蒺藜苜蓿葉綠體密碼子偏好性分析
2015-06-01 12:15:17楊國鋒蘇昆龍趙怡然宋智斌孫娟
草業學報 2015年12期
楊國鋒,蘇昆龍,趙怡然,宋智斌,孫娟
(1.青島農業大學經濟草本植物應用研究所,山東 青島 266109;2.青島農業大學生命科學學院,山東省高校植物生物技術重點實驗室,山東 青島266109)
?
蒺藜苜蓿葉綠體密碼子偏好性分析
楊國鋒1,2*,蘇昆龍1,2,趙怡然1,宋智斌1,2,孫娟1
(1.青島農業大學經濟草本植物應用研究所,山東 青島 266109;2.青島農業大學生命科學學院,山東省高校植物生物技術重點實驗室,山東 青島266109)
本文對蒺藜苜蓿葉綠體基因組全序列密碼子進行分析,篩選出50條CDS(coding DNA sequence)利用CodonW軟件進行分析其密碼子使用模式。結果顯示,蒺藜苜蓿葉綠體基因組密碼子第3位堿基GC含量為26.9%,即第3位密碼子富含A和U,ENC值在37.11~51.91之間密碼子偏好性較弱。相對同義密碼子使用度分析顯示RSCU值大于1的密碼子有23個,其中以A和U為結尾20個。中性繪圖分析顯示GC12與GC3的相關系數為0.341,相關性不顯著,回歸系數為0.4843;單基因ENC比值多分布在-0.05~0.05,即大部分基因ENC值離ENC期望值較近;對應性分析,第一軸顯示了12.50%的差異為主要影響因素,第一軸與ENC和GC3的相關系數分別為0.091和-0.092,均相關不顯著。綜合這幾項分析發現蒺藜苜蓿葉綠體基因組密碼子偏好性主要受到突變的影響,但是并不是唯一的影響因素,其他因素對密碼子偏好性也可能有一定的影響。最終通過高表達優越密碼子方法確定得出UUA、UUG、CCU等23個密碼子為最優密碼子,為之后對外源基因進行改造,提高其在葉綠體中的表達效率奠定了基礎。
蒺藜苜蓿;葉綠體;密碼子偏好性;最優密碼子
遺傳信息的傳遞是生物進行生命活動最主要環節,在這一活動中密碼子扮演著重要的角色,編碼不同氨基酸密碼子的準確識別是保證遺傳信息正確表達的關鍵。密碼子具有兼并性,即同一氨基酸能由一個或多個密碼子所對應,這樣的密碼子稱為同義密碼子(synonymous codon)。研究發現,密碼子的使用存在不均等的現象,即有的密碼子使用頻率高于其他同義密碼子的使用,這一現象稱為密碼子偏好性(codon usage bias)。密碼子偏好性是在生物長期進化過程中所形成的,不同的物種間密碼子使用的偏好性不同。由于密碼子偏好性的存在常導致外源基因在宿主中表達量降低[1]。研究密碼子的使用模式對探究物種的進化模式和提高外源基因的表達具有重要意義。
關于密碼子偏好性形成的機制,最早Grantham等[2]提出的“多基因假說”認為在長期的系統發育進化過程中,不同的基因組存在不同的密碼子使用模式,暗示環境的影響(突變壓力)是密碼子偏好性形成的主要影響因素。Ikemura[3]發現高表達基因更傾向于使用一些特定的密碼子,這與tRNA的豐富度相對應,并提出選擇壓力通過優化翻譯效率和精確度來影響同義密碼子的使用頻率。近年來的研究發現,不同物種間密碼子偏好性的差異并非由單一因素決定。Sharp和Li[4]發現密碼子的使用情況與同義替換率有關,Olejniczak和Uhlenbeck[5]通過對單細胞組織進行研究發現密碼子的使用情況與tRNA的豐富度有關系。同時密碼子親水性、DNA復制起止位點[6]、基因長度[7]、翻譯準確性選擇[8-10]和表達水平[11]等因素對密碼子偏好性也產生影響。
