加系SPF大白豬和長白豬群體遺傳學分析
2015-05-25 00:34:58賀希文高彩霞姜騫權金強蔡原曲連東
中國實驗動物學報 2015年6期
賀希文,高彩霞,姜騫,權金強,蔡原*,曲連東*
(1.甘肅農業大學動物科學技術學院,蘭州 730070;2.中國農業科學院哈爾濱獸醫研究所獸醫生物技術國家重點實驗室,哈爾濱 150001)
研究報告
加系SPF大白豬和長白豬群體遺傳學分析
賀希文1,2,高彩霞2,姜騫2,權金強1,蔡原1*,曲連東2*
(1.甘肅農業大學動物科學技術學院,蘭州 730070;2.中國農業科學院哈爾濱獸醫研究所獸醫生物技術國家重點實驗室,哈爾濱 150001)
目的研究中國農業科學院哈爾濱獸醫研究所從加拿大引進的SPF大白豬和長白豬的遺傳學背景。方法實驗采用19對微衛星引物對該群體進行群體遺傳學分析。結果19個位點在大白豬群中檢測到84個等位基因,長白豬群中檢測到89個等位基因。大白豬的平均多態信息含量和平均雜合度分別為0.5271和0.5877;長白豬的平均多態信息含量和平均雜合度分別為0.5652和0.6066。由于S0155、S0143、S0178、Sw857和Sw936位點內等位基因大小和含量差異顯著(P<0.01)可作為大白豬和長白豬品種鑒定的候選位點。F-統計和遷移率分析結果表明,群體內的分化較小,遺傳結構穩定。結論引進加系SPF純種大白豬和長白豬的遺傳結構與國內部分純種大白豬和長白豬相比更為穩定,可作為實驗動物模型應用于動物醫學和科學研究。
SPF豬;微衛星標記;實驗動物;遺傳多樣性
當今世界大白豬和長白豬是最重要的商業品系肉用豬,尤其以生長速度快、繁殖力高、適應性強和瘦肉率高的特點而被人們所熟知,它是世界上最著名,分布最廣的主導瘦肉型豬,全世界范圍內均有飼養[1-3]。據調查,國內外很多特有小型豬,如廣西巴馬小型豬、西藏小型豬、甘肅蕨麻豬、Yucatan小型豬等,作為實驗動物模型廣泛應用于動物醫學、科學研究、器官移植等諸多方面[4-7];而長白豬,杜洛克和大白豬等為重要的肉用型品種多用于經濟性狀的選育和保種[8],用于SPF級實驗動物模型研究更是鮮有報道。而中國農業科學院哈爾濱獸醫研究所從加拿大引進100頭無豬瘟、豬呼吸綜合征、豬傳染性胃腸炎、豬流感等病原體的SPF豬主要應用于實驗動物,其中大白豬和長白豬各50頭包含32窩大白豬和28窩長白豬。隨豬附贈系譜材料顯示該種群可追溯至其10代以內的血緣關系,引進之后繼續對它們進行封閉環境飼養,已繁育一代。運用微衛星標記技術檢測兩個種群的遺傳多樣性明確其遺傳背景為實驗動物化提供理論依據。現擬向哈爾濱市及周邊多個豬場、科研機構和生物制藥物企業提供標準的實驗動物素材為豬病的研究和疫苗的制備提供良好的實驗動物材料。
