香煙提取物通過減少HDAC2表達誘導小鼠C2C12成肌細胞衰老*
2015-04-17 02:16:32蘇文燕劉文婷何志義
中國病理生理雜志 2015年4期
關鍵詞:小鼠
蘇文燕, 劉文婷, 楊 霞, 白 晶, 何志義
(廣西醫科大學第一附屬醫院呼吸內科,廣西 南寧 530021)
?
香煙提取物通過減少HDAC2表達誘導小鼠C2C12成肌細胞衰老*
蘇文燕, 劉文婷, 楊 霞, 白 晶△, 何志義
(廣西醫科大學第一附屬醫院呼吸內科,廣西 南寧 530021)
目的: 探討香煙提取物(CSE)能否引起小鼠C2C12成肌細胞衰老,并研究成肌細胞衰老與組蛋白去乙酰化酶2(HDAC2)的關系。方法: 培養C2C12細胞株,分化為成熟成肌細胞,觀察CSE干預對成肌細胞衰老和HDAC2表達的影響,采用real-time PCR和Western blot方法分別檢測HDAC2 mRNA和蛋白的表達;β-半乳糖苷酶染色檢測衰老細胞的百分率。結果: MTT法測定最佳CSE濃度與干預時間分別為60 mL/L和24 h。CSE干預后β-半乳糖苷酶染色陽性細胞數增加,同時伴有HDAC2 mRNA和蛋白表達的減少。四溴苯三唑(TBB)在促進HDAC2 mRNA和蛋白表達的同時,β-半乳糖苷酶染色陽性細胞數減少;用HDAC2的特異性阻滯劑丙戊酸抑制HDAC2 mRNA和蛋白的表達時,β-半乳糖苷酶染色陽性細胞數增加。結論: 香煙提取物可通過減少小鼠C2C12成肌細胞HDAC2的表達促進細胞老化。
香煙提取物; C2C12成肌細胞; 衰老; 組蛋白去乙酰化酶2
慢性阻塞性肺疾病(chronic obstructive pulmonary disease,COPD)不僅僅是一種局限于呼吸道和肺部的疾病,還是一種可以累及肺外各器官的全身疾病,特別是骨骼肌萎縮和功能障礙,嚴重影響患者的生活質量及預后,導致運動耐力下降和生活質量的下降[1]。目前的治療措施如營養支持治療、康復訓練等不能有效預防和改善COPD患者骨骼肌的變化[2-3],最根本的原因是對這一病理狀態的發生機制還未完全清楚。近年研究顯示[4-5],COPD是一種加速老化的肺部疾病,COPD發病機制與衰老密切相關,煙草煙霧/有害氣體是聯系老化肺臟與COPD的關鍵因素,煙草煙霧及其它污染物等環境氣體可能加速了肺臟的衰老,或導致炎癥清除缺陷,并在衰老過程中促進肺功能進行性惡化,肺部炎癥加劇和肺氣腫等結構性改變。近年來也已證實一些新型抗衰老分子可以成為COPD治療的新靶點[6]。然而COPD的肺外表現,尤其是有關骨骼肌細胞衰老的研究目前尚少。本實驗通過體外培養小鼠成肌細胞(C2C12細胞株)分化成成熟成肌細胞,觀察香煙提取物干預后成肌細胞衰老和抗老化分子組蛋白去乙酰化酶2(histone deacetylase 2,HDAC2)表達的變化,探討香煙提取物對小鼠C2C12成肌細胞衰老的影響及其相關機制,為進一步尋求COPD骨骼肌病變的發病機制和新防治策略提供實驗依據。
材 料 和 方 法
1 材料
成肌細胞C2C12細胞株(中科院上海細胞庫);DMEM 培養基、胎牛血清、馬血清(Gibco);胰蛋白酶(碧云天公司);香煙(萬寶路公司);HDAC2激動劑四溴苯三唑(4, 5, 6, 7-tetrabromobenzotriazole,TBB,Sigma);HDAC2特異性抑制劑丙戊酸(南寧生利德公司);β-半乳糖苷酶染色試劑盒(上海杰美公司);RNA逆轉錄試劑盒、SYBR Premix Ex TaqII染料法熒光定量試劑盒(TaKaRa);小鼠HDAC2 I抗(CST)及II抗(中杉公司); 抗小鼠抗GAPDH I抗(Proteintech)。
2 方法
2.1 C2C12細胞的培養 10%牛血清培養基培養C2C12細胞,放入5% CO2、37 ℃培育箱培養,待細胞長到80%左右,更換成分化培養基(含10%馬血清的培養基)繼續培養。分化4~5 d后待95%以上的細胞出現肌管[7-8]時即可用于實驗。
