舟山近海日本囊對(duì)蝦野生群體與越冬養(yǎng)成群體形態(tài)性狀對(duì)體重和肉重影響的比較*
2015-03-22 00:59:21李鴻鵬任夙藝王志錚林祖好
海洋與湖沼 2015年5期
李鴻鵬 富 裕 任夙藝 王志錚① 林祖好
(1. 浙江海洋學(xué)院 舟山 316022; 2. 岱山沙城養(yǎng)殖場 岱山 316200)
日本囊對(duì)蝦(Marsupenaeus japonicus)又名日本對(duì)蝦, 俗稱“竹節(jié)蝦”、“花蝦”等, 隸屬于節(jié)肢動(dòng)物門(Arthropoda)、甲殼綱(Crustacea)、十足目(Decapoda)、對(duì)蝦科(Penaeidae), 系廣布于我國東南沿海的暖水性蝦類。日本囊對(duì)蝦體色艷麗、肉鮮味美、營養(yǎng)經(jīng)濟(jì)價(jià)值高, 且具生長迅速、抗逆性強(qiáng)、適宜活蝦長途干運(yùn)等特點(diǎn), 我國自20世紀(jì)80年代中期開展全人工養(yǎng)殖以來, 養(yǎng)殖地域逐漸拓展, 產(chǎn)業(yè)規(guī)模不斷擴(kuò)大, 現(xiàn)已與凡納濱對(duì)蝦(Litopenaeus vannamei)、中國明對(duì)蝦(Fenneropenaeus chinensis)和斑節(jié)對(duì)蝦(Penaeus monodon)一起成為當(dāng)前我國四大主養(yǎng)海水蝦類。據(jù)統(tǒng)計(jì), 目前我國日本囊對(duì)蝦養(yǎng)殖面積達(dá)1.8925×104hm2,年養(yǎng)殖產(chǎn)量為4.5949×104t (農(nóng)業(yè)部漁業(yè)局, 2014)。
據(jù)報(bào)道, 日本囊對(duì)蝦不同地理居群間以及野生群體與養(yǎng)殖群體間在形態(tài)特征上均存在較為明顯的差異(李義軍等, 2010; 蔡曉鵬等, 2010; 董宏標(biāo)等,2014); Tsio等(2005, 2007)按頭胸甲側(cè)面斜紋延伸特征將日本囊對(duì)蝦分型為在地理緯度分布上具較為明顯差異的形態(tài)變異Ⅰ型(varietyⅠ, 斜紋延伸至頭部底緣)和Ⅱ型(varietyⅡ, 斜紋僅延伸至頭胸甲側(cè)面中央), 其中前者主要分布于緯度相對(duì)較高的東亞海域,而后者則主要分布于緯度相對(duì)較低的南海海域。無疑,上述關(guān)于日本囊對(duì)蝦因所處不同生存環(huán)境而導(dǎo)致其外部形態(tài)表露差異的情形, 表明日本囊對(duì)蝦部分形態(tài)性狀具較強(qiáng)的環(huán)境可塑性。研究發(fā)現(xiàn), 日本囊對(duì)蝦北部灣野生群體以及 2種形態(tài)變異類型人工養(yǎng)殖群體的形態(tài)表型均與其體重性狀有著極為密切的關(guān)系,且經(jīng)相關(guān)分析、通徑分析和偏回歸分析均可找尋到影響其體重性狀的關(guān)鍵形態(tài)性狀組合, 其中影響北部灣野生群體體重性狀的關(guān)鍵形態(tài)性狀組合為體長、頭胸甲長、胸寬、胸高、第一腹節(jié)寬、第三腹節(jié)高和額劍上刺數(shù)等 7個(gè)變量(孫成波等, 2008), 影響形態(tài)變異Ⅰ型人工養(yǎng)殖群體體重性狀的關(guān)鍵形態(tài)性狀組合為體長、第一腹節(jié)寬、頭胸部寬、第五腹節(jié)寬、第三腹節(jié)寬和第六腹節(jié)寬等6個(gè)變量, 而影響形態(tài)變異Ⅱ型人工養(yǎng)殖群體體重性狀的關(guān)鍵形態(tài)性狀組合則為體長、頭胸部長、第五腹節(jié)寬和第一腹節(jié)寬等4個(gè)變量(董宏標(biāo)等, 2014)。綜上可知, 日本囊對(duì)蝦形態(tài)表型系對(duì)其所處生存環(huán)境及體重增長對(duì)策的綜合響應(yīng)。