微衛星標記(microsatellite),又稱為短串聯重復(short tandem repeats,STRs)或簡單重復序列(simple sequence repeats,SSRs),由2-6個核苷酸串聯重復片段構成,由于重復單位的重復次數在個體和品種間呈高度變異而產生數量豐富的等位基因(或基因型),廣泛應用于動植物優良性狀的選育和標記,如稻米的淀粉品質、豬的主要經濟性狀的選育與標記[9-11]。在家畜育種和親子鑒定(包括個體來源、品種來源、個體或品種間的親緣關系鑒定)[12-14]中有著重要意義。
1 材料與方法
1.1 樣品采集
98份豬耳組織樣本,引自加拿大,均來自中國農業科學院哈爾濱獸醫研究所牧場。其中大白豬和長白豬各49頭,所采耳組織樣品均置于無菌的1.5 mL EP管中帶回實驗室,-20℃冰箱保存備用。
1.2 實驗試劑
2×Taq PCR Starmix聚合酶購自GenStar,小劑量基因組DNA提取試劑盒購自Axygen公司。引物根據聯合國糧農組織(FAO)和國際動物遺傳學會(ISAG)聯合推薦的30對微衛星引物中,依據等位基因數的豐富程度、分布在不同染色體的原則,選取了其中的19個微衛星位點,實際實驗擴增過程中部分位點的退火溫度和擴增后分析范圍進行微調(見表1),由北京金唯智公司合成,其他試劑均為進口或國產分析純。
1.3 基因組DNA的提取
按照Axygen小劑量基因組DNA提取試劑盒說明書提取豬耳組織基因組DNA。將DNA溶于去離子水,用NanoDrop-100紫外分光光度計測定濃度和純度,然后用去離子水稀釋至 20 ng/μL,置于-80℃冰箱凍存備用。
1.4 PCR擴增與檢驗
根據擴增片段的理論值大小,每組位點中的片段大小相差至少10~25 bp,并且標記的熒光顏色不發生重疊,將19對微衛星引物分為6組。PCR反應體系為20μL:上、下游引物各1μL(10 mmol/L),2 ×Taq PCR Starmix聚合酶10μL,模板DNA 1μL,ddH2O 7μL。反應條件為:94℃預變性5 min;94℃變性30 s,55℃~65℃退火(分組及不同引物的Tm值見表1)30 s,72℃延伸30 s,30個循環;最后72℃延伸7 min,10℃保存。擴增產物按組混合加樣進行凝膠毛細管電泳檢驗,并用GeneMapper V4.0軟件進行熒光數據提取、分子量標準設定和PCR產物片段大小的計算,最后根據核心堿基數完成基因型的判定。
