細粒棘球蚴線粒體CO1基因的克隆與序列分析
2015-05-04 13:44:37于晶峰劉曉松常建華楊曉野楊蓮茹李秀霞
中國人獸共患病學報 2015年6期
于晶峰,李 濱,劉曉松,常建華,楊曉野,王 瑞,楊蓮茹,李秀霞,木 蘭
細粒棘球蚴線粒體CO1基因的克隆與序列分析
于晶峰1,3,李 濱1,劉曉松2,常建華2,楊曉野1,王 瑞1,楊蓮茹1,李秀霞3,木 蘭3
目的 研究細粒棘球蚴(Echinococcus granulasus)種間遺傳標記特點,為該蟲種的分子分類學研究提供基礎。方法 在內蒙古地區從羊肝臟采集細粒棘球蚴,肝包蟲病患者手術剝離囊包后,簽署知情同意書,采集細粒棘球蚴。提取蟲體DNA,擴增線粒體細胞色素氧化酶1(CO1)基因,將其克隆到PGM-T載體后,用PCR技術鑒定陽性菌落,并進行測序,運用DNAstar5.0 軟件計算序列間的相似性,計算遺傳距離,同時,應用MEGA4.0軟件的最大似然法(ML-Maximum Likelihood)和鄰接法(NJ-Neighbor Joining)構建系統發育樹,進行聚類分析。結果 細粒棘球蚴DNA擴增出的羊株、人株CO1基因序列片段長度為936 bp。同其他親緣關系較近的屬CO1基因序列比對:羊株、人株細粒棘球蚴CO1序列與黃花棘球絳蟲的同源性最高,分別為91.9%、91.2%;羊株、人株細粒棘球蚴CO1序列與黃花棘球絳蟲位于同一分支,自展值(Boostrap)最高,均為100%。羊株、人株細粒棘球蚴CO1序列所屬分支與原頭目的Proteocephalus macrocephalus所屬分支相隔較遠。結論 CO1基因序列穩定保守,無宿主特異性,可作為細粒棘球蚴理想的種間遺傳標記。
細粒棘球蚴;CO1基因;克隆;序列分析
細粒棘球絳蟲(Echinococcusgranulosus)屬帶科、棘球屬,又稱包生絳蟲。成蟲寄生于犬科食肉動物,幼蟲(棘球蚴)寄生于人和多種食草類家畜及其它動物,引起一種嚴重的人獸共患病,稱棘球蚴病或包蟲病。棘球蚴病分布地域廣泛,隨著世界畜牧業的發展而不斷擴散,已成為全球性重要的公共衛生和經濟問題[1-2]。細粒棘球絳蟲的蟲株鑒定非常復雜,以往主要利用其形態學特征結合其他方面的資料(如流行病學、生物化學、同工酶分析)作為蟲株鑒定的根據[3]。雖然利用形態學和生物學特征能為蟲株的鑒定提供一些有價值的資料,但由于受到多種因素(如宿主和環境)的影響,并不能完全反映其基因水平的差異。而DNA序列作為遺傳信息的載體,在世代傳遞過程中,除保有遺傳穩定外,還不斷的發生變異。因而,DNA序列分析對物種間的進化關系的確立、種間和種內親緣關系的確立均具有重要意義[4-5]。近年來,隨著生物信息學的發展,將動物寄生蟲基因組DNA中的未知序列與已知序列(如GenBank、Ensembl、UCSC)進行比較,進而確定蟲種的種屬和分類。已成為分子生物學分析動物寄生蟲進化關系的一個強有力的工具[6-7]。
1 材料與方法
1.1 材料
1.1.1 試驗樣品細粒棘球蚴包囊采自內蒙古錫盟西烏旗綿羊屠宰場,和錫林浩特市醫院包蟲病患者。錫盟西烏旗綿羊屠宰場調查成年綿羊與幼年綿羊共2 313只,患羊數量為30只,18名錫林浩特市某醫院B超檢查疑似包蟲病患者,手術后經病理證實確診為包蟲病,患者均簽署知情同意書。收集囊包后用10 mL注射器將囊液吸出,離心1 min 30 s,使囊液與原頭蚴分離,-20 ℃保存。
