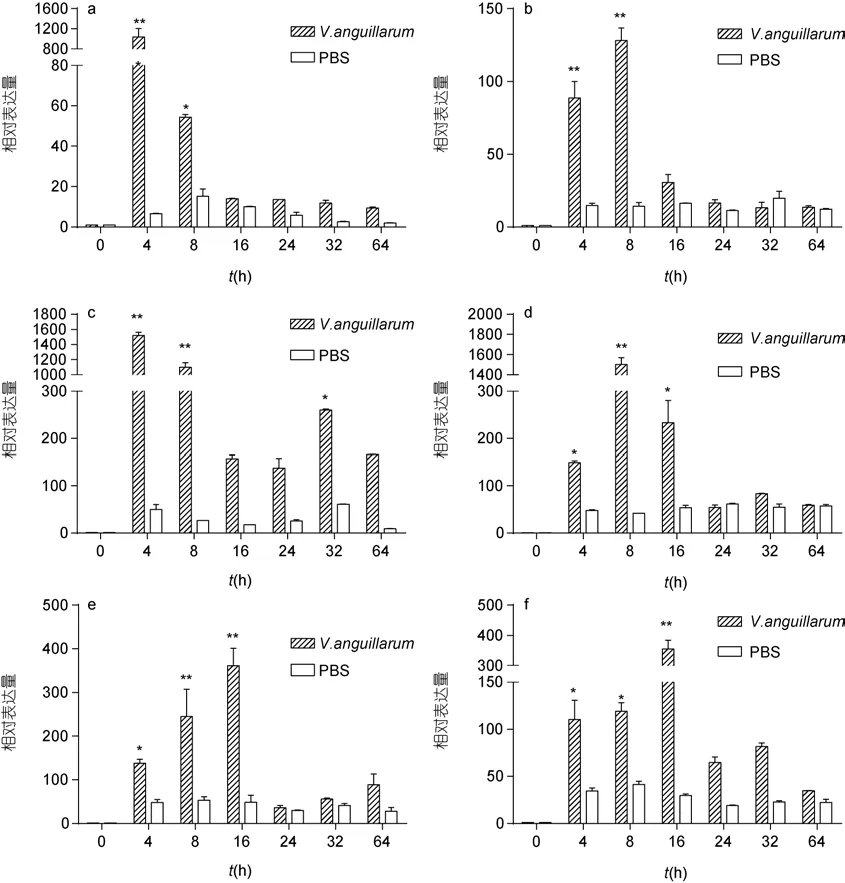
圖5 鰻弧菌感染后魁蚶6種組織SbGal基因的表達(dá)情況Fig.5 SbGal mRNA expression level after V. anguillarum challenge in five tissuesa. 閉殼肌; b. 斧足; c. 肝胰腺; d. 鰓; e. 外套膜; f. 血細(xì)胞。與0h相比, 兩個(gè)星號(hào)表示組間差異極顯著(P<0.01), 一個(gè)星號(hào)表示組間差異顯著(P<0.05)
田吉騰, 2012. 魁蚶微衛(wèi)星富集文庫的構(gòu)建及四個(gè)地理群體的遺傳多樣性分析. 上海: 上海海洋大學(xué)碩士學(xué)位論文,1—59
孫虎山, 李光友, 2001. 雙殼貝類參與免疫防御的體液因子.海洋科學(xué), 25(4): 34—36
吳 彪, 楊愛國, 劉志鴻等, 2010. 魁蚶兩個(gè)不同群體形態(tài)性狀對(duì)體質(zhì)量的影響效果分析. 漁業(yè)科學(xué)進(jìn)展, 31(6): 54—59
吳 彪, 梁 超, 楊愛國等, 2012. 基于 SSR 標(biāo)記的魁蚶(Scapharca broughtonii)不同群體遺傳結(jié)構(gòu)的分析. 海洋與湖沼, 43(4): 863—869
宋小燕, 2009. 海灣扇貝凝集素基因和櫛孔扇貝親環(huán)素A基因的克隆與表達(dá). 咸陽: 西北農(nóng)林科技大學(xué)碩士學(xué)位論文,1—84
陳 騁, 2013. 半滑舌鰨(Cynoglossus semilaevis)免疫相關(guān)基因CsG3BP的克隆、表達(dá)及功能研究. 北京: 中國科學(xué)院研究生院(海洋研究所)碩士學(xué)位論文, 1—72
周麗青, 楊愛國, 王清印等, 2012. 魁蚶四個(gè)地理群體 ITS序列變異及系統(tǒng)發(fā)生分析. 漁業(yè)科學(xué)進(jìn)展, 33(5): 79—84
周麗青, 楊愛國, 王清印等, 2013. 魁蚶血細(xì)胞分類及其免疫功能的初步分析. 水產(chǎn)學(xué)報(bào), 37(4): 599—606
周麗青, 楊愛國, 王清印等, 2014. 鰻弧菌對(duì)魁蚶血細(xì)胞形態(tài)及免疫功能的影響. 海洋與湖沼, 45(3): 536—541
鄭明剛, 2007. 皺紋盤鮑表達(dá)序列標(biāo)簽及免疫相關(guān)基因的克隆與表達(dá)研究. 青島: 中國海洋大學(xué)博士學(xué)位論文, 1—124
禹艷紅, 2007. 文昌魚凝集素基因家族和其他補(bǔ)體相關(guān)的模式識(shí)別分子的克隆表達(dá)、進(jìn)化分析稚功能研究. 廣州: 中山大學(xué)博士學(xué)位論文, 1—192
胥 煒, 王 昊, 宋林生等, 2005. 櫛孔扇貝 C型凝集素基因的克隆與表達(dá)研究. 高技術(shù)通訊, 15(1): 83—88
賀雪明, 唐 麗, 賀福初, 2012. 半乳糖凝集素1的免疫功能.生物技術(shù)通訊, 23(6): 896—899
梁 超, 楊愛國, 劉志鴻等, 2011. 4個(gè)地理群體魁蚶(Scapharca broughtonii)的形態(tài)差異與判別分析. 海洋科學(xué), 35(11): 108—113
董迎輝, 2012. 泥蚶高通量轉(zhuǎn)錄組分析及生長相關(guān)基因的克隆與表達(dá)研究. 青島: 中國海洋大學(xué)博士學(xué)位論文, 1—144
翟玉梅, 丁秀云, 李光友, 1998. 軟體動(dòng)物血細(xì)胞及體液免疫研究進(jìn)展. 海洋與湖沼, 29(5): 558—562
Almkvist J, Karlsson A, 2002. Galectins as inflammatory mediators. Glycoconj J, 19(7—9): 575—581
Barondes S H, 1984. Soluble lectins: a new class of extracellular proteins. Science, 223(4642): 1259—1264
Colnot C, Ripoche M A, Scaerou Fet al, 1996. Galectins in mouse embryogenesis. Biochem Soc Trans, 24(1): 141—146
Drickamer K, 1985. Two distinct classes of carbohydraterecognition domains in animal lectins. J Biol Chem, 263(20):9557—9560
Dumic J, Dabelic S, Fl?gel M, 2006. Galectin-3: an open-ended story. Biochem Biophys Acta, 1760(4): 616—635
Kasai K-I, Hirabayashi J, 1996. Galectins: a family of animal lectins that decipher glycocodes. J Biochem, 119(1): 1—8
Kim J Y, Kim Y M, Cho S Ket al, 2008. Noble tandem-repeat galectin of Manila clamRuditapes philippinarumis induced upon infection with the protozoan parasitePerkinsus olseni.Dev Comp Immunol, 32(10): 1131—1141