1.5 統計分析
采用Excel Microsatellite Toolkit V3.1軟件,計算大白豬和長白豬在19個微衛星位點上的等位基因數、雜合度、多態信息含量。利用Genepop V1.2軟件進行群體間的遺傳分化和基因型差異的分析[15]。利用FSTAT V2.9.3進行F-檢驗[16]。利用Structure V2.3軟件分析大白豬和長白豬的遺傳分化結構圖,所設參數“Burnin Period”和“After Burnin”均為50 000次,K值為2-5,然后利用distructure軟件顯示圖形。
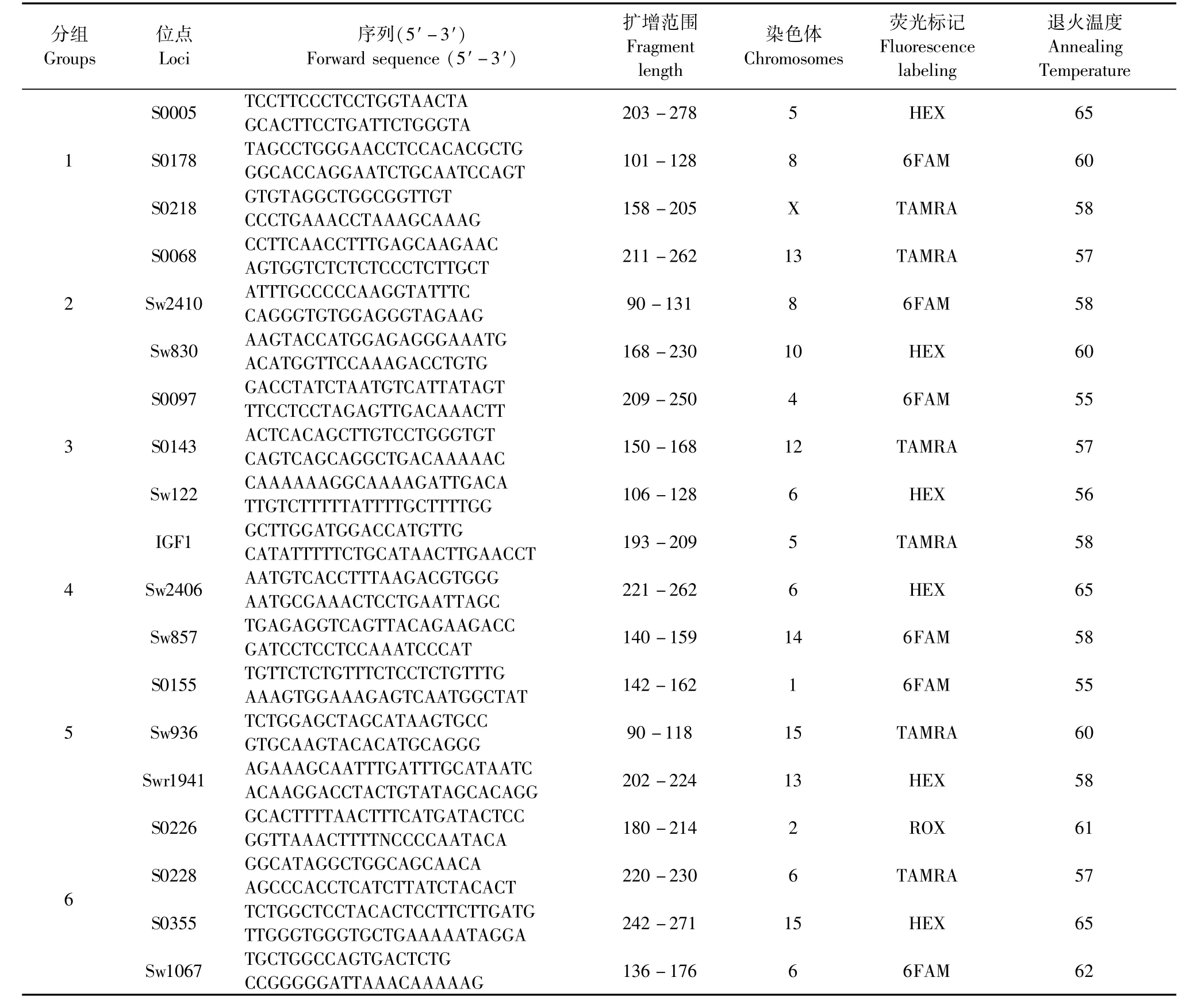
表1 19對微衛星引物詳細信息Tab.1 Detailed information of the 19 microsatellite loci primers
2 結果
2.1 擴增結果
PCR產物經2%的瓊脂糖凝膠檢測,擴增片段大小與預期結果相符(圖1)。大白豬和長白豬在19個微衛星位點分別檢測到84和89個等位基因,兩群體共享57個等位基因,平均含有4.42和4.63個等位基因(表2)。19個位點中S0005位點等位基因最多有14個,Sw2406和S0218位點等位基因最少各3個(表3)。
2.2 等位基因和雜合度分析
19個微衛星位點的平均雜合度相對較高,其中長白豬略高于大白豬分別為0.6066和0.5877,均在0.5~0.7之間(表2)。大白豬群中S0005位點的雜合度最高為0.8395檢測到雜合體43個,S0178位點的雜合度最低為 0.1515僅檢測到 8個。Sw2406位點共檢測到221、233、251 bp 3個等位基因,而251 bp等位基因僅存在于大白豬群中,在長白豬群中并沒有檢測到該等位基因,表明251 bp等位基因為大白豬特有的等位基因;長白豬S0178位點的雜合度最高為0.7570,S0218位點的雜合度最低為0.0204,在S0218位點僅檢測到167 bp和169 bp兩個等位基因而169 bp等位基因在長白豬群中只檢測到一個。此外,Sw857 144 bp、Sw936 117 bp、S0143 168 bp、S0155 152 bp和S0178 109 bp這5個位點在兩群體間的分布和含量差異極顯著。結果表明相同位點的等位基因個數和雜合度在不同群體差異極顯著。