舟山海域日本囊對(duì)蝦野生捕撈群體的主要漁獲期為6—9月(毛錫林等, 1994)。受其沖擊, 舟山沿海乃至東海區(qū)日本囊對(duì)蝦春苗和夏苗人工養(yǎng)成群體的市場平均售價(jià)均遠(yuǎn)低于秋苗養(yǎng)成群體; 而秋苗人工養(yǎng)成群體因在露天池塘越冬養(yǎng)成過程中需耐受低溫和高pH值的雙重脅迫(富裕等, 2014), 致使養(yǎng)成產(chǎn)量遠(yuǎn)低于春苗和夏苗人工養(yǎng)成群體, 故日本囊對(duì)蝦秋苗親體的選育工作就顯得十分重要。鑒于此, 本文作者以舟山近海可作為日本囊對(duì)蝦秋苗親體選育的夏季海捕野生群體和經(jīng)越冬養(yǎng)成的秋苗同生群為研究對(duì)象, 以體重和肉重為目標(biāo)變量, 采取多元分析方法定量比較兩者間影響體重和肉重關(guān)鍵形態(tài)性狀組合的差異, 并據(jù)此分析引起兩者在體重和肉重增長機(jī)制上產(chǎn)生差異的原因, 旨為提高該蝦秋苗親體的選種精度與效率提供基礎(chǔ)資料。
1 材料與方法
1.1 樣品來源
本研究所用露天池塘日本囊對(duì)蝦秋苗越冬養(yǎng)成群體(以下簡稱“越冬養(yǎng)成群體”)實(shí)驗(yàn)蝦取自岱山沙城養(yǎng)殖場內(nèi)面積為4.3×666.7 m2的日本囊對(duì)蝦秋苗與毛蚶混養(yǎng)池塘, 系 8月齡秋苗同生群養(yǎng)殖個(gè)體(蝦苗體長0.8cm, 放養(yǎng)時(shí)間為2013年9月27日, 起捕時(shí)間為2014年4月29日), 所用日本囊對(duì)蝦海捕野生群體(以下簡稱“海捕群體”)實(shí)驗(yàn)蝦于2014年8月11日捕自舟山市岱山縣長涂島鄰近海域。
1.2 表型性狀測定
分別于所采越冬養(yǎng)成群體和海捕群體中隨機(jī)選取肢體完整、無病無傷、反應(yīng)敏捷的健壯個(gè)體各150尾作為實(shí)驗(yàn)測定群體。樣品逐一編號(hào)后, 用數(shù)顯游標(biāo)卡尺(精度0.02 mm)逐尾測量體長(X1)、頭胸甲長(X2)、胸寬(X3)、胸高(X4)、第二腹節(jié)長(X5)、第二腹節(jié)寬(X6)、第二腹節(jié)高(X7)、尾節(jié)長(X8)、尾節(jié)寬(X9)、尾節(jié)高(X10)、眼徑(X11)、眼窩距(X12)、額劍長(X13)并計(jì)數(shù)額劍上緣鋸齒數(shù)(X14), 用上海恒平科學(xué)儀器有限公司出品的RS-232型精密電子天平(精度0.01g)稱量經(jīng)濾紙吸凈體表水分后的體重(Y1)和經(jīng)蒸煮、去頭、去殼并瀝干表面水分后的肉重(Y2)。
1.3 分析方法
整理所測結(jié)果, 運(yùn)用SPSS 17.0軟件依次統(tǒng)計(jì)各表型性狀和體型比例性狀(以體長為基準(zhǔn))的均值、標(biāo)準(zhǔn)差和變異系數(shù); 經(jīng)表型相關(guān)分析和各形態(tài)性狀對(duì)重量性狀(Y1,Y2)的通徑分析, 剔除那些通徑系數(shù)未達(dá)到顯著水平的形態(tài)性狀(P>0.05)后, 按相關(guān)系數(shù)組成效應(yīng), 將那些被保留下來的形態(tài)性狀與重量性狀的相關(guān)系數(shù)(rij)剖分為直接作用(通徑系數(shù)P1)和通過其它性狀的間接作用(P2)兩部分, 進(jìn)而計(jì)算決定系數(shù)和復(fù)相關(guān)指數(shù), 并據(jù)此確定影響重量性狀的關(guān)鍵形態(tài)性狀組合; 通過偏回歸分析, 保留偏回歸系數(shù)達(dá)到顯著水平(P<0.05)的形態(tài)性狀, 分別建立這些被保留的形態(tài)性狀與體重和肉重間的多元回歸方程, 并進(jìn)行擬合度檢驗(yàn)。
2 結(jié)果與分析
2.1 表型性狀參數(shù)估計(jì)值