2.2 MTT法和細胞β-半乳糖苷酶染色觀察最佳CSE干預濃度和時間 收集對數期的細胞,制成濃度為1×107/L細胞懸浮液,在 96孔板中每孔加入100 μL細胞懸液,每個濃度設5個平行孔。第1列只加100 μL培養基,作為調零孔;第2列加入100 μL細胞懸液,不加CSE,作為空白對照組。待細胞完全貼壁后,其余各列分別加入CSE,使得終濃度分別為20、40、60、80、100、120 mL/L。培養0、12、24、48、72 h后每孔加入20 μL MTT(5 g/L),4 h后小心棄上清,每孔加入150 μL DMSO,置搖床上低速振蕩10 min,使結晶物充分溶解,在酶聯免疫檢測儀波長為 490 nm處測量各孔的吸光度(A)值。
收集已分化好的細胞制成細胞懸液接種于6孔板中,細胞數為每孔5×105。待細胞完全貼壁后加入不同濃度的CSE干預, CSE終濃度分別為0、20、40、60、80、100和120 mL/L。干預24 h后行β-半乳糖苷酶染色,藍染細胞為陽性細胞。200 倍顯微鏡隨機選取視野,計數每個視野里陽性細胞,每個濃度觀察5 個視野,染色重復3次,取平均值。
2.3 Western blot檢測CSE刺激下HDAC2蛋白的表達 收集已分化好的細胞制成細胞懸液接種于6孔板中,細胞數為每孔5×105。細胞分為24 h對照組、24 h CSE組、48 h對照組、48 h CSE組、72 h對照組、72 h CSE組。待細胞完全貼壁后加入終濃度為60 mL/L CSE干預,干預時間如分組時間所示。待干預結束后各組細胞用胰酶消化,用PBS清洗3遍,1 000 r/min離心10 min。棄上清,加入100 μL細胞裂解液(碧云天公司),加入1 μL PMSF后在冰上裂解30 min,每間隔10 min振蕩30 s,離心后棄沉淀,吸取2 μL上清測蛋白濃度。取總蛋白50 μg上樣,SDS-PAGE 分離,然后電轉移至PVDF膜,50 g/L脫脂奶粉封閉1 h,封閉后的濾膜再分別與小鼠抗HDAC2抗體(稀釋比為 1∶1 000)、小鼠抗GAPDH抗體(1∶10 000) 4 ℃溫育過夜。HRP 偶聯的 IgG 作為 II 抗(稀釋比為 1∶10 000)室溫溫育 1 h,暗室膠片曝光顯影、定影。Bio-Rad公司的全自動掃描儀掃描膠片,Quantity One軟件分析結果。
2.4 觀察CSE刺激下成肌細胞HDAC2與衰老的關系 取成熟的成肌細胞接種于6孔板中,分為:(1)對照組(不加任何干預,培養48 h);(2)CSE組(不加任何干預培養24 h后,加入終濃度為60 mL/L CSE繼續孵育24 h);(3)TBB組(終濃度為25 μmol/L,孵育48 h);(4)TBB+CSE組(TBB終濃度為25 μmol/L,預孵育24 h后加入終濃度為60 mL/L CSE繼續孵育24 h);(5)丙戊酸組(終濃度為20 μmol/L,孵育48 h);(6)丙戊酸+CSE組(丙戊酸終濃度為20 μmol/L,預孵育24 h后加入60 mL/L CSE繼續孵育24 h)。干預結束后,收集各組細胞,經β-半乳糖苷酶染色;Western blot 方法檢測HDAC2蛋白表達;real-time PCR檢測各組細胞中HDAC2 的mRNA表達,方法如下:將上述每組細胞均加入1 mL TRIzol提取總RNA,然后按照TaKaRa逆轉錄試劑盒說明逆轉錄成cDNA,用實時熒光定量PCR儀(Roche)行逆轉錄反應,反應體系為20 μL。HDAC2的上游引物為5’-AGCCCATGGCGTACAGTCAA-3’,下游引物為5’-GGGATGACCCTGGCCATAATAA-3’;GAPDH的上游引物為5’-TGTGTCCGTCGTGGATCTGA-3’,下游引物為5’-TTGCTGTTGAAGTCGCAGGAG-3’。反應條件為95 ℃ 30 s;95 ℃ 5 s,60 ℃ 20 s,擴增40個循環。擴增結束后基因的相對表達量用2-ΔΔCt法計算比較。