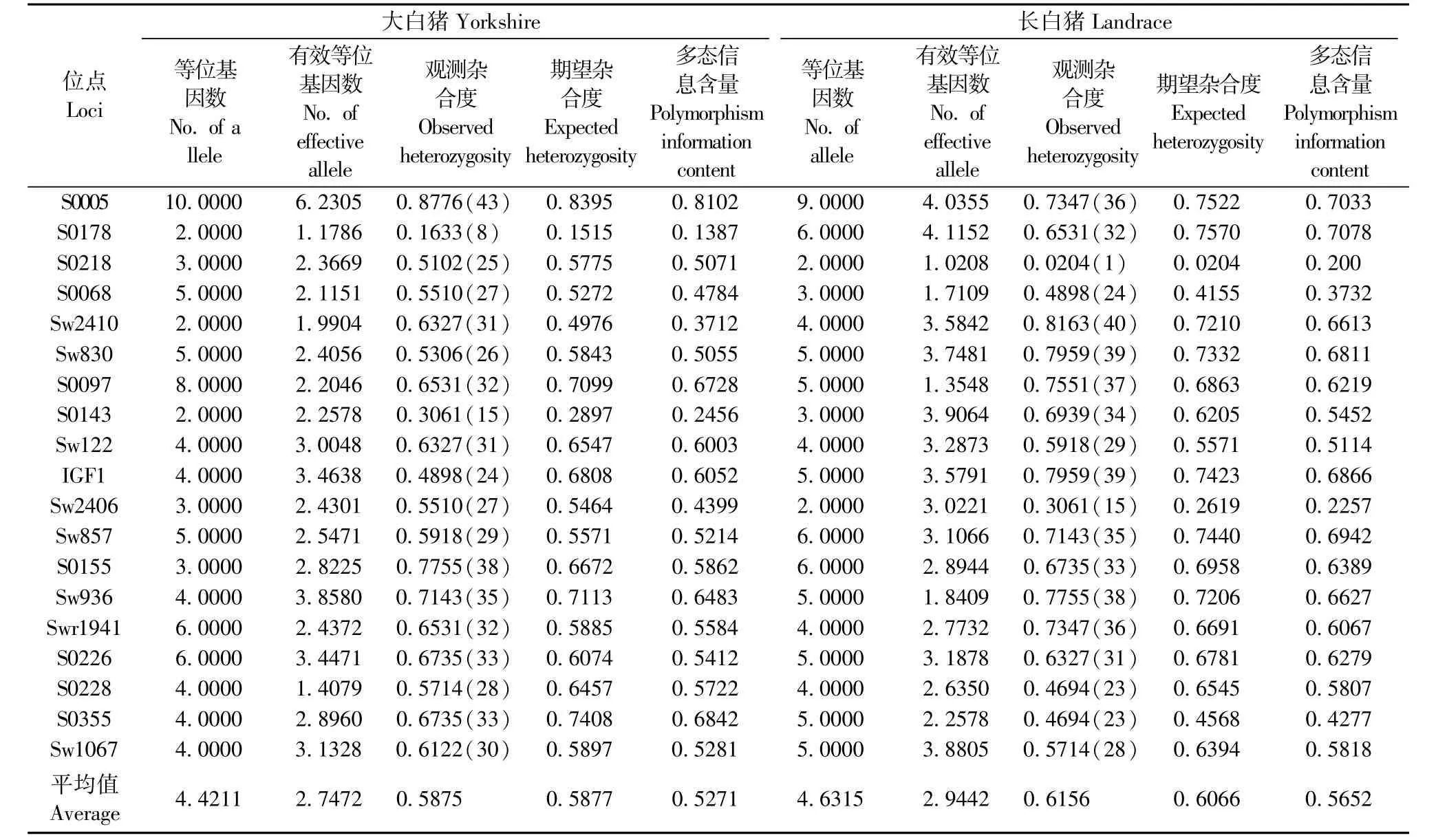
表2 大白豬和長白豬兩個群體在19個微衛星位點上的雜合度和多態信息含量Tab.2 Value of heterozygosities and polymorphism information content(PIC)in the pedigree Yorkshire and Landrace populations at19 microsatellite loci
圖1 S0143位點擴增產物Fig.1 PCR amplification of the products of S0143 in agarose
2.3 多態信息含量
兩個群體中S0005位點的多態信息含量最高為0.8102,S0178位點多態信息含量最低0.1387,大白豬和長白豬的平均多態信息含量分別為0.5271和0.5652。S0005、S0218、S0068、Sw2410、S0097、Sw122、Sw2406、S0355位點的多態信息含量在大白豬群普遍高于長白豬群,而其他11個位點則反之(表2)。19個微衛星位點中S0005位點的多態性信息含量最高為0.8696,Sw2406位點多態信息含量最低為0.3328,平均多態信息含量為0.6388(表3)。結果表明大白豬和長白豬兩個群體間的平均多態信息含量差異無顯著性(P>0.05),沒有表現直接的線性關系。
2.4 F-統計和遺傳結構分析
通過微衛星座位的F-統計以及Nm來檢驗群體間的遺傳分化程度和遷移率(表3)。群體的固定指數Fst為0.0273-0.5218,均值為0.2145,而Fis和Fit指數有正有負,如 S0178、S0218、IGF1、S0228、S0355、Sw1067位點的Fis值均為正值,而Fit除了SOO68為負值外其他18個位點均為正值。Fis和Fit出現正值表明群體基因型分化顯著。Nm為0.2291-8.9075,平均值為1.5320。Nm越小表明群體的遺傳結構越相似。群體遺傳結構分析表明,兩豬群遺傳分化結構穩定(圖2),一方面可能因為它們是純系品種,另一方面則主要是因為兩豬群在19個位點的等位基因較少,共享極為有限的等位基因。