與核基因組相比葉綠體基因組密碼子使用模式的研究相對落后些,目前數據庫中完整的葉綠體基因序列數據相對有限,僅有玉米(Zeamays)、水稻(Oryzasativa)、文心蘭(Oncidium)、銀白楊(Populusalba)等植物的完整葉綠體基因組數據。苜蓿(Medicago)為一年生或多年生草本植物,是重要的豆科牧草。隨著基因測序技術的發展,蒺藜苜蓿(Medicagotruncatula)為研究豆科植物的尤其是苜蓿屬的模式植物[12],其葉綠體基因組的測序已經完成,但是關于苜蓿葉綠體密碼子使用模式的研究還未見報道。本研究通過對蒺藜苜蓿葉綠體基因組密碼子使用模式以及影響密碼子使用模式的主要因素進行分析,為以后苜蓿葉綠體的研究和應用奠定基礎。
1 材料與方法
1.1材料
2014年10月從GenBank下載完整的蒺藜苜蓿葉綠體基因組(登錄編號:NC_003119.6)。共下載到不含重復的76條CDS(coding DNA sequence),由于短序列不能正確地計算有效密碼子數[13],研究需去除其中長度小于300 bp的CDS。本研究去除26條長度小于300 bp的CDS,剩余50條CDS用于后面的數據分析。
1.2方法
在分析時去除不編碼任何氨基酸的終止密碼子UAA、UGA和UAG,還有色氨酸的唯一密碼子UGG和蛋氨酸的唯一密碼子AUG,這些密碼子不存在偏好性。使用DNAMAN將50條DNA序列編碼成可在CodonW軟件中運行的.dat格式文件,運行CodonW軟件對基因序列進行分析得到相應數據。對所得到的數據進行整理,使用SPSS及Excel進行下文所述的相關作圖及分析。對序列進行3次重復分析,每次分析均得到相同的實驗結果。
1.2.1相對同義密碼子使用度分析 相對同義密碼子使用度(relative synonymous codon usage, RSCU)表示該密碼子的實際使用值與理論使用值之間的比值[14]。當RSCU<1時,表示該密碼子的使用頻率低于其他同義密碼子;RSCU>1時,則表示該密碼子的使用頻率高于同義密碼子;RSCU=1表示該密碼子沒有偏好性。
1.2.2中性繪圖分析 統計密碼子3個位置的GC含量,第1,2,3位的GC含量分別表示為GC1、GC2、GC3,GC12表示GC1和GC2的平均值。中性繪圖分析以GC12為縱坐標,以GC3為橫坐標作圖。通過分析密碼子的第1,2位和第3位堿基組成的相關性,研究密碼子的偏好性影響因素。當GC12與GC3之間顯著相關,說明3個位置上的堿基組成無差異,密碼子的使用受突變的影響。當GC12與GC3相關性不顯著,回歸系數接近0,說明第1,2位和第3位堿基組成不同,基因組GC含量高度保守,密碼子的使用更多地受選擇影響[15]。
1.2.3ENC-plot繪圖分析 有效密碼子數(effective number of codon,ENC)反映的是密碼子偏離隨機選擇的程度[16],是反映同義密碼子非均衡使用偏好程度的重要指標,通常高表達基因其密碼子偏好程度較大,因此ENC取值較小;低表達基因含有較多種類的稀有密碼子偏好性較弱,ENC值也較大。ENC-plot繪圖分析以ENC為縱坐標,GC3為橫坐標作圖。該分析主要檢測堿基組成對密碼子偏好性的影響,繪圖中的標準曲線表示密碼子偏好性僅由堿基組成決定基因位置,標準曲線計算方法如下:
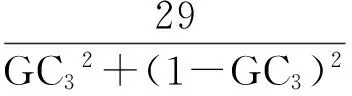
基因沿標準曲線分布或落在標準曲線附近表示該基因密碼子偏好性僅受突變影響,基因落在標準曲線下方較遠的位置表示該基因密碼子偏好性受到選擇的影響。
1.2.4PR2-plot繪圖分析 PR2偏倚分析(PR2-bias plot analysis)是為了避免密碼子第3位堿基A與T和C與G之間突變不平衡,根據偏倚規則(parity rule 2,PR2),如果兩條互補鏈間不存在任何突變或選擇效應上的偏倚,那么堿基含量上應該有A=T和C=G。對由4個同義密碼子編碼的氨基酸的各個密碼子第3位A、T、C、G的情況進行分析,計算每個基因A3/(A3+T3)和G3/(G3+C3),分別作縱坐標和橫坐標作圖,圖中中心點表示A=T且C=G,其余的點由中心點向該點發出的矢量表示了該基因的偏倚程度和方向[17]。