由表1和表2可見, 越冬養(yǎng)成群體和海捕群體在本研究所涉生物學(xué)測定指標(biāo)和體型比例指標(biāo)(以體長為基準(zhǔn))上的異同主要表露為: (1) 越冬養(yǎng)成群體雖在所測16項(xiàng)生物學(xué)指標(biāo)中, 僅X14測量值顯著小于海捕群體(P<0.05), 但其與海捕群體的變異系數(shù)均仍以重量性狀(Y1,Y2)為最大且遠(yuǎn)大于其它形態(tài)性狀, 表明日本囊對(duì)蝦重量性狀較形態(tài)性狀更具選育潛力; (2) 在所涉 12項(xiàng)體型比例指標(biāo)中, 越冬養(yǎng)成群體與海捕群體間無顯著差異的僅為C1、C5和C6(P>0.05), 即兩者體型相似性指數(shù)為 25%, 其中越冬養(yǎng)成群體較海捕群體表露出眼徑大, 胸圍寬大, 尾節(jié)粗短, 以及額劍、第二腹節(jié)和眼窩距均較短的體型特征, 表明日本囊對(duì)蝦體型具較強(qiáng)的環(huán)境可塑性。
2.2 體重和肉重與形態(tài)性狀間的相關(guān)性
由表3、表4可知, 越冬養(yǎng)成群體和海捕群體在本研究所涉重量性狀與形態(tài)性狀間相關(guān)性的異同上主要表現(xiàn)為: (1) 本研究兩測定群體的重量性狀均與除X14外的其它形態(tài)性狀呈極顯著相關(guān)(P<0.01);(2) 越冬養(yǎng)成群體Y1和Y2兩重量性狀與形態(tài)性狀間的相關(guān)系數(shù)大小排序基本一致, 而海捕群體Y1和Y2與形態(tài)性狀間的相關(guān)系數(shù)大小排序則不僅具較為明顯的差異, 且均不同于對(duì)應(yīng)的越冬養(yǎng)成群體, 即越冬養(yǎng)成群體形態(tài)性狀在對(duì)Y1和Y2的影響上較海捕群更具一致性, 且兩者形態(tài)性狀在對(duì)Y1和Y2的響應(yīng)特征上也存在較明顯的差異, 表明本研究所選形態(tài)性狀可綜合反映日本囊對(duì)蝦在所處生存環(huán)境及Y1、Y2增長特征上的差異。

表1 日本囊對(duì)蝦實(shí)驗(yàn)測定群體表型性狀的統(tǒng)計(jì)量描述Tab.1 The statistics of phenotypic traits of tested groups
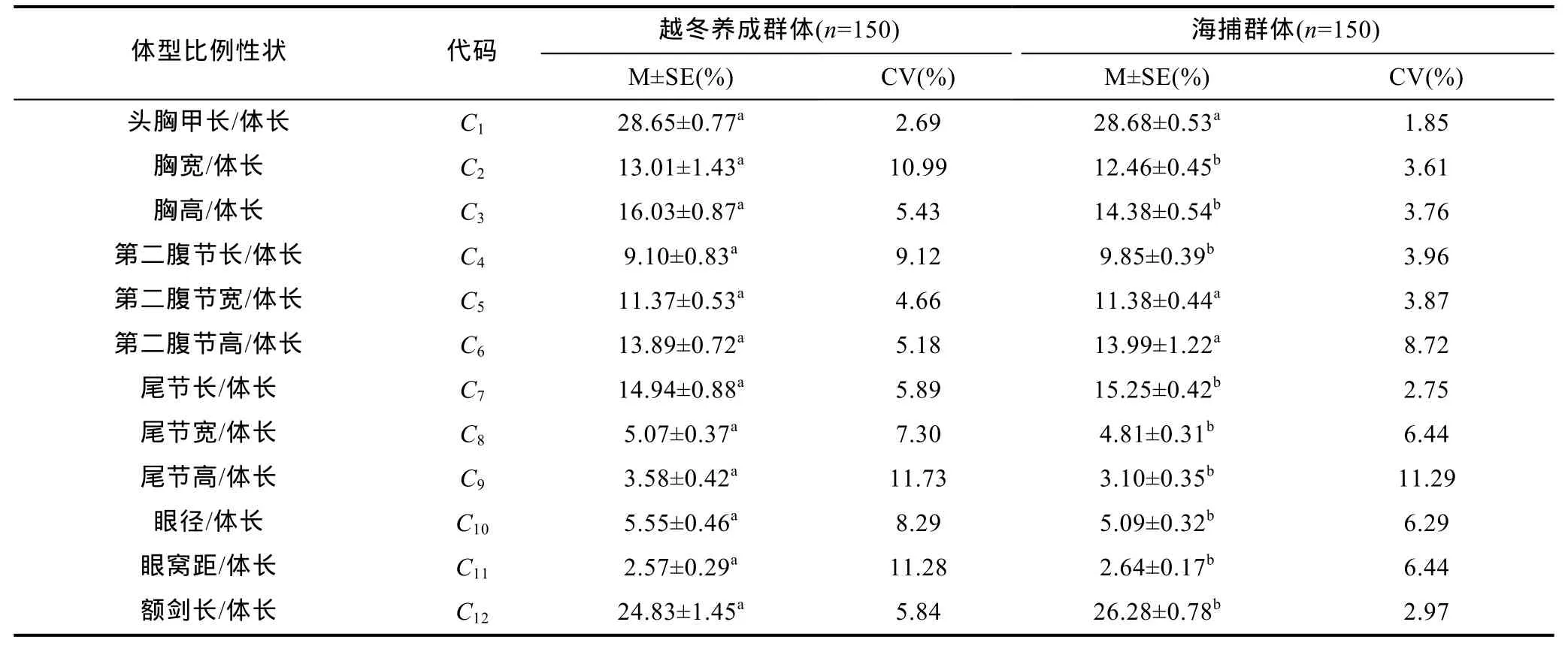
表2 日本囊對(duì)蝦實(shí)驗(yàn)測定群體體型比例性狀的統(tǒng)計(jì)量描述Tab.2 The statistics of characteristics of somatotype proportion of tested groups
2.3 形態(tài)性狀對(duì)體重和肉重的通徑分析
由表5可見, 在越冬養(yǎng)成群體被保留的形態(tài)性狀中, 對(duì)Y1和Y2的直接作用大于間接作用的性狀均僅為X1, 且均以X1對(duì)Y1和Y2的直接作用為最大(分別為0.541和0.820), 而其余被保留的形態(tài)性狀對(duì)Y1和Y2的間接作用則均主要通過X1來實(shí)現(xiàn),X2對(duì)Y1及X6對(duì)Y2的直接作用均僅弱于X1, 且X1對(duì)Y1、Y2的間接
作用分別主要通過X2和X6來實(shí)現(xiàn), 表明X1為影響越冬養(yǎng)成群體Y1、Y2的核心變量,X2、X6分別為影響越冬養(yǎng)成群體Y1、Y2的重要變量, 而其余被保留的形態(tài)性狀則均為影響越冬養(yǎng)成群體Y1、Y2的相對(duì)次要變量。經(jīng)計(jì)算, 上述被保留的形態(tài)性狀組合對(duì)Y1、Y2的相關(guān)指數(shù)(R2=∑Pirxiy,Pi為通徑系數(shù),rxiy為形態(tài)性狀與重量性狀的相關(guān)系數(shù))分別為0.955和0.907。

表3 日本囊對(duì)蝦越冬養(yǎng)成群體各表型性狀間的相關(guān)系數(shù)(df=148)Tab.3 The correlation coefficient among the phenotypic traits of overwintered cultivated group