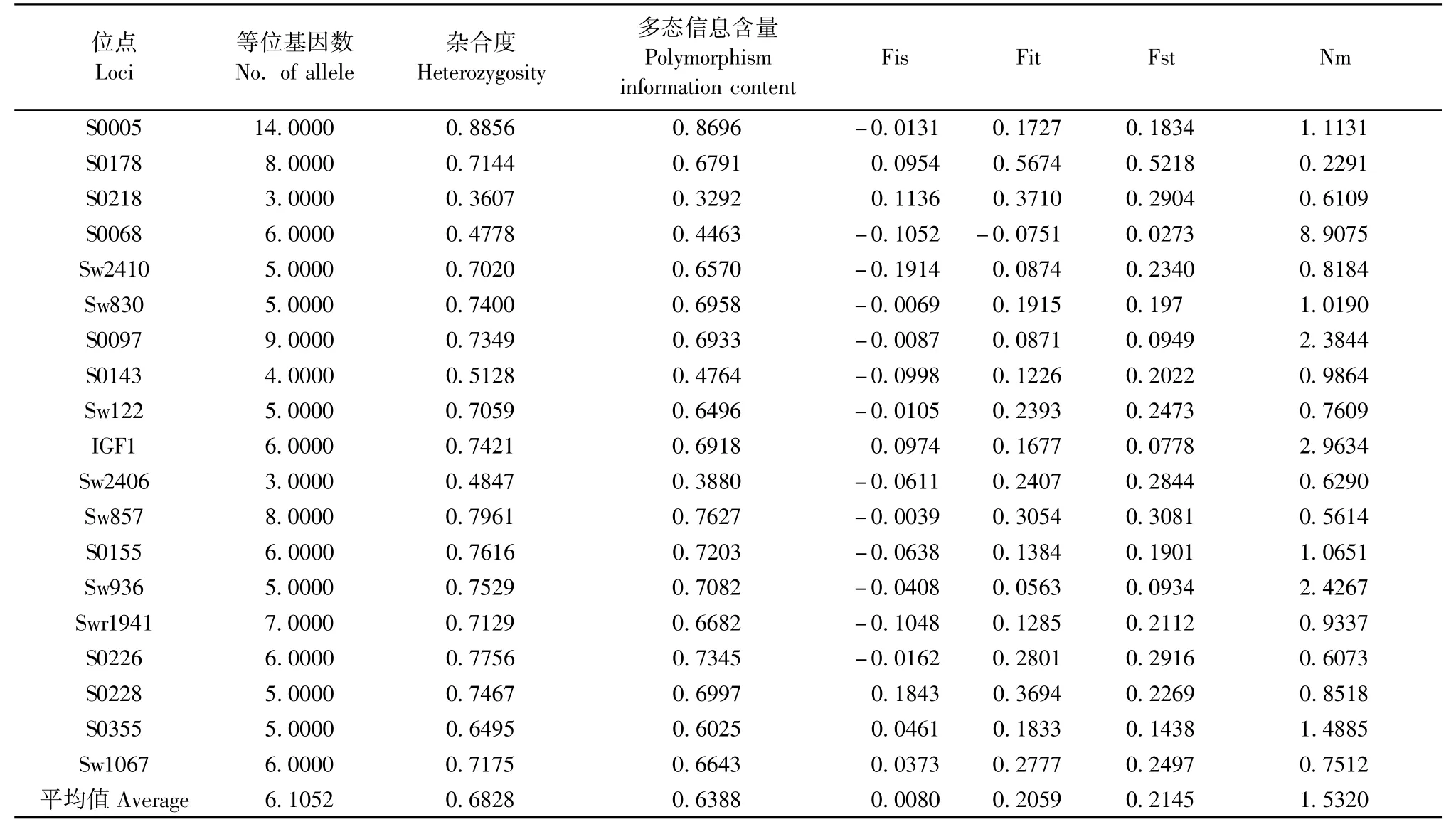
表3 19個微衛星座位的雜合度,多態信息含量和F-統計Tab.3 The F-statistics and polymorphism information content(PIC)of the 19 microsatellite loci
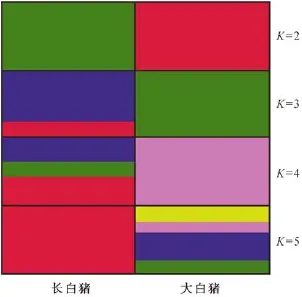
圖2 大白豬和長白豬的遺傳結構Fig.2 Genetic structure of Yorkshire and Landrace pigs
3 討論
3.1 群體的多樣性
微衛星標記以其分析方法便捷、操作簡單等優點,在親子鑒定、輔助標記和群體遺傳多樣性等方面得到廣泛應用。研究表明19個微衛星位點之間的多樣性差異存在顯著性(P>0.05),主要表現在相同位點的等位基因頻率在大白豬和長白豬群體中的分布差異有顯著性(P>0.05),如在S0178位點109 bp的等位基因在大白豬種群出現的頻率最高,而長白豬種群在109 bp處等位基因僅檢測到3個。其中19個位點特異性等位基因出現在長白豬群的頻率較高。如S0178位點處121、123、125 bp等位基因,Sw936位點處117 bp等位基因,S0228位點處230和234 bp等位基因和S0155位點處152 bp等位基因僅在長白豬種群中被檢測到,而Sw857位點處144 bp和S0143位點處168 bp等位基因僅在大白豬被檢測到,可能與長白豬具有較高的雜合度有關。可將S0155、S0143、S0178、Sw857和Sw936位點作為長白豬和大白品種鑒定的候選微衛星位點。李何君等[17]和Costa等[18]分別應用12和14對微衛星引物對美系、英系、法系大約克夏豬群體和歐洲野豬進行品系鑒定及個體鑒定,鑒定正確率均為100%,本研究選取的5對候選品種鑒定引物均在兩者的研究之列,說明該位點是準確、有效、可靠的品系鑒定位點。此外,分別對大白豬和長白豬群體的觀測雜合度和期望雜合度進行卡方檢驗,表明兩群體差異無顯著性(P>0.05),符合封閉群條件(參照實驗用小型豬遺傳質量控制,北京地方標準: DB11/T 828.3 2011)。可從分子水平明確兩種群的遺傳背景,反映出大白豬和長白豬兩群體之間的遺傳差異并作為“活的儀器”廣泛應用于病毒的感染實驗,疫苗的檢定和評價,為遺傳保種和實驗動物化提供良好的分子基礎。