1.2.5對應性分析 對應性分析(correspondence analysis)是一種多元相依的變量統計分析技術,通過分析由定性變量構成的交互匯總表來揭示變量間的聯系,可以揭示同一變量的各個類別之間的差異,以及不同變量各個類別之間的對應關系。為探究不同基因在密碼子用法上的變異趨勢,基于RSCU值進行對應性分析,CodonW軟件會將樣本中所有基因分布到一個58維(58個同義密碼子)的向量空間中,其中每個點就代表一個同義密碼子。密碼子的用法特點通過圖中每個點之間的位置體現出來。通過探究基因向量及基因間的值的變異情況,推測出密碼子偏好性的主要影響因素。第一軸表現最大差異的密碼子使用變化,隨后的副軸變化量逐漸降低。McInerney[18]指出,兩個主軸間基因的分散代表了主要的密碼子變化趨勢。將表現差異最大的第一軸和第二軸分別作為橫縱坐標作圖,圖中點代表不同基因,根據圖中點的分布情況判斷基因密碼子的使用模式。
1.2.6最優密碼子的確定 最優密碼子(the most preferred codon)的選擇,以ENC為偏好性標準,兩極各選10%的基因,分別建成高低偏性庫,取兩庫ΔRSCU>0.08的密碼子[19]。
本研究所使用的軟件包括DNAMAN 6.0、CodonW 1.4.2、SPSS 19和Excel 2013。
2 結果與分析
2.1密碼子組成分析
使用CodonW軟件對各基因編碼序列進行分析(表1)。原則上ENC的取值范圍為20~61,ENC值大小反映的密碼子偏性的強弱,當ENC為20時,表示同義密碼子完全偏倚;為61時,表示同義密碼子沒有偏倚;按照慣例以35作為偏性強弱的區分標準[18],表中ENC取值范圍在37.11~51.91之間,并且大部分大于45,所以蒺藜苜蓿葉綠體基因密碼子偏性較弱。密碼子第3位GC的平均含量為26.9%,明顯低于前兩位45.5%和36.8%的GC含量,說明密碼子第3位堿基組成多為A和U。
GC1、GC2、GC3、GCall、ENC和密碼子數目N之間的關聯分析見表2。GCall與GC1、GC2、GC3之間的相關性均達到了極顯著水平,GC1與GC2顯著相關,GC3與GC1、GC2均未達到顯著水平,說明密碼子的第1位與第2位的堿基組成比較相似,但是與第3位不同。ENC與密碼子第1、第2位置GC含量相關但是相關性不顯著,與密碼子第3位的GC含量顯著相關。ENC與密碼子數目N之間未達到顯著水平,說明密碼子數目對ENC的影響很弱,即排除了基因序列過短對密碼子偏好性的影響。
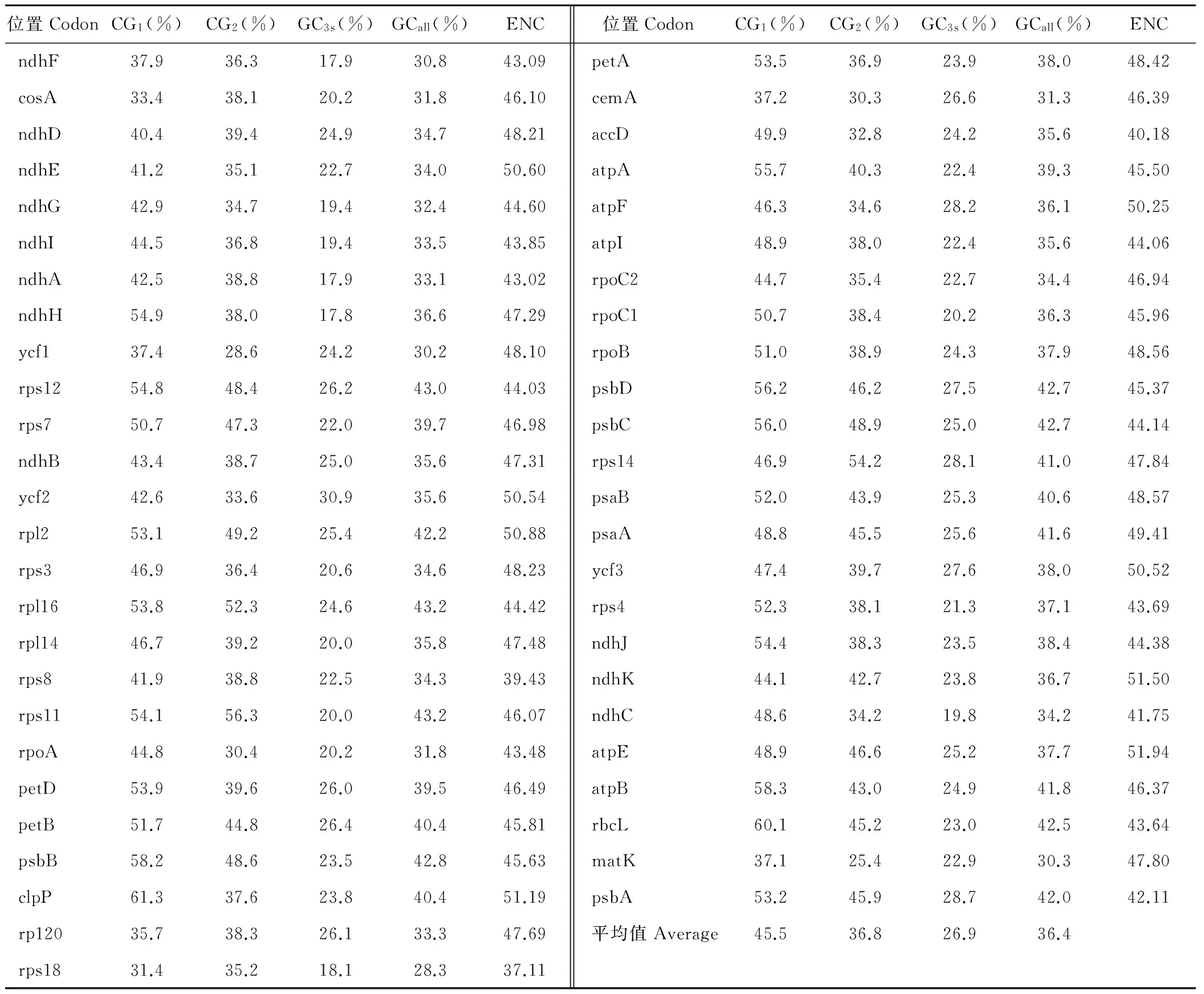
表1 蒺藜苜蓿葉綠體基因組不同位置的GC含量Table 1 GC content of different positions of codon in chloroplast of M.truncatula
各種氨基酸的RSCU值分析見表3。從表中可以看出,RSCU值大于1的主要是以U和A為結尾的密碼子,密碼子以U和A為結尾的出現頻率比較高,說明這些為基因組偏愛密碼子。相反以C和G為結尾的密碼子出現頻率比較低,這些是基因組的非偏愛密碼子。
2.2中性繪圖分析
苜蓿葉綠體基因組各基因中性繪圖分析見圖1,可以看出GC12的取值范圍在0.178~0.309之間,GC3取值范圍在0.321~0.552之間。圖中各基因未落在對角線上或沿對角線分布,都在對角線上方分布。GC12與GC3的相關系數為0.341,相關性不顯著,回歸系數(即圖中斜率)為0.4843,說明第一、二位和第三位堿基組成無差異,蒺藜苜蓿葉綠體基因組密碼子的使用更多地受突變的影響。