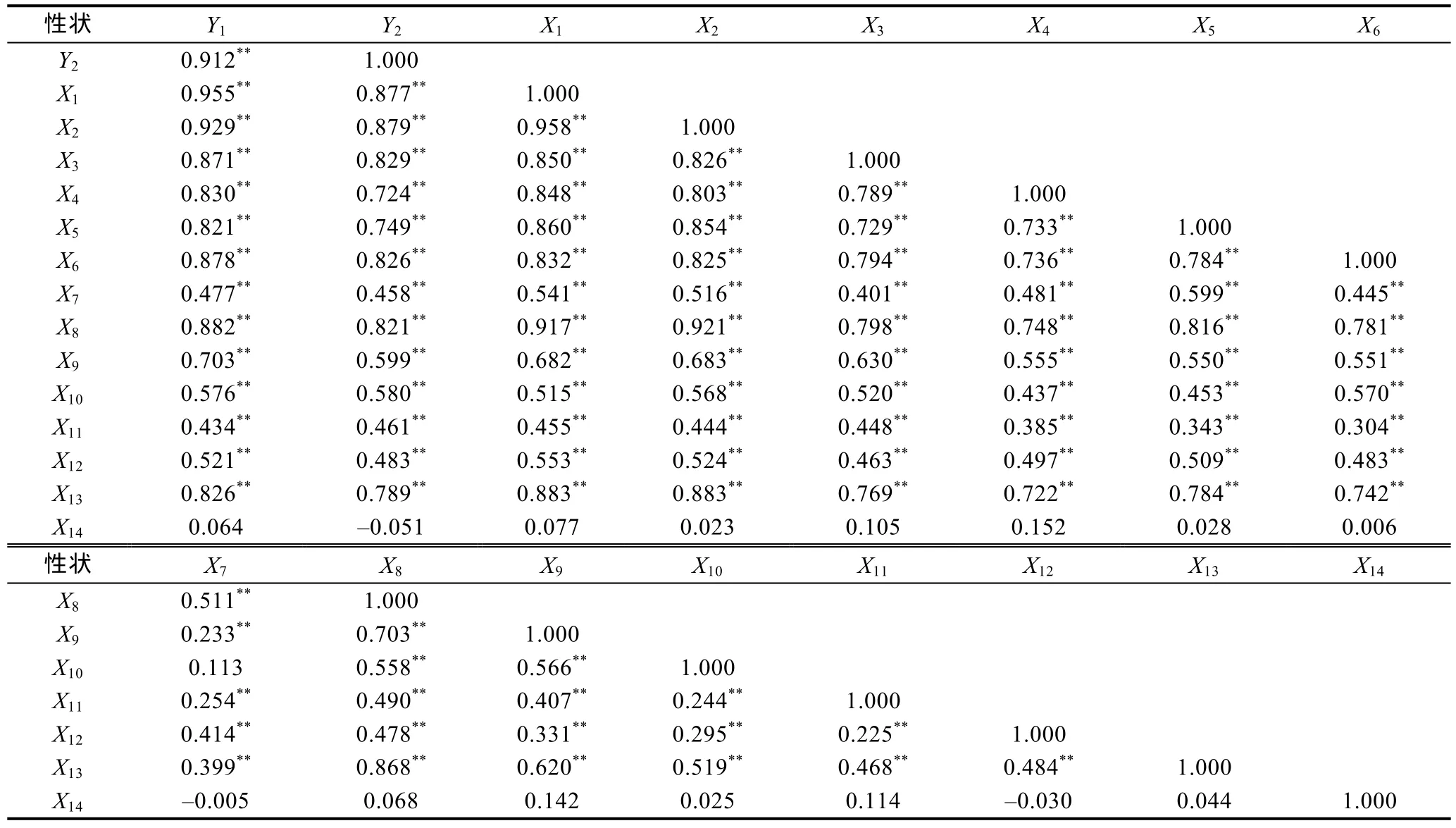
表4 日本囊對(duì)蝦海捕群體各表型性狀間的相關(guān)系數(shù)(df=148)Tab.4 The correlation coefficient among the phenotypic traits of wild group
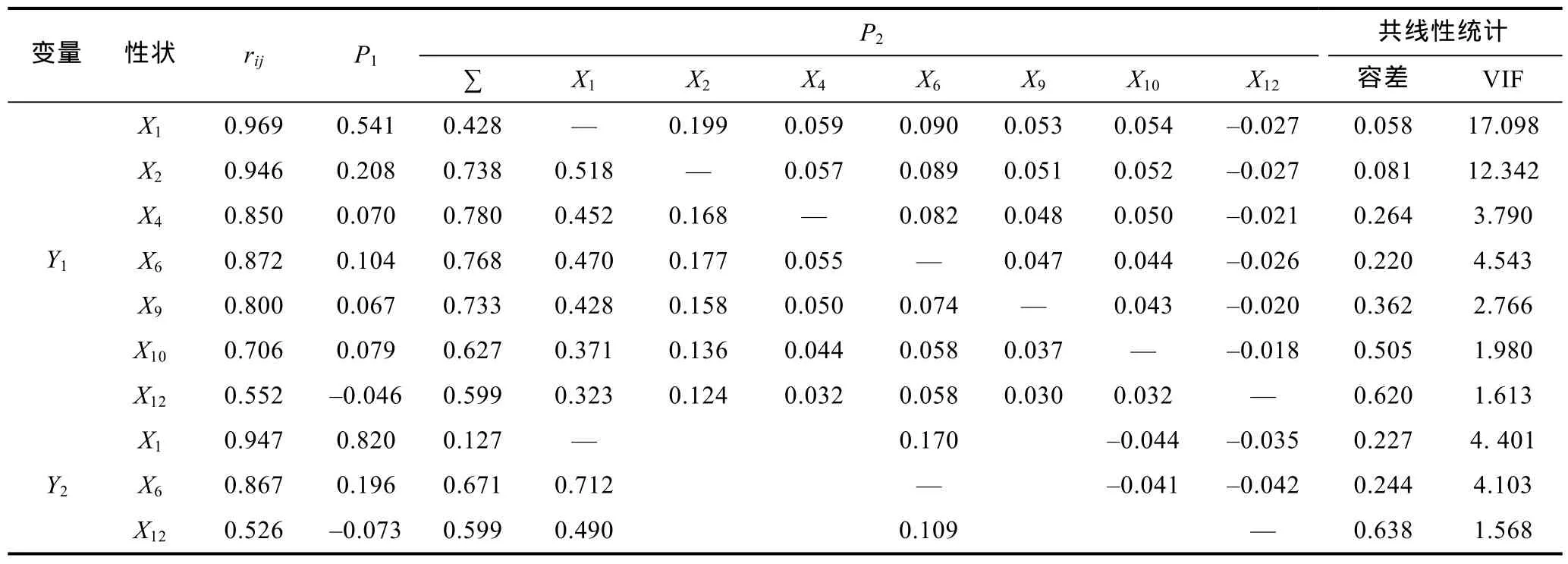
表5 日本囊對(duì)蝦形態(tài)性狀對(duì)越冬養(yǎng)成群體體重和肉重影響的通徑分析Tab.5 The path analysis of morphological characters on body weight and meat weight for overwintered cultivated group
由表6可見, 海捕群體被保留的形態(tài)性狀中對(duì)Y1和Y2的直接作用大于間接作用的性狀分別僅為X1和X2, 且分別以X1對(duì)Y1和X2對(duì)Y2的直接作用為最大(分別為0.671和0.440), 而其余被保留的形態(tài)性狀對(duì)Y1和Y2的間接作用也均分別主要通過X1和X2來實(shí)現(xiàn),X6對(duì)Y1和Y2的直接作用分別僅弱于X1和X2, 且X1對(duì)Y1的間接作用和X2對(duì)Y2的間接作用均主要通過X6來實(shí)現(xiàn), 表明X1、X2為分別影響海捕群體Y1和Y2的核心變量,X6為影響海捕群體Y1、Y2的重要變量,而其余被保留的形態(tài)性狀則均為影響海捕群體Y1、Y2的相對(duì)次要變量。經(jīng)計(jì)算, 上述被保留的形態(tài)性狀組合對(duì)Y1、Y2的相關(guān)指數(shù)分別為0.944和0.837。
2.4 形態(tài)性狀對(duì)體重和肉重的決定程度分析
據(jù)表3、表4、表5及表6所列數(shù)據(jù), 分別計(jì)算單個(gè)性狀對(duì)Y1和Y2的決定系數(shù)(di=Pi2,Pi為性狀對(duì)Y1或Y2的通徑系數(shù))以及性狀兩兩交互對(duì)Y1和Y2的共同決定系數(shù)(dij=2rijPiPj,rij為兩性狀間的相關(guān)系數(shù),Pi、Pj分別為兩性狀對(duì)Y1或Y2的通徑系數(shù)), 并列結(jié)果于表7和表8。
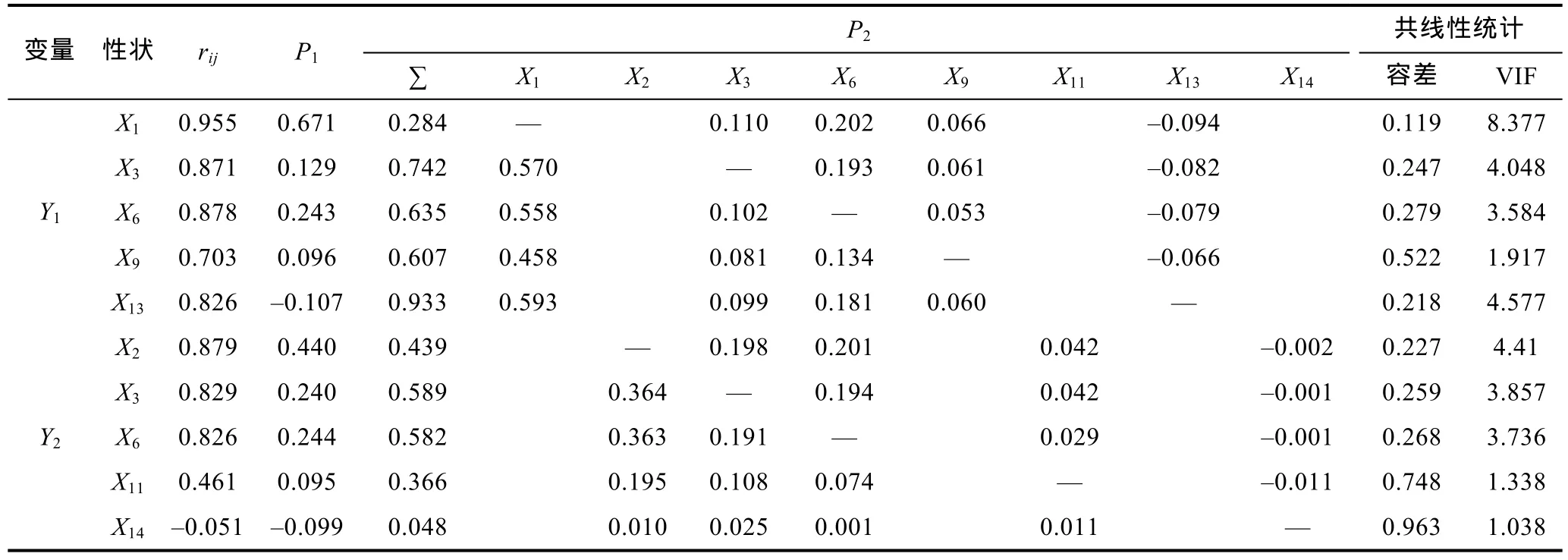
表6 日本囊對(duì)蝦形態(tài)性狀對(duì)海捕群體體重和肉重影響的通徑分析Tab.6 The path analysis of morphological characters on body weight and meat weight of wild group
由表7和表8可見, 越冬養(yǎng)成群體X1對(duì)Y1和Y2的相對(duì)決定程度(分別為 29.3%和 67.2%)均遠(yuǎn)高于被保留的其它形態(tài)性狀, 兩性狀交互對(duì)Y1和Y2的共同決定系數(shù)均分別以X1—X2、X1—X6組合(依次為0.215和0.279)為最大; 海捕群體X1對(duì)Y1以及X2對(duì)Y2的相對(duì)決定程度(分別為 45.3%和 19.4%)均遠(yuǎn)高于被保留的其它形態(tài)性狀, 兩性狀交互對(duì)Y1和Y2的共同決定系數(shù)均分別以X1—X6、X2—X6組合(依次為 0.271和0.177)為最大。上述結(jié)果, 無疑進(jìn)一步印證了表 5和表6所揭示的X1為影響越冬養(yǎng)成群體Y1、Y2的核心變量,X2、X6分別為影響越冬養(yǎng)成群體Y1、Y2的重要變量, 以及X1、X2為分別影響海捕群體Y1和Y2的核心變量,X6為影響海捕野生群體Y1、Y2的重要變量的可靠性。經(jīng)統(tǒng)計(jì), 表7、表8中所列形態(tài)性狀組合對(duì)越冬養(yǎng)成群體和海捕群體Y1、Y2決定系數(shù)的加和值分別為0.955、0.907和0.944、0.837, 均等于其對(duì)應(yīng)的相關(guān)指數(shù)R2值(表9)。