遺傳多樣性的衡量指標通常為群體的平均雜合度和多態信息含量,大白豬的平均多態信息含量和平均雜合度分別為0.5271和0.5877;長白豬的平均多態信息含量和平均雜合度分別為0.5652和0.6066。大白豬和長白豬兩個品種的平均雜合度與平均多態信息含量與趙振華等[19]和鞏元芳等[20]對國內大白豬和長白豬的研究結果相比較低,表明該種群與國內純系品種相比人工選育的程度更高,遺傳結構更趨穩定。此外,19個微衛星座位的平均雜合度比Li等[21]對臺灣歐洲純系品種豬的研究結果略高,表明它們的選育程度非常接近,遺傳結構比較穩定,更適合選用于標準的實驗動物材料。
3.2 群體的遺傳分化與變異
Fis值和Fit值均有正有負,表明群體的等位基因和基因型分化顯著,主要表現為相同位點的等位基因在不同品種間的分布差異顯著(P>0.01)。而Fst值表示群體間的分化程度,Fst的平均值為0.2145與趙振華等[19]和楊彤彤等[22]相比此次引進種群的分化程度顯著(P>0.01)低于國內部分純種豬群[15]。Nm值越小表示群體間的遺傳結構越相似,Nm的平均值為1.5320也顯著(P>0.01)低于楊彤彤等[22]對我國部分純系豬的研究。與國內部分保存的純系豬相比又一次證明了中國農業科學院哈爾濱獸醫研究所引進的大白豬和長白豬兩純系種群群體內分化較小遺傳結構穩定適合用于實驗動物材料,這可能與該種群的長期人工選育有關。
[1] Molina RM,Cha SH,Chittick S,etal.Immune response against porcine reproductive and respiratory syndrome virus during acute and chronic infection[J].Vet Immunol Immunopathol,2008,126:283-292.
[2] Drew TW.The emergence and evolution of swine viral diseases: to what extent have husbandry systems and global trade contributed to their distribution and diversity?[J].Rev SciTech,2011,30(1):95-106.