3 統計學處理
應用SPSS 16.0統計分析軟件檢測實驗數據的正態性分布,正態分布的計量資料采用均數±標準差(mean±SD)表示。組間比較用方差分析,正態分布不齊的計量資料用秩和檢驗。以P<0.05為差異有統計學意義。
結 果
1 C2C12細胞分化為成熟成肌細胞
細胞呈成纖維狀態。分化第3天出現部分肌細胞。至第4天、第5天,95%以上細胞分化為長圓柱形,每個細胞含多個細胞核,核呈扁橢圓形的成熟骨骼肌細胞,可用于實驗,見圖1。
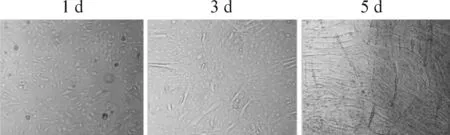
Figure 1.The different stages of C2C12 cells differentiated into skeletal muscle cells.
圖1 C2C12細胞逐漸分化為成熟成肌細胞
2 不同濃度CSE對成肌細胞活性和衰老的影響
MTT法顯示隨著CSE濃度增加,干預時間越長,對成肌細胞活性的抑制作用越大。濃度小于60 mL/L的CSE在12 h、24 h對細胞生長并無明顯抑制作用。當濃度大于80 mL/L時,細胞生長明顯受到抑制,見圖2、表1。

Figure 2.TheAvalues of the C2C12 myoblasts treated with different concentrations of CSE at various time points. Mean±SD.n=5.
圖2 不同濃度CSE干預下C2C12成肌細胞各時點的A值
β-半乳糖苷酶定位在細胞質,是衰老細胞的特異性指標,染成藍色為陽性細胞。CSE干預24 h內,當CSE濃度在一定濃度范圍內(CSE<60 mL/L),隨著CSE濃度的增加,衰老細胞數量增加,繼續增加CSE濃度時,細胞出現形態縮小,核碎裂,繼而死亡。故本實驗將CSE干預的終濃度和時間分別設定為60 mL/L、24 h,見圖3。
表1 不同濃度CSE干預下C2C12成肌細胞各時點的A值
Table 1.TheAvalues of the C2C12 skeletal muscle myocytes treated with different concentrations of CSE at various time points (Mean±SD.n=5)

CSE0h12h24h48h72h0mL/L0.94±0.070.94±0.070.95±0.020.95±0.070.98±0.0620mL/L0.94±0.060.94±0.060.93±0.050.89±0.06*0.85±0.08*40mL/L0.95±0.070.94±0.070.92±0.030.87±0.06*0.85±0.06*60mL/L0.94±0.080.93±0.040.91±0.050.84±0.07*0.80±0.07*80mL/L0.93±0.060.76±0.06*0.62±0.07*0.57±0.04*0.51±0.06*100mL/L0.94±0.040.67±0.06*0.58±0.07*0.54±0.05*0.48±0.07*120mL/L0.93±0.050.58±0.07*0.46±0.08*0.43±0.03*0.42±0.06*
*P<0.05vs0 mL/L group at the same time point.
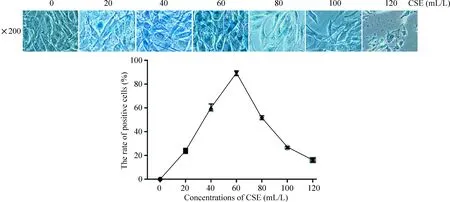
Figure 3.The positive cell rate of β-galactosidase staining in the C2C12 myoblasts treated with different concentrations of CSE.Mean±SD.n=3.