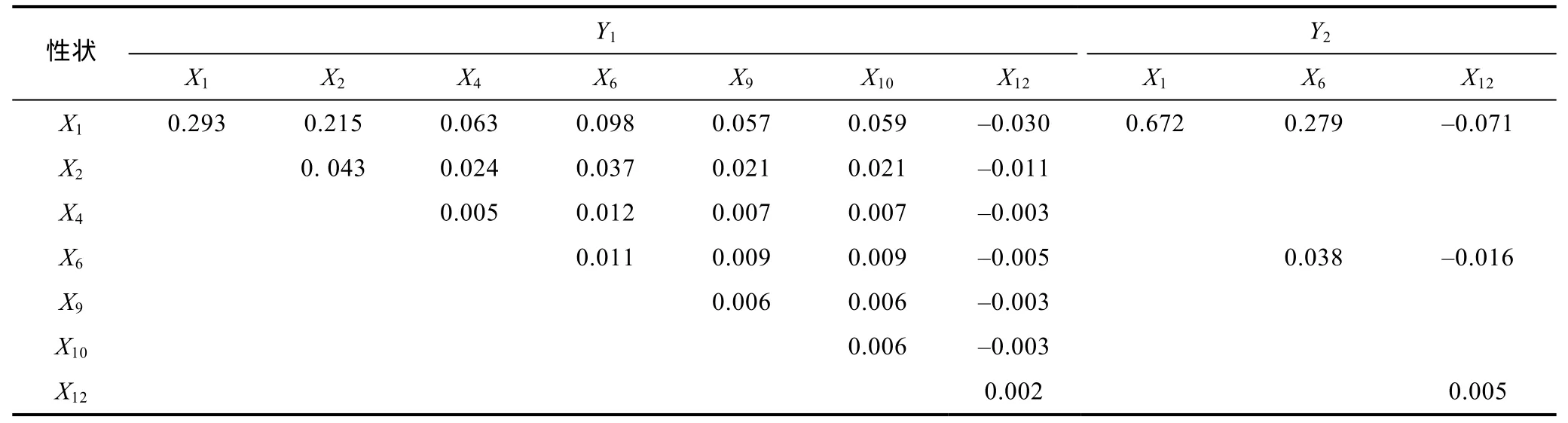
表7 日本囊對(duì)蝦越冬養(yǎng)成群體形態(tài)性狀對(duì)體重和肉重的決定系數(shù)Tab.7 The determination coefficient of morphological characters on body weight and meat weight of overwintered cultivated group
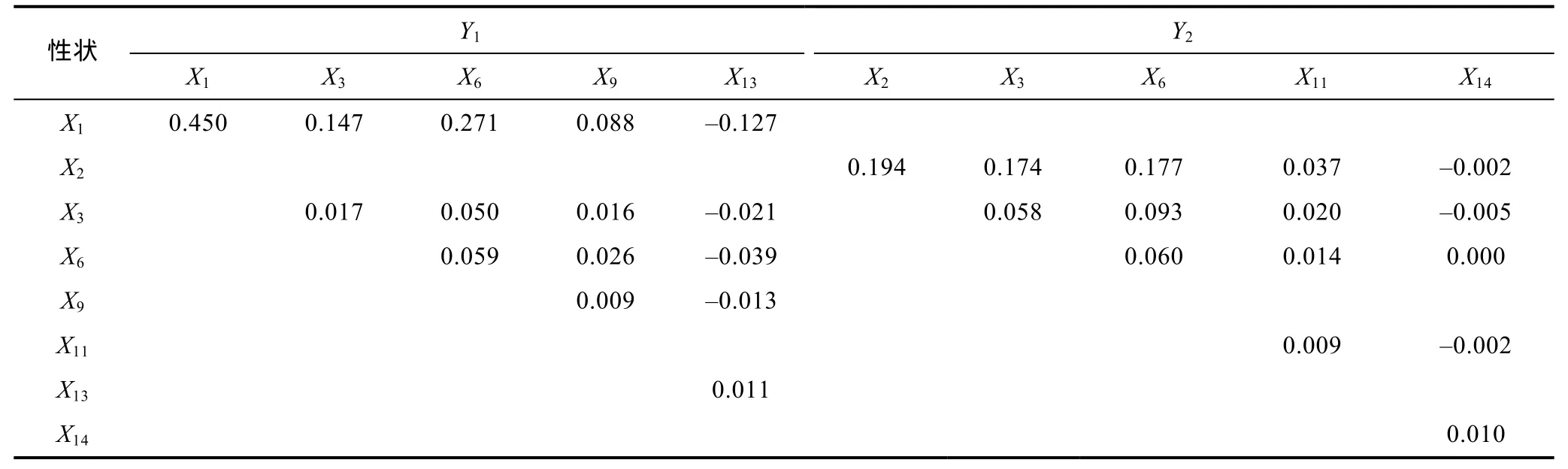
表8 日本囊對(duì)蝦海捕群體形態(tài)性狀對(duì)體重和肉重的決定系數(shù)Tab.8 The determination coefficient of morphological characters on body weight and meat weight of wild group
2.5 形態(tài)性狀與體重和肉重間多元回歸方程的建立
對(duì)經(jīng)通徑分析被保留的形態(tài)性狀進(jìn)行復(fù)相關(guān)分析和回歸分析, 得表9、表10和表11。由表9可見, 越冬養(yǎng)成群體和海捕群體中那些被保留的形態(tài)性狀組合與其所對(duì)應(yīng)的Y1和Y2間的復(fù)相關(guān)系數(shù)均達(dá)到極顯著水平(R>r0.01), 表明它們與Y1和Y2間具極為密切的內(nèi)在聯(lián)系, 在對(duì)Y1和Y2的影響上較其它未被保留的形態(tài)性狀更具重要性。

表9 日本囊對(duì)蝦形態(tài)性狀與體重、肉重間的復(fù)相關(guān)分析Tab.9 The multiple correlation analysis between morphological characters, and body weight and meat weight of M. japonicus
由表 10可見, 用于估算越冬養(yǎng)成群體體重和肉重的回歸方程分別為Y1=0.180X1+0.228X2+0.129X4+0.287X6+0.330X9+0.412X10–0.371X12–20.677 和Y2=0.127X1+0.253X6–0.276X12–9.972, 用于估算海捕群體體重和肉重的回歸方程分別為Y1=0.152X1+0.217X3+0.439X6+0.327X9–0.100X13–12.456 和Y2=0.129X2+0.155X3+0.17X6+0.184X11–0.078X14–4.267, 且上述方程的回歸截距及
所涉形態(tài)性狀的偏回歸系數(shù)均達(dá)到顯著水平(P<0.05);經(jīng)方差分析和回歸預(yù)測(表11), 所建各回歸方程的回歸系數(shù)均達(dá)到極顯著水平(P<0.01), 且估計(jì)值與實(shí)測值間也均無顯著差異(P>0.05)。表明, 本研究所建上述回歸方程可精確反映越冬養(yǎng)成群體和海捕群體形態(tài)性狀與Y1和Y2間的真實(shí)關(guān)系。

表10 日本囊對(duì)蝦形態(tài)性狀與體重、肉重間的偏回歸系數(shù)檢驗(yàn)Tab.10 The partial regression coefficient between morphological characters, and body weight and meat weight of M. japonicus
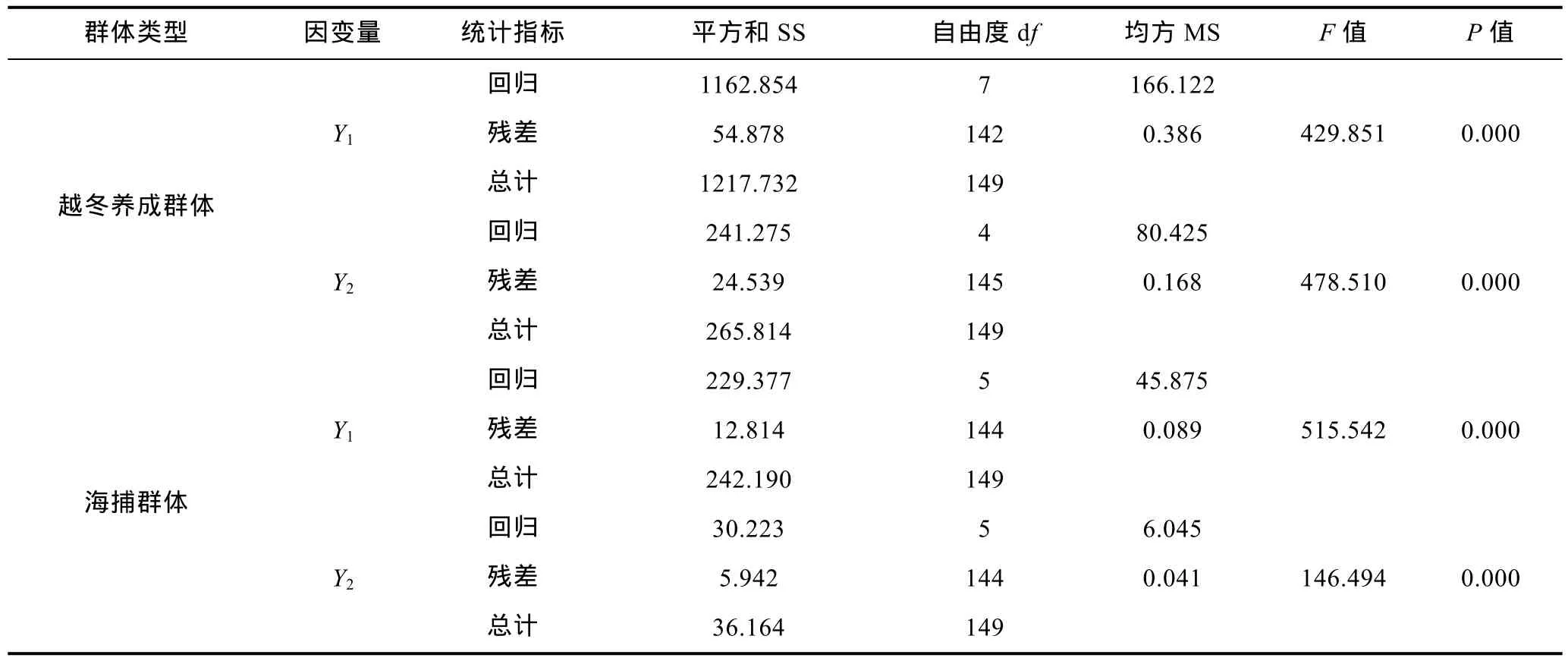
表11 日本囊對(duì)蝦形態(tài)性狀與體重性狀間多元回歸方程的方差分析Tab.11 The variance analysis of multiple regression equation between morphological characters and body weight traits of M.japonicus
3 討論