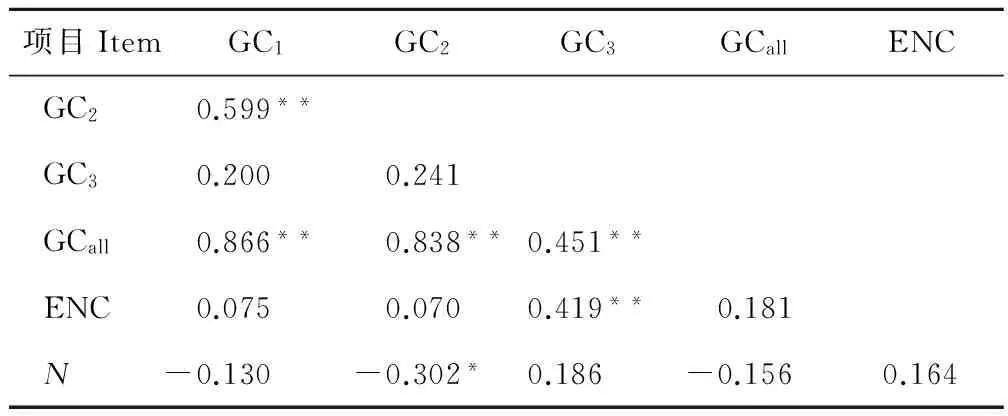
表2 各基因相關性分析Table 2 Correlation analysis of each genes related parameters
** 在0.01水平上顯著相關;* 在0.05水平上顯著相關。
** Significant correlation atP<0.01; *Significant correlation atP<0.05.

表3 蒺藜苜蓿各氨基酸相對同義密碼子使用度Table 3 RSCU analysis of protein coding region in M.truncatula
2.3ENC-plot繪圖分析
ENC與GC3的關聯分析見圖2,由圖可以看出部分基因位點沿著標準曲線分布或者落在標準曲線附近,這些基因的實際ENC值與預期ENC值接近,部分基因位點落在期望值的下方。為了更直接地顯示實際ENC值和預期ENC值之間的差異,計算了ENC比值即(預期ENC值-實際ENC值)/預期ENC值,統計ENC比值頻數,從而顯示出有多少基因距離期望值更近。從表4中可以看出大多數的ENC比值分布在-0.05~0.05之間,這一范圍內實際ENC值與預期ENC值相差較小,因此蒺藜苜蓿葉綠體基因組密碼子偏好性與GC3的差異相關。所以蒺藜苜蓿葉綠體基因組密碼子偏好性更多地受突變的影響,受選擇地影響相對較弱一些。
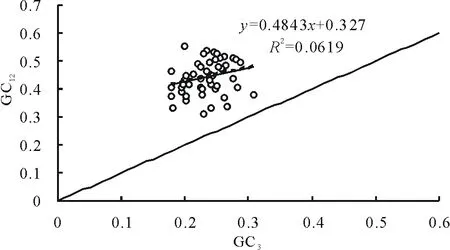
圖1 中性繪圖分析
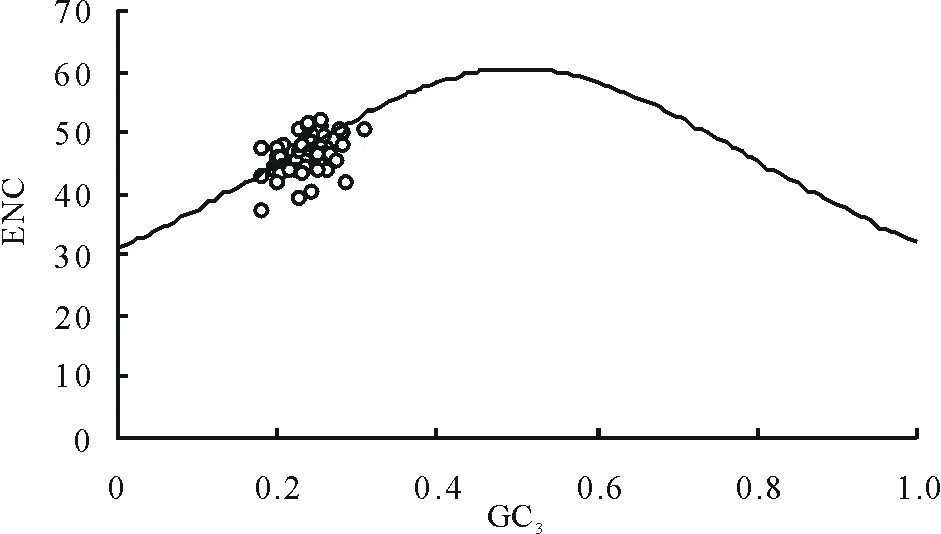
圖2 ENC-plot繪圖分析
2.4PR2-plot繪圖分析