1.1.2 菌種、載體及主要試劑 大腸桿菌Top10感受態細胞、血液組織細胞基因組提取試劑盒(DP304)、瓊脂糖膠回收試劑盒(DP209-03)、pGM-T連接試劑盒( VT202-02)均購自天根(北京)生化科技有限公司;Premix Ex Taq(RR003A)、DL2000 DNA Marker、氨芐青霉素(Amp)、5-溴-4-氯-3-吲哚-β-D半乳糖苷(X-gal)、異丙基-β-D-硫代半乳糖苷(IPTG)、10×Loading Buffer均購自大連寶生物工程有限公司;核酸染料,購自北京市賽百盛基因技術有限公司;其它鹽酸,氫氧化鈉等常規生化試劑由內蒙古農業大學獸醫學院寄生蟲學研究室提供。
1.2 基因組DNA提取及PCR擴增 細粒棘球蚴原頭蚴基因組DNA的提取按照血液組織細胞基因組提取試劑盒進行。根據楊俊克[8]報道的細粒棘球蚴CO1基因的引物序列,上游引物(CO1-F):5′-TTGTTAGGTGGTTTGTCTGA-3′和下游引物(CO1-R):5′-GGCCATCACAAATAAACAT-3′由生工生物工程(上海)股份有限公司合成。
以提取的細粒棘球蚴原頭蚴基因組DNA為模板,進行目的基因擴增。體系為80 μL(各組分用量:Premix Taq 40 μL,CO1-F 4 μL,CO1-R 4 μL模板4 μL,dH2O補足至80 μL)。PCR反應條件為:95 ℃預變性5 min;95 ℃變性40 s,56 ℃退火30 s,72 ℃延伸1 min 20 s, 35個循環;72 ℃完全延伸10 min。反應結束后,對PCR產物分別進行1%瓊脂糖凝膠電泳檢測,于紫外燈下觀察結果。
1.3 擴增片段的克隆與測序 利用瓊脂糖膠回收試劑盒對PCR產物的目的片段進行純化。根據pGM-T載體說明書將膠回收的PCR產物與PGM-T載體進行連接,目的PCR片段,XμL,pGM-T vector 1 μL,10×T4 DNA Ligation Buffer 1 μL,ddH2O補足至10 μL。載體與目的片段的摩爾比控制在1∶3~1∶8。輕彈混勻短暫離心后16 ℃水浴過夜連接。待反應結束后立即將PCR管置于冰上。
連接產物轉化至感受態細胞,進行藍白斑篩選。挑取白色菌落,置于2 mL LB液體培養基中(含有Amp),150 r/min、37 ℃振蕩培養12~16 h,取1 μL進行菌液PCR鑒定。PCR反應體系為20 μL,反應條件同1.2的PCR擴增。PCR產物經瓊脂糖凝膠電泳檢測,有目的條帶的陽性菌液送生工生物工程(上海)股份有限公司測序。
1.4 序列分析 應用NCBI中的nucleotide blast對測得序列細粒棘球蚴的CO1基因進行同源搜索。綜合考慮序列覆蓋率(Query coverage)、最大序列相似度(Max ident)、隨機匹配可能性(E value)3個因素,選取圓葉目、裂頭目(Diphyllobothriide)、原頭目(Proteocephalidea)等其他科屬種的CO1基因的序列(表1),運用DNAstar5.0 軟件計算序列間的相似性,計算遺傳距離,同時,應用MEGA4.0軟件的最大似然法(ML-Maximum Likelihood)和鄰接法(NJ-Neighbor Joining)構建系統發育樹,進行聚類分析。系統發育樹進行自舉檢驗(bootstrap),檢驗次數為1 000。
2 結 果
2.1 PCR擴增結果 PCR擴增產物經1%瓊脂糖凝膠電泳檢測,結果見圖1。結果表明CO1基因在約936 bp處獲得了特異性條帶,與預期片段大小吻合,無非特異性的擴增條帶。
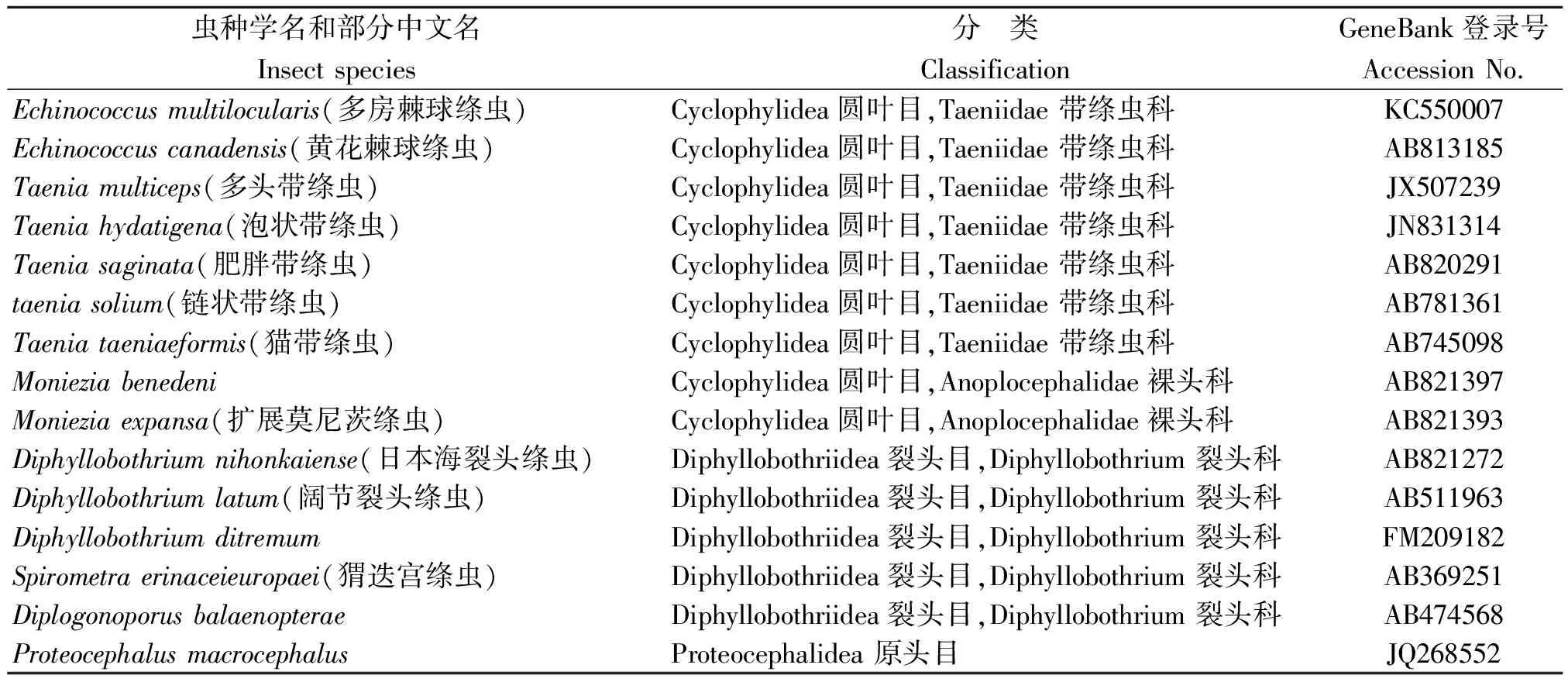