圖3 不同CSE濃度干預下小鼠C2C12成肌細胞內β-半乳糖苷酶的表達
3 CSE刺激下成肌細胞HDAC2蛋白的表達
在濃度為60 mL/L的CSE作用下,24 h CSE組、48 h CSE組、72 h CSE組的HDAC2蛋白表達均較相同時點的對照組顯著下降(P<0.05);隨著CSE干預時間的增加,HDAC2蛋白表達呈遞減趨勢,見圖4。
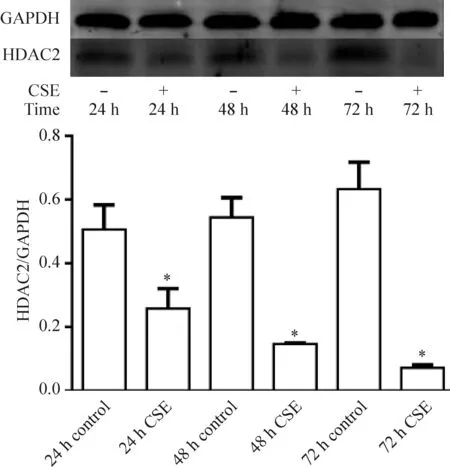
Figure 4.The protein expression of HDAC2 in the C2C12 myoblasts treated with CSE over time. Mean±SD.n=3.*P<0.05vscontrol group at the same time point.
圖4 CSE干預作用下小鼠C2C12成肌細胞HDAC2蛋白表達隨時間的變化
4 CSE刺激下成肌細胞HDAC2 mRNA、蛋白的表達與衰老的關系
CSE組HDAC2 mRNA表達較對照組明顯降低(P<0.05)。與對照組比較,TBB組HDAC2 mRNA表達增加(P<0.05),丙戊酸(valproic acid)組HDAC2 mRNA表達明顯降低(P<0.05)。與CSE組比較, TBB+CSE組HDAC2 mRNA表達明顯增加,與TBB組比較,TBB+CSE組HDAC2 mRNA表達降低(P<0.05)。與CSE組比較,丙戊酸+CSE組HDAC2 mRNA表達明顯降低(P<0.05)。與丙戊酸組比較,丙戊酸+CSE組HDAC2 mRNA表達降低更明顯(P<0.05),見圖5。
與對照組比較,CSE組干預后 HDAC2 的蛋白表達明顯降低(P<0.05),TBB組的HDAC2 蛋白表達明顯增加,丙戊酸組的HDAC2 蛋白表達明顯減少。與CSE組比較,加入TBB聯合干預組的HDAC2 蛋白表達均明顯增加,但與TBB組比較,CSE+TBB組的HDAC2蛋白表達降低(P<0.05)。與CSE組比較,加入丙戊酸聯合干預組的HDAC2 蛋白表達明顯降低(P<0.05),見圖6。

Figure 5.The mRNA expression of HDAC2 in the C2C12 myoblasts treated with CSE plus the agonist or antagonist of HDAC2. Mean±SD.n=3.*P<0.05vscontrol group;#P<0.05vsCSE group;△P<0.05vsTBB group;&P<0.05vsvalproic acid group.
圖5 CSE對小鼠C2C12成肌細胞HDAC2的 mRNA表達的影響
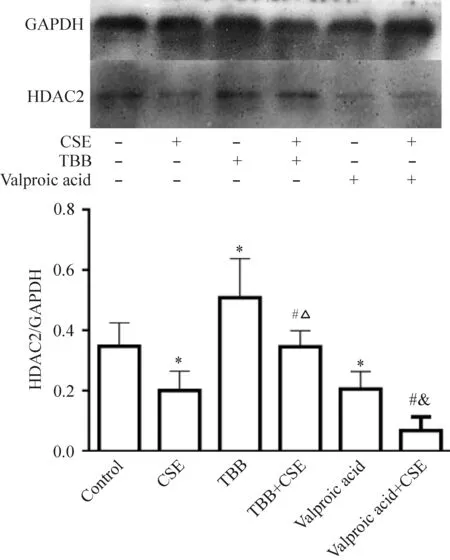
Figure 6.The protein expression of HDAC2 in the C2C12 myoblasts treated with CSE plus the agonist or antagonist of HDAC2. Mean±SD.n=3.*P<0.05vscontrol group;#P<0.05vsCSE group;△P<0.05vsTBB group;&P<0.05vsvalproic acid group.