通過PR2-plot繪圖分析了各基因部分氨基酸的嘌呤(A和G)嘧啶(T和C)的關系。由圖3可以看出大部分基因分布在平面圖的右下方,說明T的使用頻率高于A,G的使用頻率高于C。若完全受到突變的影響,A和T以及G和C的使用頻率應當相等,因此蒺藜苜蓿葉綠體基因組密碼子使用模式不僅是受到突變的影響,還受其他因素的影響。
2.5對應性分析
基于RSCU的對應性分析中第1向量軸顯示了12.50%的差異,另外3個軸分別顯示了9.01%,8.89%,8.23% 的差異,前4軸的累積差異38.63%,說明第1軸是影響密碼子葉綠體基因組密碼子偏好性的主要因素。第1軸與ENC和GC3的相關系數分別為0.091和-0.092,均相關不顯著,說明GC3并不是造成蒺藜苜蓿葉綠體基因組密碼子偏好性的主要因素,暗示影響密碼子使用模式的因素并不單一。以第1軸為橫坐標,第2軸為縱坐標,按照基因功能將基因分布于平面上(圖4)。編碼核糖體蛋白基因和遺傳系統基因分布相對比較集中,說明這些基因的密碼子使用模式相近;而其他基因則分布比較分散,說明這些密碼子的使用模式存在差異。
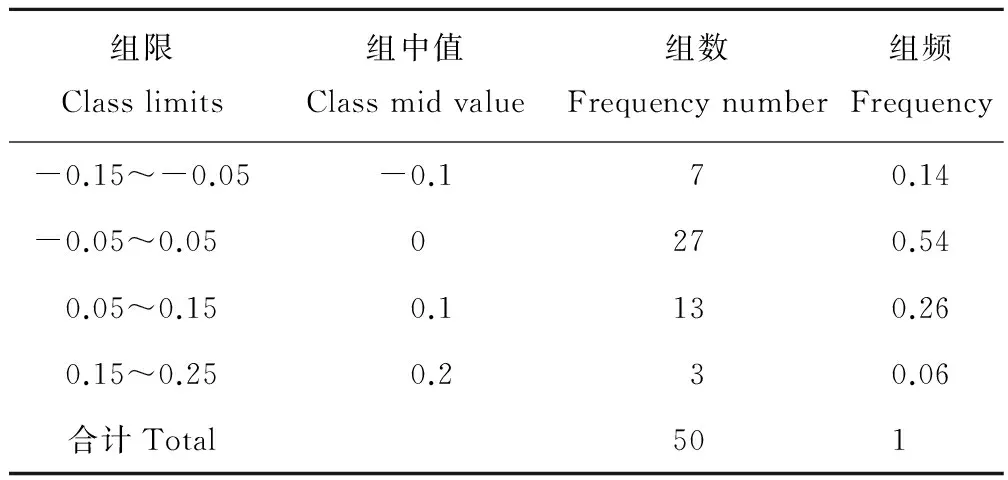
表4 ENC比值頻數分布Table 4 Distribution of ENC ration
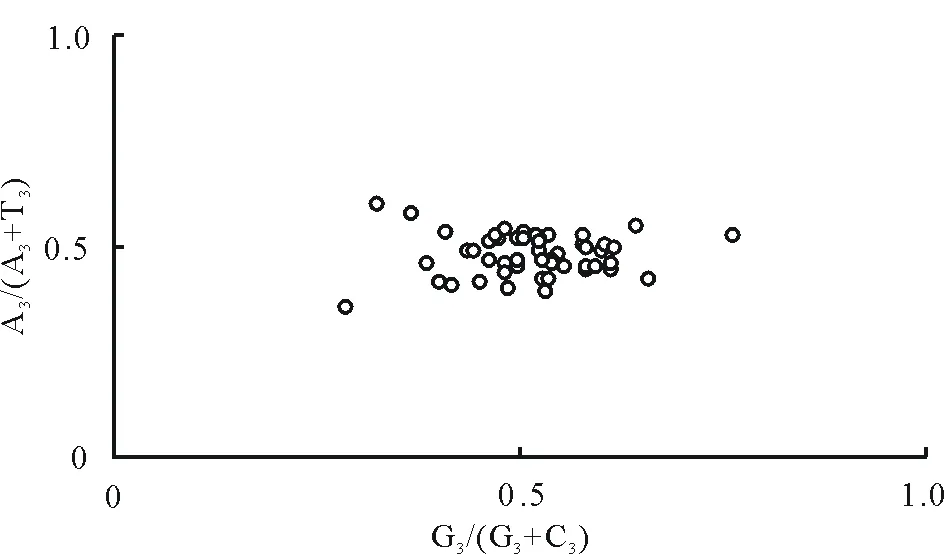
圖3 PR2-plot繪圖分析
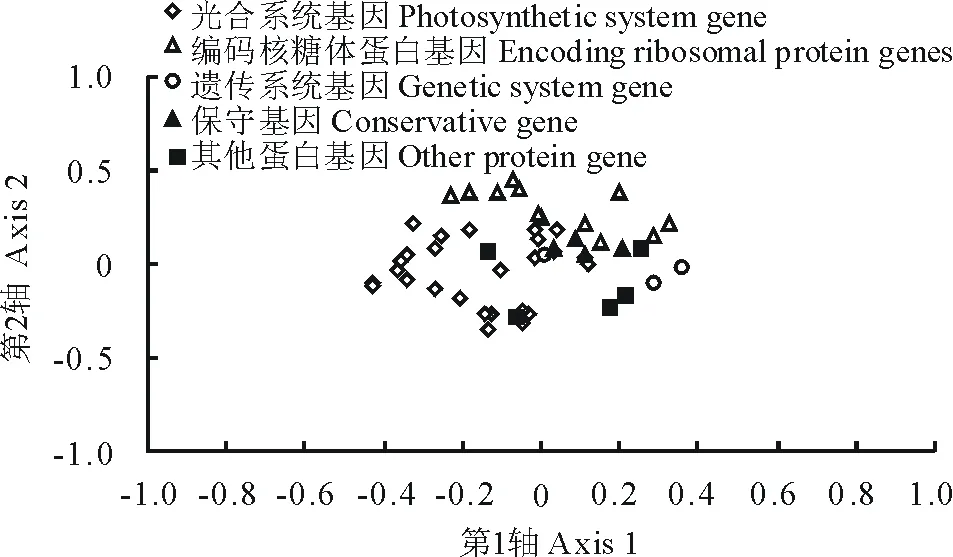
圖4 基于RSCU的對應性分析
2.6最優密碼子分析
按ENC值的大小對各基因進行排序,兩極各選取5個基因構建高表達基因庫和低表達基因庫,并對兩個庫中密碼子分別計算RSCU值,計算ΔRSCU值(表5),表中帶星號的為最優密碼子,其中可以看出ΔRSCU>0.5的均為以A和U為結尾的密碼子。