表1 15種絳蟲線粒體CO1基因相關資料

M:DL-2000分子量標記;1:西烏旗羊株樣本;2~7:錫林浩特人株樣本;8:陰性對照
M:DL 2000 DNA marker;1:Xuwuqi sheep samples;2~7:Xilinhaote human samples;8:Negative control
圖1 細粒棘球蚴CO1基因PCR產物電泳結果
Fig.1 Electrophoresis results of PCR products of CO1 gene fromEchinococcusgranulosus
2.2 克隆結果 陽性菌液經PCR鑒定和1%瓊脂凝膠電泳檢測后,在800 bp~1 000 bp之間出現了清晰的特異性條帶(見圖2)。
2.3 測序結果

M:DL-2000分子量標記;1:西烏旗羊株樣本;2~7:錫林浩特樣人株本;8:陰性對照
M:DL 2000 DNA marker;1:Xuwuqi sheep samples;2~7: Xilinhaote human samples;8:Negative control
圖2 陽性克隆菌液PCR鑒定電泳結果
Fig.2 Electrophoresis results of positive clone bacterium by PCR amplification
2.3.1 西烏旗羊株細粒棘球蚴CO1序列 西烏旗羊株細粒棘球蚴以CO1-F、CO1-R為引物,通過PCR糖擴增獲得了CO1基因部分序列,經克隆測序、去載體接頭等處理獲得了長度為936 bp的序列,其A、G、T、C的含量分別為15.60%、25.00%、48.72%、10.68%,A+T含量為64.32%。具體序列如下:

The shaded region was the prime poistion
2.3.2 錫林浩特市人株細粒棘球蚴CO1序列 錫林浩特市人株細粒棘球蚴DNA樣品經克隆測序所得的序列長度也為936 bp,其A、G、T、C的含量分別為15.60%、25.11%、47.54%、11.75%,A+T含量為63.14%。具體序列如下:

The shaded region was the prime poistion
2.3.3 序列比對結果 對比結果顯示西烏旗羊株與錫林浩特人株細粒棘球蚴的同源性為98.5%,變異率為1.5%,相應區域存在12個變異位點,突變類型均為轉換,即G與A置換和T與C置換。將獲得的羊株細粒棘球蚴CO1序列、人株細粒棘球蚴CO1序列與NCBI數據庫進行同源分析,通過nucleotide blast同源比對(http://blast.ncbi.nlm.nih.gov)結果表明羊株細粒棘球蚴CO1序列與蒙古細粒棘球蚴的CO1基因序列相似性最高,人株細粒棘球蚴CO1序列與伊朗細粒棘球蚴的CO1基因序列相似性最高,相似性均在99%以上,基因型均是G1型。羊株細粒棘球蚴CO1序列與人株細粒棘球蚴CO1序列的相似性也達到了99%。
2.3.4 基因序列同源性與遺傳距離分析 羊株細粒棘球蚴CO1序列、人株細粒棘球蚴CO1序列均與圓葉目、帶絳蟲科的黃花棘球絳蟲的同源性最高,分別為91.9%、91.2%;羊株細粒棘球蚴CO1序列、人株細粒棘球蚴CO1序列與圓葉目、帶絳蟲科的黃花棘球絳蟲的遺傳距離最小,分別為0.086、0.092。