圖6 CSE對小鼠C2C12成肌細胞HDAC2蛋白表達的影響
與對照組比較,CSE組的β-半乳糖苷酶染色陽性細胞數較對照組增加(P<0.05),TBB組沒有出現β-半乳糖苷酶染色陽性細胞,丙戊酸組的β-半乳糖苷酶染色陽性細胞數明顯增加。與CSE組比較,CSE+TBB組的β-半乳糖苷酶染色陽性細胞數明顯減少;與TBB組比較,CSE+TBB組的β-半乳糖苷酶染色陽性細胞數增多;與CSE組比較,CSE+丙戊酸組的β-半乳糖苷酶染色陽性細胞數明顯增加;與丙戊酸組比較,CSE+丙戊酸組的β-半乳糖苷酶染色陽性細胞數增多更明顯(P<0.05),見圖7。
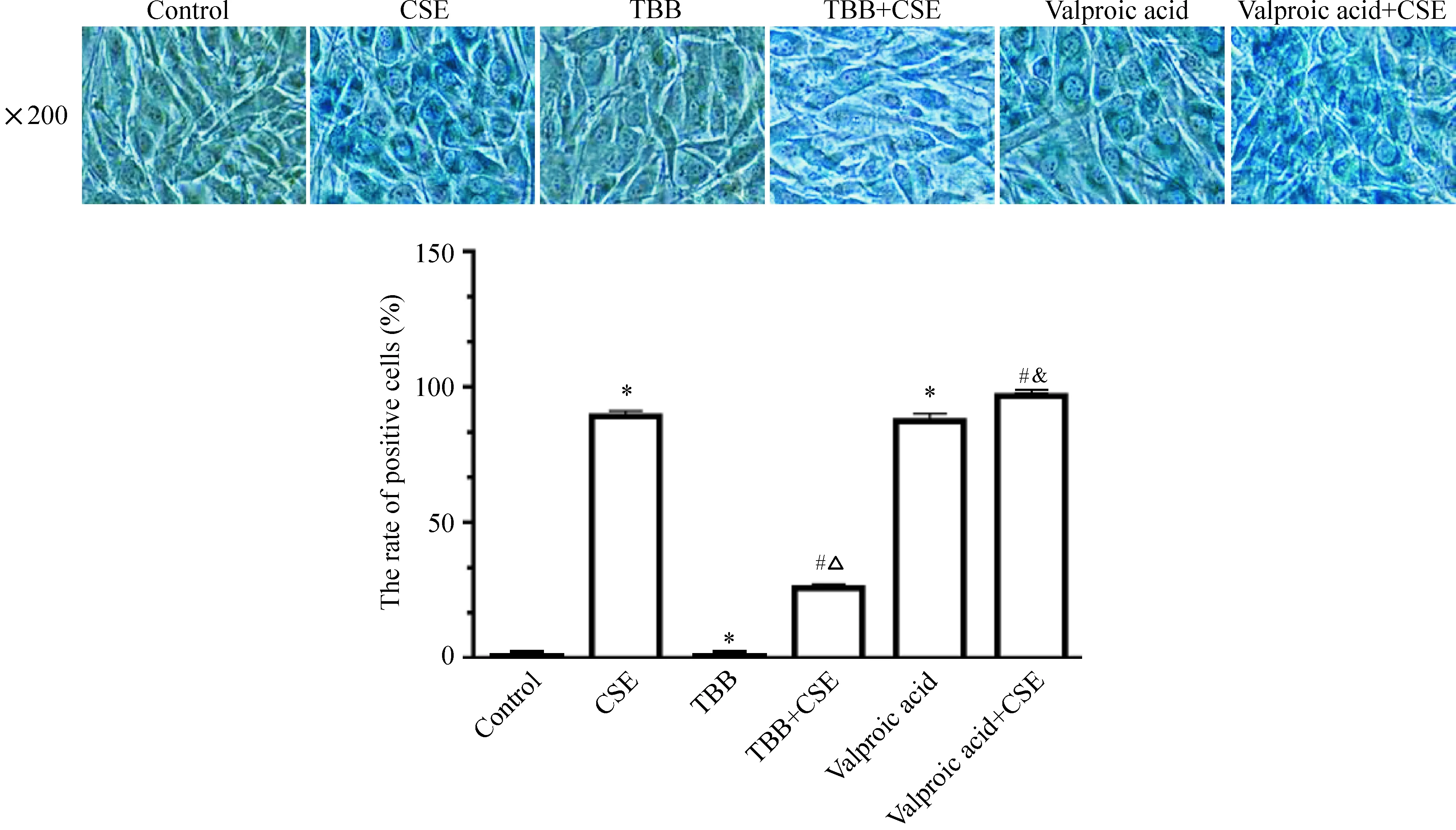
Figure 7.The positive cell rate of β-galactosidase staining in the C2C12 myoblasts treated with CSE plus the agonist or antagonist of HDAC2. Mean±SD.n=3.*P<0.05vscontrol group;#P<0.05vsCSE group;△P<0.05vsTBB group;&P<0.05vsvalproic acid group.