3 討論
密碼子偏好性是由多個因素共同作用的結果,其中中性選擇影響和方向突變影響是兩個主要因素,哪一個因素為主導因素是目前的關注熱點。密碼子的堿基組成是由核苷酸突變和回復突變的結果,是對密碼子使用最普遍的影響[19-21],尤其是密碼子第3位的改變通常不會引起編碼氨基酸的改變,第3位上的堿基突變受到的選擇壓力比較小,因此研究第3位堿基的堿基組成對研究密碼子偏好性有重要意義。通過對第3位密碼子的GC含量計算發現,第3位密碼子富含A和T。本研究通過對蒺藜苜蓿葉綠體基因組中的基因部分氨基酸的密碼子進行PR2-plot繪圖分析發現,密碼子第3位堿基T的使用頻率高于A,G的使用頻率高于C。尚明照等[22]以及Zhang等[23]分別對陸地棉(Gossypiumhirsutum)和小麥(Triticumaestivum)葉綠體基因組進行研究,發現密碼子第3位堿基T的使用頻率高于A,C的使用頻率略高于G;張月榮[24]對23種禾本科植物葉綠體密碼子進行分析發現密碼子第3位堿基C和G使用頻率相當,T的使用頻率高于A。這表明不同植物的葉綠體基因組的堿基組成存在一定的差異,暗示影響密碼子偏好性的因素也不相同。
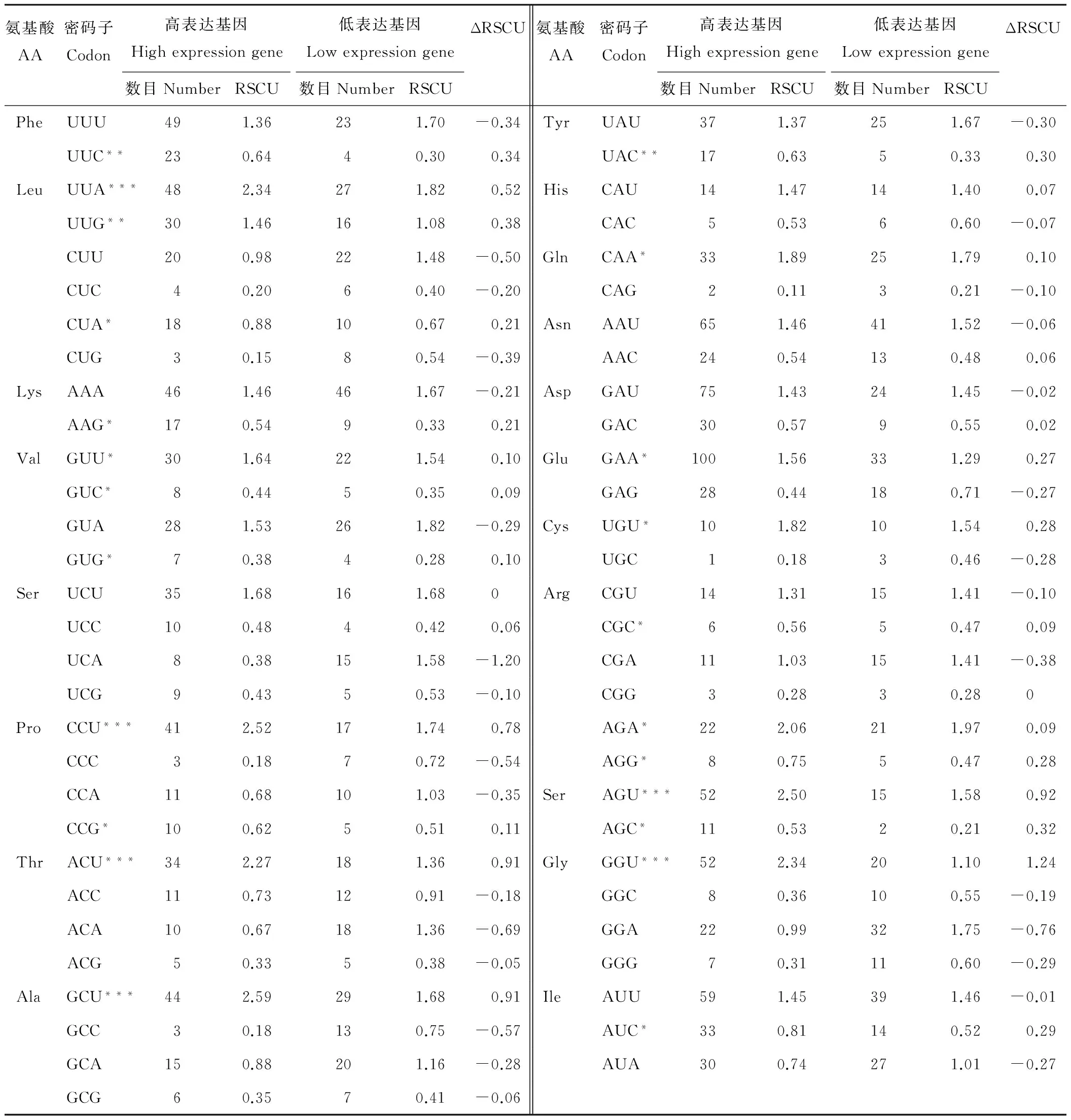
表5 蒺藜苜蓿葉綠體基因組最優密碼子確定Table 5 Preferred codons in chloroplast genome of M.truncatula
*表示ΔRSCU>0.08,**表示ΔRSCU>0.3,***表示ΔRSCU>0.5。
* mean ΔRSCU>0.08,** mean ΔRSCU>0.3,*** mean ΔRSCU>0.5.