采用ML法和NJ法構建的系統發育樹結果顯示羊株細粒棘球蚴CO1序列、人株細粒棘球蚴CO1序列與帶絳蟲科的黃花棘球絳蟲位于同一分支,系統發育樹的自舉檢驗值(Boostrap)最高,均為100%(圖3、圖4)。羊株細粒棘球蚴CO1序列、人株細粒棘球蚴CO1序列所屬分支與原頭目的Proteocephalus macrocephalus所屬分支相隔較遠。
3 討 論

圖3 基于CO1構建的17種絳蟲之間的系統發育樹(ML-Maximum Likelihood法)
Fig.3 Phylogenetic treebased CO1 among 17 kinds of tapeworm(ML-Maximum Likelihood method)

圖4 基于CO1構建的17種絳蟲之間的系統發育樹(NJ-NeighborJoining法)
Fig.4 Phylogenetic treebased CO1 among 17 kinds of tapeworm(NJ method)
mtDNA是生物體細胞內重要的細胞器,是能量產生與轉換的場所。線粒體因其進化速率快,母系遺傳、基因重組率低、獨立進化、基因組精簡等特點[3]。已成為重要的分子標記,廣泛的用于物種基因遺傳變異的研究,尤其更適合于種下分類學的研究[8]。目前mtDNA基因組及其相關編碼基因序列已廣泛應用于寄生蟲的分類和系統發生等方面的研究。在細粒棘球蚴研究方面,馬秀敏等(2007)[9]對新疆不同地區44個患有細粒棘球蚴病的病人進行了研究,根據細粒棘球絳蟲mtDNA rrnL的特征構建了PCR-RFLP技術,即可用于其基因型的鑒定。近年來,國內外專家陸續報道了有關細粒棘球絳蟲基因型的研究進展,而基因型之間的對比也是當今學術領域研究的熱點問題[10-13]。楊俊克等(2004年)[8]用線粒體DNA的CO1基因測序的方法,調查了采自青海省、甘肅省和新疆維吾爾族自治區的24株細粒棘球蚴分離株基因型的變異情況,結果表明不同地區CO1基因變異率均在1%以下,這與本實驗得出的結果相一致,即內蒙古兩地區的細粒棘球蚴線粒體CO1基因序列高度相似,不因宿主的不同而出現明顯的差異,這為將來全面了解內蒙古地區基因變異情況提供了理論依據。
進化樹的構建有助于我們了解物種的進化歷史,為生物學中物種的分類提供可靠的依據。通過比較不同絳蟲CO1基因序列顯示,雖然此基因在不同種屬絳蟲間有部分序列相當保守,但各種屬絳蟲間仍存在著特定的差異。這些差異與種系發生的遠近程度存在一定的關系。如細粒棘球蚴與同屬帶絳蟲科的黃花棘球絳蟲的同源性達到了91.9%;而與演化關系較為疏遠的Diplogonoporus balaenopterae則差異較大(同源性僅為75.2%),分屬不同的種屬(Diplogonoporus balaenopterae,裂頭目,裂頭科)。CO1基因的進化分析都將細粒棘球蚴與黃花棘球絳蟲劃到了同一個分支,這說明他們之間發生分歧的時間較短,可能在生活習性方面還存在某些相似性。
[1]Dakkak A. Echinococcosis / hydatidosis: a severe threat in Mediterranean countries[J]. Vet parasitological, 2010, 174 (1/2):2-11.
[2]Romig T, Dinkel A, Mackenstedt U. The present situation of echinococcosis in Europe[J]. Parasitol Int, 2006, 55(Suppl): S187-191.