圖7 各組細胞β-半乳糖苷酶染色陽性細胞率
討 論
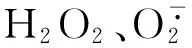
為了進一步研究吸煙能否誘導HDAC2下降促進成肌細胞衰老,我們將濃度小于60 mL/L的 CSE與成肌細胞共同培養,發現在12 h、24 h對細胞生長并無明顯抑制作用。當濃度大于80 mL/L時CSE對細胞生長有明顯抑制作用。故本實驗將CSE干預的終濃度和時間分別設定為60 mL/L和24 h。β-半乳糖苷酶染色是特異性檢測衰老的指標。我們觀察到,CSE干預可誘導成肌細胞衰老,當CSE濃度≤60 mL/L時,隨著CSE濃度的增加,成肌細胞衰老細胞百分率增加。同時我們觀察到,60 mL/L CSE在促進成肌細胞衰老的同時伴有HDAC2蛋白表達的下降,而且HDAC2蛋白表達的下降隨著CSE作用時間的延長而進一步減少。為了進一步研究HDAC2與衰老的相關性,我們以四溴苯三唑激活HDAC2[15]、HDAC2 特異性阻滯劑丙戊酸抑制HDAC2[16],發現與CSE組比較,當TBB促進HDAC2對 mRNA和蛋白表達的同時,CSE+TBB組成肌細胞衰老細胞數量減少,TBB可抑制CSE誘導的細胞衰老,丙戊酸抑制HDAC2 的mRNA和蛋白的表達,CSE+丙戊酸組成肌細胞衰老細胞數量增加,丙戊酸可加重CSE誘導的細胞衰老,提示干預HDAC2的 mRNA和蛋白表達可以影響C2C12成肌細胞的衰老,吸煙通過抑制HDAC2的表達促進成肌細胞的衰老。
對骨骼肌細胞衰老分子學機制的研究可以使我們更深入了解COPD全身反應的機制,并將進一步有助于拓寬COPD 藥物治療的新靶點。目前關于COPD骨骼肌細胞老化的研究尚少,其具體途徑和機制有待于進一步研究。
[1] Gosker HR, Wouters EF, van der Vusse GJ, et al.Skeletal muscle dysfunction in chronic obstructive pulmonary disease and chronic heart failure: underlying mechanisms and therapy perspectives[J]. Am J Clin Nutrition, 2000,71(5): 1033-1047.
[2] Zainuldin R, Mackey MG, Alison JA. Optimal intensity and type of leg exercise training for people with chronic obstructive pulmonary disease[J]. Cochrane Database Syst Rev, 2011, 11:CD008008.
[3] Ferreira IM, Brooks D, White J, et al. Nutritional supplementation for stable chronic obstructive pulmonary disease[J]. Cochrane Database Syst Rev, 2012, 12:CD000998.
[4] Ito K, Barnes PJ. COPD as a disease of accelerated lung aging[J]. Chest J, 2009, 135(1):173-180.
[5] MacNee W. Accelerated lung aging: a novel pathogenic mechanism of chronic obstructive pulmonary disease (COPD) [J]. Biochem Soc Transactions, 2009, 37(Pt 4):819-823.
[6] Rahman I, Kinnula VL, Gorbunova V, et al. SIRT1 as a therapeutic target in inflammaging of the pulmonary disease[J]. Prev Med, 2012, 54 Suppl:S20-S28.