結合中性繪圖分析、ENC-plot繪圖分析以及對應性分析,發現蒺藜苜蓿葉綠體基因組密碼子偏好性主要受到突變的影響,但是并不是唯一的影響因素,其他因素也對密碼子偏好性造成影響。Zhou等[25]在對擬南芥(Arabidopsisthaliana)、楊樹等植物的研究中指出,葉綠體基因組密碼子偏好性主要受到突變影響,其他因素也對其產生影響,本研究結果與之相同。但是續晨等[26]對蝴蝶蘭(Phalaenopsisaphrodite)葉綠體密碼子偏好性進行研究,發現其密碼子偏好性主要受到堿基差異和自然選擇的影響。因此可以看出不同植物密碼子偏好性的影響因素不同,但是影響植物密碼子偏好性的因素都不是單一的,是多因素綜合的結果。
本研究采用高表達優越密碼子方法確定最優密碼子,選出了23個最優密碼子,確定的最優密碼子中第3位堿基為A、U、C、G的分別5,7,6,5個,但是ΔRSCU>0.5的均以A和U為結尾。這與前人對水稻[27]、銀白楊[28]等植物的研究結果中葉綠體基因組中密碼子偏好NNA、NNU型相符合。
4 結論
蒺藜苜蓿葉綠體基因組密碼子偏好性受突變的影響比較大,但與此同時也受到其他因素的影響,密碼子偏好性是多因素影響的結果。確定了23個最優密碼子,為以后通過外源基因密碼子改造后轉入苜蓿葉綠體中提高其表達量[29]。
[1] Wu X M, Wu S F, Ren D M,etal.The analysis method and progress in the study of codon bias.Hereditas, 2007, 29(4):420-426.
[2] Grantham R, Gautier C, Gouy M.Codon frequencies in 119 individual genes confirm consistent choices of degenerate bases according to genome type.Nucleic Acids Research, 1980, 8(9):1893-1912.
[3] Ikemura T.Codon usage and tRNA content in unicellular and multicellular organisms.Molecular Biology and Evolution, 1985, 2(1):13-34.
[4] Sharp P M, Li W H.The rate of synonymous substitution in enterobacterial genes is inversely related to codon usage bias.Molecular Biology and Evolution, 1987, 4(3):222-230.
[5] Olejniczak M, Uhlenbeck O C.tRNA residues that have coevolved with their anticodon to ensure uniform and accurate codon recognition.Biochimie, 2006, 88(8):943-950.
[6] Deschavanne P, Filipski J.Correlation of GC content with replication timing and repair mechanisms in weakly expressedE.coligenes.Nucleic Acids Research, 1995, 23(8):1350-1353.
[7] Sun Z, Ma L, Murphy R,etal.Analysis of codon usage onWolbachiapipientiswMel genome.Science in China Series C:Life Sciences, 2009, 39(10):948-953.
[8] Xia X H.Mutation and selection on the anticodon of tRNA genes in vertebrate mitochondrial genomes.Gene, 2005, 345(1):13-20.
[9] Zalucki Y M, Power P M, Jennings M P.Selection for efficient translation initiation biases codon usage at second amino acid position in secretory proteins.Nucleic Acids Research, 2007, 35(17):5748-5754.
[10] Huang Y, Koonin E V, Lipman D J,etal.Selection for minimization of translational frame shifting errors as a factor in the evolution of codon usage.Nucleic Acids Research, 2009, 37(20):6799-6810.
[11] Hiraoka Y, Kawamata K, Haraguchi T,etal.Codon usage bias is correlated with gene expression levels in the fission yeastSchizosaccharomycespombe.Genes to Cells, 2009, 14(4):499-509.
[12] Zhang J, Song L L, Guo D L,etal.Genome-wide identification and investigation of the MADS-box gene family inMedicagotruncatula.Acta Prataculturae Sinica, 2014, 23(6):233-241.
[13] Rosenberg M S, Subramanian S, Kumar S.Patterns of transitional mutation biases within and among mammalian genomes.Molecular Biology and Evolution, 2003, 20(6):988-993.
[14] Sharp P M, Li W H.The codon adaptation index-a measure of directional synonymous codon usage bias, and its potential applications.Nucleic Acids Research, 1987, 15(3):1281-1295.
[15] Sueoka N.Directional mutation pressure and neutral molecular evolution.Proceedings of the National Academy of Sciences of the United States of America, 1988, 85(8):2653-2657.
[16] Wright F.The “effective number of codons” used in a gene.Gene, 1990, 87(1):23-29.
[17] Sueoka N.Near homogeneity of PR2-Bias fingerprints in the human genome and their implications in phylogenetic analyses.Journal of Molecular Evolution, 2001, 53(4-5):469-476.
[18] McInerney J O.Replicational and transcriptional selection on codon usage inBorreliaburgdorferi.Proceedings of the National Academy of Sciences of the United States of America, 1998, 95(18):10698-10703.
[19] Yang H J, Liu G S, Zhang S T,etal.Codon usage bias studies and cluster analysis on chloroplastic gene in tobacco.Acta Tabacaria Sinica, 2012, 18(2):37-43.
[20] Jiang Y, Deng F, Wang H L.An extensive analysis on the global codon usage pattern of baculoviruses.Archives of Virology, 2008, 153(12):2273-2282.
[21] Sueoka N, Kawanishi Y.DNA G+C content of the third codon position and codon usage biases of human genes.Gene, 2000, 261(1):53-62.
[22] Shang M Z, Liu F, Hua J P,etal.Analysis on codon usage of chloroplast genome ofGossypiumhirsutum.Scientia Agricultura Sinica, 2011, (2):245-253.
[23] Zhang W J, Zhou J, Li Z F,etal.Comparative analysis of codon usage patterns among mitochondrion, chloroplast and nuclear genes inTriticumaestivumL.Journal of Integrative Plant Biology, 2007, 49(2):246-254.