3.1 日本囊對(duì)蝦形態(tài)表型的生境適應(yīng)性
范志勤(1973)指出, 動(dòng)物為增加或減少散熱面積,其體型大小常隨溫度而變化, 一定的體型適應(yīng)于一定溫度范圍的代謝水平。本研究中, 越冬養(yǎng)成群體較海捕群體呈現(xiàn)出眼徑大、胸圍寬大、尾節(jié)粗短、以及額劍、第二腹節(jié)長和眼窩距均較短的體型特征(表2),既與內(nèi)溫動(dòng)物在氣候寒冷地區(qū)體型趨于增大的情勢(shì)(Bergmann, 1847)相仿, 也與內(nèi)溫動(dòng)物身體突出部分在氣候寒冷地區(qū)有變短趨勢(shì)的情形(Allen, 1877)相似,表明兩者在體型特征上的差異為適應(yīng)各自所處溫度環(huán)境下的生理代謝所致。
據(jù)報(bào)道, 在海南養(yǎng)成的種苗源自福建廈門的日本囊對(duì)蝦人工養(yǎng)殖群體, 在形態(tài)表型上系明顯有別于福建廈門、海南臨高和廣東湛江海域日本囊對(duì)蝦野生種群的又一獨(dú)立種群(李義軍等, 2010)。這一研究結(jié)果與本研究所涉越冬養(yǎng)成群體和海捕群體雖均屬Tsio等(2005, 2007)認(rèn)為的日本囊對(duì)蝦形態(tài)變異Ⅰ型,但兩者在額劍上緣鋸齒數(shù)和以體長為基準(zhǔn)的 9項(xiàng)體型比例性狀中均存在顯著差異(表1, 表2)的情形, 表明生存環(huán)境差異是導(dǎo)致日本囊對(duì)蝦養(yǎng)殖群體在形態(tài)表型上顯著偏離野生群體的主因, 即日本囊對(duì)蝦為適應(yīng)所處生存環(huán)境在形態(tài)表型上具較強(qiáng)的可塑性。另外, 日本囊對(duì)蝦因地域分布不同而分化為形態(tài)變異Ⅰ型和Ⅱ型的兩群體間在主要外形參數(shù)上均具顯著差異(董宏標(biāo)等, 2014); 分布于海南臨高和廣東湛江海域的野生日本囊對(duì)蝦因地理距離較近而被判為同一地理種群, 兩者與地理距離均較遠(yuǎn)的福建廈門海域日本囊對(duì)蝦野生種群在形態(tài)表型上均具顯著差異(李義軍等, 2010); 蔡曉鵬等(2010)對(duì)分布于海南陵水、廣西北海、廣東惠來、福建詔安和福建廈門沿海的 5個(gè)日本囊對(duì)蝦地理群體開展了形態(tài)差異比較分析, 結(jié)果表明同一性別不同地理群體間的形態(tài)分別以陵水和北海群體間較為相近, 詔安和廈門群體間較為相近。無疑, 上述有關(guān)日本囊對(duì)蝦因所處生存環(huán)境迥異而導(dǎo)致其形態(tài)表型產(chǎn)生顯著差異的情形, 進(jìn)一步驗(yàn)證了王志錚等(2012)指出的同一物種不同養(yǎng)殖模式養(yǎng)成群體間形質(zhì)特征差異的本質(zhì), 為它們對(duì)各自特定生境的表型適應(yīng)的觀點(diǎn)的可靠性。故, 鑒于養(yǎng)殖環(huán)境在對(duì)日本囊對(duì)蝦體型塑造上所起的“磨刀石”般的作用, 為盡可能降低越冬養(yǎng)殖過程中因表型適應(yīng)而付出的體能損耗和提高苗種養(yǎng)殖成活率之考量,日本囊對(duì)蝦秋苗親體外觀體型的選擇應(yīng)以秋苗越冬養(yǎng)成群體為參照。
3.2 日本囊對(duì)蝦越冬養(yǎng)成群體和海捕群體間重量性狀增長對(duì)策的差異分析
盡管本研究中在露天養(yǎng)殖池塘內(nèi)經(jīng)歷冬季持續(xù)低溫和高pH值脅迫(富裕等, 2014)而存活下來的日本囊對(duì)蝦越冬養(yǎng)成群體, 與常面臨饑餓脅迫而在夏季長成的日本囊對(duì)蝦海捕群體, 經(jīng)通徑分析被保留的形態(tài)性狀對(duì)體重的相關(guān)指數(shù)均大于劉小林等(2002)所認(rèn)為的已找尋到影響目標(biāo)變量的關(guān)鍵自變量組合的相關(guān)指數(shù)閾值0.85, 且影響本研究所涉日本囊對(duì)蝦越冬養(yǎng)成群體和海捕群體體重的核心表型變量均為X1的情形(表 