[3] Lunney JK,Ho CS,WysockiM,et al.Molecular geneticsof the swinemajor histocompatibility complex,the SLA complex[J]. Dev Comp Immunol,2009,33:362-374.
[4] 張德福,戴建軍,吳彩鳳,等.小型豬實驗動物化研究現狀及其應用前景[J].豬業科學,2009,3:74-76.
[5] Smith DM,Martens GW,Ho CS,et al.DNA sequence based typing of swine leukocyte antigens in Yucatan miniature pigs [J].Xenotransplantation,2005,12:481-488.
[6] 徐玲玲,岳敏,吳艷,等.花西藏小型豬群體遺傳結構分析[J].中國比較醫學雜志,2009,19(2):26-31.
[7] Wang C,Xu LL,Wu YH,et al.Selected representativemicrosatellite loci for geneticmonitoring and population structure analysis ofminiature swine[J].GenetMol Res,2015,14(2):3910 -3920.
[8] 潘晚平,申祥科,邱漢華,等.美系長白豬“大白豬”杜洛克豬新品系的選育研究[J].養豬,2003,5:23-24.
[9] 包勁松,舒慶堯,吳殿星,等.水稻Wx基因(CT)n微衛星標記與稻米淀粉品質的關系研究[J].農業生物技術學報,2000,8(3):241-244.
[10] 樊斌,彭中鎮,李奎,等.微衛星標記及其在豬遺傳育種中的應用[J].河北農業大學學報,1999,33(2):161-164.
[11] 劉永祥,劉艷麗,朱寬佑,等.小尾寒羊微衛星標記的多態性及其與繁殖性能的相關性研究 [J].安徽農業科學,2008,36(6):2233-2234.
[12] 吳繼法,吳登俊.微衛星DNA在家畜親子鑒定中的應用及研究進展[J].國外畜牧科技,2001,28(5):28-30.
[13] 錢林東,張自芳,田應華,等.利用微衛星DNA標記進行黃牛的親子鑒定[J].云南農業大學學報,2010,25(1):69-73.