Kong P F, Zhang H, Wang L Let al, 2010. AiC1qDC-1, a novel gC1q-domain-containing protein from bay scallopArgopecten irradianswith fungi agglutinating activity. Dev Comp Immunol, 34(8): 834—846
Leffler H, 2001. Galectins structure and function——A synopsis.In:Crocker P R ed. Mammalian Carbohydrate Recognition System. Berlin: Springer, 57—83
Li M, Zhu L, Zhou C Yet al, 2012. Molecular characterization and expression of a novel big defensin (Sb-BDef1) from ark shell,Scapharca broughtonii. Fish Shellfish Immunol, 33(5):1167—1173
Livak K J, Schmittgen T D, 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods, 25(4): 402—408
Nakamura O, Watanabe M, Ogawa Tet al, 2012. Galectins in the abdominal cavity of the conger eelConger myriasterparticipate in the cellular encapsulation of parasitic nematodes by host cells. Fish Shellfish Immunol, 33(4):780—787
Prokhorova E E, Tsymbalenko N V, Ataev G L, 2010. Expression of genes encoding defense factors in the snailPlanorbarius corneus(Gastropoda, Pulmonata) infested with trematodes.Parazitologiia, 44(4): 310—325
Rajan B, Kiron V, Femandes J Met al, 2013. Localization and functional properties of two galectin-1 proteins in Atlantic cod (Gadus morhua) mucosal tissues. Dev Comp Immunol,40(2): 83—93
Song X Y, Zhang H, Wang L Let al, 2011. A galectin with quadruple-domain from bay scallopArgopecten irradiansis involved in innate immune response. Dev Comp Immunol,35(5): 592—602
Song X Y, Zhang H, Zhao J Met al, 2010. An immune responsive multidomain galectin from bay scallopArgopecten irradians. Fish Shellfish Immunol, 28(2):326—332
Tasumi S, Vasta G R, 2007. A galectin of unique domain organization from hemocytes of the Eastern oyster(Crassostrea virginica) is a receptor for the protistan parasitePerkinsus marinus. J Immunol, 179(5): 3086—3098
Vasta G R, 2009. Roles of galectins in infection. Nat Rev Microbiol, 7(6): 424—438
Weiss I M, Kaufmann S, Mann Ket al, 2000. Purification and characterization of perlucin and perlustrin, two new proteins from the shell of the molluskHaliotis laevigata. Biochem Biophys Res Commun, 267(1): 17—21
Zhang D C, Jiang S G, Hu Y Tet al, 2011. A multidomain galectin involved in innate immune responseof pearl oysterPinctada fucata. Dev Comp Immunol, 35(1): 1—6
Zhao J M, Qiu L H, Ning X Xet al, 2010. Cloning and characterization of an invertebrate type lysozyme fromVenerupis philippinarum. Comp Biochem Physiol B Biochem Mol Biol, 156(1): 56—60