[7] Jové M, Planavila A, Sánchez RM, et al. Palmitate induces tumor necrosis factor-α expression in C2C12 skeletal muscle cells by a mechanism involving protein kinase C and nuclear factor-κB activation[J]. Endocrinol, 2006, 147(1):552-561.
[8] 何志義,梁 毅,梁秋麗,等. 香煙對小鼠C2C12成肌細胞分化的影響[J]. 中國病理生理雜志, 2010,26(5):881-884.
[9] Adenuga D, Yao H, March TH, et al. Histone deacetylase 2 is phosphorylated, ubiquitinated, and degraded by cigarette smoke[J]. Am J Respir Cell Mol Biol, 2009, 40 (4):464-473.
[10]Hongwei Y, Irfan R. Role of histone deacetylase 2 in epigenetics and cellular senescence: implications in lung inflammaging and COPD[J]. Am J Physiol: Lung Cell Mol Physiol, 2012, 303(7):L557-L566.
[11]Tsuji T, Aoshiba K, Nagai A. Alveolar cell senescence in patients with pulmonary emphysema[J]. Am J Respir Crit Care Med, 2006, 174(8):886-893.
[12]Morlá M, Busquets X, Pons J, et al. Telomere shortening in smokers with and without COPD[J]. Eur Respir J, 2006, 27(3):525-528.
[13]Müller KC, Welker L, Paasch K, et al. Lung fibroblasts from patients with emphysema show markers of senescenceinvitro[J]. Respir Res, 2006, 7:32.
[14]閆海龍,龔勇珍.氧化應激及p16和p53/p21與細胞衰老關系的研究進展[J]. 醫學綜述, 2011, 17(5):682-685.
[15]Adenuga D, Yao H, March TH, et al. Histone deacetylase 2 is phosphorylated, ubiquitinated, and degraded by cigarette smoke[J]. Am J Respir Cell Mol Biol, 2009, 40(4):464-473.
[16]Kr?mer OH, Zhu P, Ostendorff HP, et al. The histone deacetylase inhibitor valproic acid selectively induces proteasomal degradation of HDAC2[J]. EMBO J, 2003, 22(13):3411-3420.
Cigarette smoke extract promotes senescence by reducing expression of HDAC2 in C2C12 myoblasts
SU Wen-yan, LIU Wen-ting, YANG Xia, BAI Jing, HE Zhi-yi
(DepartmentofRespiratoryMedicine,FirstAffiliatedHospital,GuangxiMedicalUniversity,Nanning530021,China.E-mail:bj1312002@aliyun.com)
AIM: To investigate whether the cigarette smoke extract (CSE) causes senescence of C2C12 myoblasts and the relationship between senescence and histone deacetylase 2(HDAC2). METHODS: Murine C2C12 cells were induced to differentiate into myoblasts. The HDAC2 activator and inhibitor were used to investigate the effects of CSE in the myoblasts on cell senescence and the expression of HDAC2. The expression of HDAC2 at mRNA and protein levels was determined by real-time PCR and Western blot, respectively, and the positive cell rate of β-galactosidase staining for cell senescence was also detected. RESULTS: The optimal concentration of CSE was 60 mL/L and the intervention time was 24 h. After the intervention of CSE, the positive cell rate of β-galactosidase staining was increased, accompanied with the reduction of HDAC2 expression at mRNA and protein levels. The expression of HDAC2 at mRNA and protein levels was increased by 4, 5, 6, 7-tetrabromobenzotriazole (TBB), accompanied with the reduction of positive cell rate of β-galactosidase staining. Furthermore, when HDAC2 expression at mRNA and protein levels was reduced by HDAC2 inhibitor valproic acid, the positive cell rate of β-galactosidase staining was increased.CONCLUSION: CSE promotes the senescence by reducing the expression of HDAC2 in C2C12 myoblasts.
Cigarette smoke extract; C2C12 myoblasts; Senescence; Histone deacetylase 2
1000- 4718(2015)04- 0738- 06
2014- 11- 24
2015- 02- 04
國家自然科學基金資助項目(No. 81260011)
R363; R563
A
10.3969/j.issn.1000- 4718.2015.04.029
△通訊作者 Tel: 0771-5356702; E-mail: bj1312002@aliyun.com
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