[14] 張春雷,佟廣香,匡友誼,等.哲羅魚微衛星親子鑒定的應用[J].動物學研究,2010,31(4):395-400.
[15] Raymond M,Rousset F.GENEPOP(version 1.2):population genetics software for exact tests and ecumenicism[J].Heredity,1995,86:248-249.
[16] Weir BS,Cockerham CC.Estimating F-statistics for the analysis of population structure[J].Evolution,1984,38:1358-1370.
[17] 李何君,劉煒,吳昊旻,等.12個微衛星標記在大約克夏種豬品系鑒定中的應用研究 [J].上海畜牧獸醫通訊,2014,6:7-9.
[18] Costa V,Pérez-González J,Santos P,etal.Microsatellitemarkers for identification and parentage analysis in the European wild boar(Sus scrofa)[J].BMC Res Notes,2012,5:479-485.
[19] 趙振華,賈青,墨鋒濤,等.杜洛克、長白和大約克夏純種豬微衛星多態性分析[J].江蘇農業學報,2007,23(2):103 -108.
[20] 鞏元芳,段玲欣,倪靜,等.長白豬微衛星標記的遺傳多樣性[J].河北科技師范學院學報,2012,4(4):1672-7983.
[21] Li KY,Li KT,Cheng CC,et al.A genetic analysis of Taoyuan pig and its phylogenetic relationship to Eurasian pig breeds[J]. Asian-Australas Anim Sci,2015,28(4):457-466.
[22] 楊彤彤,趙族,張婷婷,等.大約克和二花臉豬5個微衛星座位多態性分析[J].畜牧與獸醫,2008,40(9):34-37.
Analjsis of the pedigreed population genetics of SPF Yorkshire and Landrace pigs im ported from Canada
HE Xi-wen1,2,GAO Cai-xia2,JIANG Qian2,QUAN Jin-qiang1,CAIYuan1*,QU Lian-dong2*
(1.College of Animal Science and Technology of Gansu Agriculture University,Lanzhou 730070,China. 2.State Key Laboratory of Veterinary Biotechnology,Harbin Veterinary Research Institute,Chinese Academy of Agricultural Sciences(CAAS),Harbin 150001)
ObjectiveTo understand the genetic background of the specific pathogen-free Yorkshire and Landrace pigs,imported from Canada by Harbin Veterinary Research Institute,Chinese Academy of Agricultural Sciences.MethodsThe population genetics were investigated by using 19 pairs ofmicrosatellite primers.ResultsIn the Yorkshire pigs and Landrace pigs,84 and 89 alleles,respectively,were detected at19 microsatellites loci.The average polymorphic information content and mean heterozygosity in the Yorkshire pigs were 0.5271 and 0.5877,and in the Landrace population were 0.5652 and 0.6066,respectively.Because of the significant(P<0.01)differences of alleles in different loci such as S0155,S0143,S0178,Sw857 and Sw936,itmade them possible to be used to identify Yorkshire and Landrace pigs'breed. F-statistics showed that the differentiation within the population was small and genetic structure was stable.ConclusionsCompare with the domestic pedigree largeWhite and Landrace pigs,the SPF pedigree pigs imported from Canada aremore stable in genetic structure,and can be used as laboratory animalmodels in animal science research.
Specific pathogen-free pigs;Microsatellite;Laboratory Animal;Genetic diversity
Q95-33
A
1005-4847(2015)06-0551-06
10.3969/j.issn.1005-4847.2015.06.001
2015-09-28
國際科技合作計劃項目(2010DFB33620);哈爾濱市青年科技創新人才專項資金項目(2013RFQYJ043)。
賀希文(1987-),男,碩士研究生,專業:動物遺傳育種與繁殖,E-mail:hexiwenso@163.com
蔡原,副教授,研究方向:動物遺傳育種與繁殖.E-mail:caiyuan@gsau.edu.cn
曲連東,研究員,研究方向:分子病毒學與動物疫病診斷學.E-mail:qld@hvri.ac.cn