5, 表 6, 表 7, 表 8, 表 9), 與董宏標(biāo)等(2014)報(bào)道的日本囊對(duì)蝦形態(tài)變異Ⅰ型與Ⅱ型群體均雷同, 但兩者在以體長為基準(zhǔn)的體型比例性狀和影響體重的形態(tài)性狀組成上卻均存在較大差異(表 2,表 5, 表 6), 并與已報(bào)道的北部灣野生日本囊對(duì)蝦以及在同池飼養(yǎng)條件下日本囊對(duì)蝦形態(tài)變異類型Ⅰ和Ⅱ人工養(yǎng)殖群體在影響體重性狀的形態(tài)性狀組成上也均具一定區(qū)別(孫成波等, 2008; 董宏標(biāo)等, 2014)的情形, 無疑反映了日本囊對(duì)蝦體重增長特征與種質(zhì)規(guī)定性和環(huán)境異質(zhì)性共同作用所致的形態(tài)表型可塑性間有著較為密切的關(guān)系, 即日本囊對(duì)蝦的體重增長對(duì)策受種質(zhì)規(guī)定性和環(huán)境異質(zhì)性的雙重影響, 與Wagner等(2001)認(rèn)為同一物種的不同群體因長期適應(yīng)不同的生存環(huán)境而形成了各自特有包括生長、發(fā)育、繁殖以及對(duì)環(huán)境因子的適應(yīng)力等的生物學(xué)特性的觀點(diǎn)相吻。
由表5、表7可見, 影響本研究所涉越冬養(yǎng)成群體肉重的形態(tài)表型變量均包含于影響體重的形態(tài)表型變量組合中, 且影響兩者的核心形態(tài)表型變量均為X1, 而影響體重的形態(tài)變量不僅較肉重增加了X2、X4、X9和X10, 且其重要表型變量為X2而非肉重的X6,表明越冬養(yǎng)成群體的肉重增長雖隱含于其體重增長對(duì)策之中, 但體重增長較肉重增長在更側(cè)重于包含血循、呼吸和消化器官的頭胸部增重的同時(shí), 也兼顧到了在保持蝦體運(yùn)動(dòng)平衡和遇敵急速逃逸方面具重要功能的尾部增重, 這無疑與其在越冬養(yǎng)殖過程中需經(jīng)受持續(xù)低溫和高pH值的雙重脅迫及較高強(qiáng)度的種內(nèi)競食有關(guān)。與越冬養(yǎng)成群體相較, 影響本研究所涉海捕群體體重和肉重的核心表型變量分別為X1和X2, 而重要表型變量則均為X6的情形(表6, 表8), 表明其體重增長較越冬養(yǎng)成群體更側(cè)重于通過腹部肌肉的增重以提升其游泳能力, 而其肉重的增長較越冬養(yǎng)成群體則更注重于通過提高血循、呼吸和消化效率來實(shí)現(xiàn)。無疑, 海捕群體食物的相對(duì)匱乏以及越冬養(yǎng)成群體所歷養(yǎng)殖理化環(huán)境和種內(nèi)斗爭的高嚴(yán)峻度是導(dǎo)致兩者間體重和肉重增長對(duì)策產(chǎn)生顯著差異的主因。
綜上, 鑒于本研究所涉越冬養(yǎng)成群體基于形態(tài)表型的肉重增長對(duì)策已隱含于體重增長對(duì)策中, 及海捕群體經(jīng)通徑分析被保留的形態(tài)性狀對(duì)肉重的相關(guān)指數(shù)未達(dá)到劉小林等(2002)所認(rèn)為的已找尋到影響目標(biāo)變量的關(guān)鍵自變量組合的相關(guān)指數(shù)閾值之考量,本文作者認(rèn)為從舟山沿海夏季海捕野生群體中遴選以體重為目標(biāo)選育性狀的日本囊對(duì)蝦秋苗親體時(shí),為提高選擇精度, 宜取Y1=0.152X1+0.217X3+0.439X6+0.327X9–0.100X13–12.456 和Y1=0.180X1+0.228X2+0.129X4+0.287X6+0.330X9+0.412X10–0.371X12–20.677 兩值相近者作為秋苗親體選擇的基本群, 以使其外觀體型及基于表型的體重增長對(duì)策均與秋苗越冬養(yǎng)成群體基本相吻。令上述兩式相等, 整理得 0.018X1+0.228X2–0.217X3+0.129X4–0.152X6+0.003X9+0.412X10–0.371X12+0.100X13–8.221=0, 該式為篩選日本囊對(duì)蝦秋苗親本的形態(tài)評(píng)估模型。