[3]Li Bin.Epidemiological Investigation ofEchinococcusgranulosusand Analysis of CO1 and ND1 Gene Sequence[D].Department of Preventive Veterinary Medicine, Inner Mongolia Agricultural University,2013:14.(in Chinese) 李濱.細粒棘球蚴的流行病調查及CO1與ND1基因序列分析[D].內蒙古農業大學預防獸醫學,2013:14.
[4]Folmer O, Biack M, Hoeh W, et al. DNA primers for amplification and mitochondrial cytoehrome coxidase subunit 1 from diverse metazoan invertebrate[J]. Mo1 MarBio1 Biotechnol, 1994, 3:29.
[5]Tie YM, Ma JX, Wang YD. Investigation on cystic echinococcosis for sheep from Xinghai region[J]. Qinghai Agr Anim Husb, 2008, 38(7): 612-615. (in Chinese) 鐵永梅,馬建霞,王永殿.興海地區綿羊棘球蚴病調查[J].青海農牧業,2008, 38(7): 612-615. (in Chinese)
[6]Xu ZJ, Zhang CY, Tan YH, et al. Prevalence of hydatid disease in Heilongjiang Province[J]. Chin J Parasitol Parasit Dis, 2004, 22(2): 93. (in Chinese) 徐之杰,張崇友,鐔云輝,等.黑龍江省棘球蚴病流行概況[J].中國寄生蟲學和寄生蟲病,2004,22(2):93.
[7]Ishida N, Hasegawa T, Takeda K, et al. Polymorphic sequence in the D-loop region of equine mitochondrial DNA[J]. Animal Genetics, 1994, 25: 215-221.(in Chinese)
[8]Yang JK, Jia WZ, Jing tao,et al.Three provinces E.granulosus Gene Variation Analysis in china[J].Chinese Journal of Veterinary Science and Technology,2004,34(9):12-16.(in Chinese) 楊俊克,賈萬忠,景濤,等.我國三省區細粒棘球絳蟲基因的變異分析[J].中國獸醫科技,2004,34(9):12-16.
[9]Ma XM.WULAMU MT,Ding JB,et al.PCR-RFLP technique in E.granulosus seed strains Identification[J].Journal of Pathogen Biology,2008,3(4):281-287.(in Chinese) 馬秀敏,吾拉木馬木提,丁劍冰,等.PCR-RFLP技術在細粒棘球絳蟲種株鑒定中的應用[J].中國病原生物學雜志,2008,3(4):281-287.