[24] Zhang Y R.Analysis of Codon Usage Patterns of the Chloroplast Genomes in the Poaceae Family and RNA Editing Sites in the Chloroplast Transcripts ofAgreartinaadenophorum[D].Yangling:Northwest A&F University, 2013.
[25] Zhou M, Long W, Li X.Patterns of synonymous codon usage bias in chloroplast genomes of seed plants.Forest Study of China, 2008, 11(4):235-242.
[26] Xu C, Ben A L, Cai X N.Analysis of synonymous codon usage in chloroplast genome ofPhalaenopsisaphroditesubsp.Formosana.Molecular Plant Breeding, 2011, 8(5):945-950.
[27] Liu Q P, Xue Q Z.Codon usage in the chloroplast genome of rice (OryzasativaL.ssp.japonica).Acta Agronomica Sinica, 2004, 30(12):1220-1224.
[28] Zhou M, Long W, Li X.Analysis of synonymous codon usage in chloroplast genome ofPopulusalba.Journal of Forestry Research, 2008, 19(4):293-297.
[29] Zhang X M, Xin W, Wang S F,etal.Optimizing the codon usage of HIV-1 gag gene according to the codon bias of vaccinia virus improves the gag gene expression.Chinese Journal of Virology, 2005, 21(3):210-216.
參考文獻:
[1] 吳憲明, 吳松鋒, 任大明.等.密碼子偏性的分析方法及相關研究進展.遺傳, 2007, 29(4):420-426.
[7] 孫錚, 馬亮, Murphy R, 等.Wolbachia pipientis wMel 基因組水平上的密碼子使用分析.中國科學C輯:生命科學, 2009, 39(10):948-953.
[12] 張軍, 宋麗莉, 郭東林, 等.MADS-box 基因家族在蒺藜苜蓿的全基因組分析.草業學報, 2014, 23(6):233-241.
[19] 楊惠娟, 劉國順, 張松濤, 等.煙草葉綠體密碼子的偏好性及聚類分析.中國煙草學報, 2012, 18(2):37-43.
[22] 尚明照, 劉方, 華金平, 等.陸地棉葉綠體基因組密碼子使用偏性的分析.中國農業科學, 2011, (2):245-253.
[24] 張月榮.禾本科葉綠體基因組密碼子使用模式及紫莖澤蘭葉綠體RNA編輯分析[D].楊凌:西北農林科技大學, 2013.
[26] 續晨, 賁愛玲, 蔡曉寧.蝴蝶蘭葉綠體基因組密碼子使用的相關分析.分子植物育種, 2010, 8(5):945-950.
[27] 劉慶坡, 薛慶中.粳稻葉綠體基因組的密碼子用法.作物學報, 2004, 30(12):1220-1224.
[29] 張相民, 辛偉, 王世峰, 等.按痘苗病毒優勢密碼子改造HIV-1 gag基因提高表達水平的研究.病毒學報, 2005, 21(3):210-216.
Analysis of codon usage in the chloroplast genome ofMedicagotruncatula
YANG Guo-Feng1,2*, SU Kun-Long1,2, ZHAO Yi-Ran1, SONG Zhi-Bin1,2, SUN Juan1
1.InstituteofEconomicHerbPlants,QingdaoAgriculturalUniversity,Qingdao266109,China; 2.CollegeofLifeSciences,QingdaoAgriculturalUniversity,KeyLabofPlantBiotechnologyinUniversitiesofShandongProvince,Qingdao266109,China
The complete nucleotide sequence of the chloroplast genome ofMedicagotruncatulawas investigated.Fifty CDS (coding DNA sequences) selected from the chloroplast genome sequence ofM.truncatula, were analyzed using CodonW software.The results show that the third codon position was rich in A and U.ENC ranged from 37.1 to 51.9 meaning that the codon bias was weak.There were 23 codons with relative synonymous codon usage greater than 1 and 20 codons ending with A and T.ENC-plot analysis showed that GC3was not correlated with GC12; ENC ratio’s of most genes ranged from -0.05 to 0.05.In the correspondence analysis of the first group of four axes, the first axis showed 10.3% variation.The correlation coefficients for axis 1 between ENC and GC3were 0.091 and -0.092 respectively (not significant).Synonymous codon usage bias was found, mainly due to the effect of mutation pressure, but there were other factors.In addition, analysis of the high expression codons enabled 23 to be affirmed as the “optimal codons” as UAA, UUG, CCU.The results provide evidence for molecular modification of exogenous genes to increase the expression efficiency inM.truncatulachloroplasts.
Medicagotruncatula; chloroplast; codon usage bias; codon usage bias
10.11686/cyxb2015016
http://cyxb.lzu.edu.cn
2015-01-12;改回日期:2015-03-12
現代農業產業技術體系(CARS-35),公益性行業項目(200903060)和國家自然科學基金青年科學基金項目(31200906)資助。
楊國鋒(1977-),男,山東青島人,在讀博士。
*通信作者Corresponding author.E-mail:yanggf@qau.edu.cn
楊國鋒, 蘇昆龍, 趙怡然, 宋智斌, 孫娟.蒺藜苜蓿葉綠體密碼子偏好性分析.草業學報, 2015, 24(12):171-179.
YANG Guo-Feng, SU Kun-Long, ZHAO Yi-Ran, SONG Zhi-Bin, SUN Juan.Analysis of codon usage in the chloroplast genome ofMedicagotruncatula.Acta Prataculturae Sinica, 2015, 24(12):171-179.
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
媽媽寶寶(2017年3期)2017-02-21 01:22:28
山東工業技術(2016年15期)2016-12-01 05:31:22
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
終身教育研究(2014年5期)2014-02-28 01:23:06