王志錚, 楊 磊, 朱衛(wèi)東, 2012. 三種養(yǎng)殖模式下日本鰻鱺養(yǎng)成品的形質(zhì)差異. 應(yīng)用生態(tài)學(xué)報(bào), 23(5): 1385—1392
毛錫林, 蔣文波, 1994. 舟山海域海洋生物志. 杭州: 浙江人民出版社, 114
劉小林, 常亞青, 相建海等, 2002. 櫛孔扇貝殼尺寸性狀對(duì)活體重的影響效果分析. 海洋與湖沼, 33(6): 673—678
農(nóng)業(yè)部漁業(yè)局, 2014. 中國漁業(yè)統(tǒng)計(jì)年鑒 2014. 北京: 中國農(nóng)業(yè)出版社, 33, 56
孫成波, 鄧先余, 李鎮(zhèn)泉等, 2008. 北部灣野生日本囊對(duì)蝦(Marsupenaeus japonicus)體重和形態(tài)性狀的關(guān)系. 海洋與湖沼, 39(3): 263—268
李義軍, 李 婷, 王 平等, 2010. 日本囊對(duì)蝦(Marsupenaeus japonicus)3個(gè)野生種群和 1個(gè)養(yǎng)殖種群的形態(tài)差異與判別分析. 海洋與湖沼, 41(4): 500—504
范志勤, 1973. 大耳林姬鼠數(shù)量配置、體型和突出部分的大小與環(huán)境溫度的關(guān)系. 動(dòng)物學(xué)報(bào), 19(1): 54—60
董宏標(biāo), 蘇永全, 毛 勇等, 2014. 日本囊對(duì)蝦 2種形態(tài)變異類型群體形態(tài)性狀對(duì)體質(zhì)量的影響效果分析. 廈門大學(xué)學(xué)報(bào)(自然科學(xué)版), 53(2): 289—296
富 裕, Diouf F, 陳漢春等, 2014. 密植滸苔對(duì)冬季露天池塘池底水溫、酸堿度和溶解氧的影響效應(yīng). 浙江海洋學(xué)院學(xué)報(bào)(自然科學(xué)版), 33(5): 389—393, 413
蔡曉鵬, 游欣欣, 曾凡榮等, 2010. 中國沿海日本囊對(duì)蝦 5個(gè)地理群體間形態(tài)差異比較分析. 中國水產(chǎn)科學(xué), 17(3):478—486
Allen J A, 1877. The influence of physical conditions in the genesis of species. Radical Review, 1: 108—140
Bergmann C, 1847. über die Verh?ltnisse der W?rme?konomie der Thiere zu ihrer Gr?sse. G?ttinger Studien, G?ttingen,3(1): 595—708
Tsio K H, Wang Z Y, Chu K H, 2005. Genetic divergence between two morphologically similar varieties of the kuruma shrimpPenaeus japonicus. Marine Biology, 147(2): 367—379
Tsio K H, Wang Z Y, Chu K H, 2007. Molecular population structure of the kuruma shrimpPenaeus japonicusspecies complex in western Pacific. Marine Biology, 150(6):1345—1364
Wagner E J, Arndt R E, Brough M, 2001. Comparative tolerance of four stocks of cutthroat trout to extremes in temperature,salinity, and hypoxia. Western North American Naturalist,61(4): 434—444