[10]Feagin JE. Mitoehondrial genome diversity in parasites[J].Int J Parasitol, 2000, 30: 371-390.
[11]Tu BX, Zhang WL, Zhang WB, et al. Investigation onEchinococcusgranulosusinfection in camel from Alashan of Inner Mongolia[J]. Endem Dis Bull, 2003, 18(2): 99 (in Chinese) 圖布新,張文林,張文彬,等.內蒙古阿拉善左旗駱駝中細粒棘球蚴感染的調查[J].地方病通報,2003,18(2):99.
[12]Liu GH, Dai RS, Zhao GH, et al. Cloning and sequence analysis of mitochondrial cytochrome c oxidase subunit 1 fromTaeniahydatigenaisolates from Changsha and Xiangxi in Hunan province[J]. J Trop Med, 2009, 9(2): 117-120. (in Chinese) 劉國華,戴榮四,趙光輝,等.湖南長沙及湘西泡狀帶絳蟲分離株線粒體cox1基因的克隆及序列分析[J].熱帶醫學雜志,2009,9(2):117-120.
[13]Brown G G, Gadaleta G, Pepe G, et al. Structural conservation and variation in the D-loop containing region of vertebrate mitochondrial DNA[J]. J Mol Biol, 1986, 192: 503-511.
Yang Xiao-ye,Email:xiaoyeyang122@sohu.com;Mu lan,Email: wodetenghe@126.com
Cloning and sequence analysis of the partial CO1 gene within mitochondrial DNA ofEchinococcusgranulosus
YU Jing-feng1,3,LI Bin1,LIU Xiao-song2,CHANG Jian-hua2, YANG Xiao-ye1,WANG Rui1,YANG Lian-ru1,LI Xiu-xia3,MU Lan3
(1.KeyLaboratoryofClinicalDiagnosisandTreatmentTechnologyinAnimalDisease,MinistryofAgriculture;CollegeofVeterinaryMedicine,InnerMongoliaAgriculturalUniversity,Hohhot010018,China; 2.InnerMongoliaAcademyofAgricultural&AnimalHusbandrySciences,Hohhot010031,China; 3.SchoolofBasicMedicalScience,InnerMongoliaMedicalUniversity,Hohhot010110,China)
We investigated the inter-specific genetic marker ofEchinococcusgranulosus, which was used for further study in molecular classification. The genomic DNA ofEchinococcusgranulosuscollected from the infected sheep and people in Inner Mongolia were extracted. The mitochondrial cytochrome oxidase subunit 1 ( CO1) gene was amplified by PCR using universal primers, and then the PCR product was cloned into pGM-T vectors. The insert was sequenced successfully and compared with other cestodes sequences by DNAStar 5.0 and MEGA 4.0. Results showed that the PCR product was 936 bp in length. Compared with other CO1 gene sequences of related cestodes, the homology amongEchinococcusgranulosusandEchinococcuscanadensiswere 91.9% and 91.2%.EchinococcusgranulosusandEchinococcuscanadensiswere in the same branch, and bootstrap values (Boostrap) in the highest was 100%.Echinococcusgranulosusaffiliated branch of the original leaders of the branch belonged toProteocephalusmacrocephalusfar apart. It suggested that the CO1 gene sequences were stably conservative and non-host-specific, which can be used as an ideal species amongEchinococcusgranulosusgenetic markers.
Echinococcusgranulosus; CO1 gene; cloning; sequence analysis
國家公益性行業(農業)專項資助(No.201103008,No.201303037)
楊曉野,Email:xiaoyeyang122@sohu.com; 木 蘭, Email:wodetenghe@126.com
1.農業部動物疾病臨床診療技術重點實驗室/內蒙古農業大學獸醫學院,內蒙古呼和浩特 010018; 2.內蒙古農牧業科學院獸醫研究所,呼和浩特 010031; 3.內蒙古醫科大學基礎醫學院,呼和浩特 010110
Supported by the National Public Service Sectors (Agriculture) Special Funding(Nos.201103008 and No.201303037 )
10.3969/cjz.j.issn.1002-2694.2015.06.008
R383.3
A
1002-2694(2015)06-0532-05
2015-03-02;
2015-05-20
