溫度對解除休眠的紫椴種子萌發過程中內源激素的影響
2015-06-12 09:03:38李巖李娟勾天兵林士杰
防護林科技 2015年2期
李巖,李娟,勾天兵,林士杰
(吉林省林業科學研究院,吉林 長春 130033)
紫椴(Tilia amurensis Rupr.),國家Ⅱ級保護植物[1]。近年來,其資源大量減少,主要是因為其種子具有休眠特性[2,3]。解除休眠的種子需要適宜的萌發環境條件,特別是溫度條件對種子順利萌發至關重要[4,5]。我們已經研究了解除休眠的紫椴種子萌發的適宜溫度及內含物的變化[6],本文旨在探索已經解除休眠的紫椴種子不同溫度下其萌發過程中內源激素的變化,揭示紫椴種子萌發的生理機制。
1 材料與方法
1.1 材料
試驗材料來源于吉林省臨江市。種子從植株上采下后,自然風干,置于4 ℃冰箱中備用。種子平均千粒質量(40.4 ± 2.88)g,平均含水量(10.82 ±1.66)%。
1.2 不同溫度下紫椴種子萌發過程中內源激素的變化
經層積處理解除休眠的種子用0.5%的KMnO4消毒30 min 并用自來水沖凈,然后置于鋪有雙層濾紙的發芽盒中,保持發芽床濕潤,發芽盒留有空隙,以保持通氣。將裝有種子的發芽盒置于不同溫度的培養箱中,在黑暗條件下進行萌發。萌發溫度設15℃/10 ℃(種子萌發最適溫度)和25 ℃(種子萌發最差溫度)2 種處理,分別于種子萌發的第0、4、8、12、16、20 天取樣,委托中國農業大學采用酶聯免疫法測定激素含量。
2 結果與分析
2.1 不同溫度下紫椴種子內源激素含量的變化
2.1.1 GA 含量的變化 從圖 1 可知,0 ~ 12 d 25℃和15 ℃/10 ℃下種子中GA 含量變化較為一致,是持續降低的,12 ~20 d 25 ℃下種子中GA 含量仍是持續降低的,而15 ℃/10 ℃下種子中GA 含量則出現了一個上升的過程。說明高溫下GA 含量降低,使種子萌發速度減慢,以緩解高溫造成的對種子完成正常生理活動的壓力。
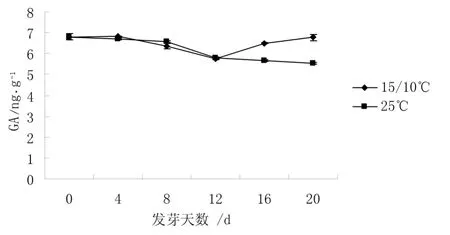
圖1 不同溫度下GA 含量的變化
2.1.2 ABA 含量的變化
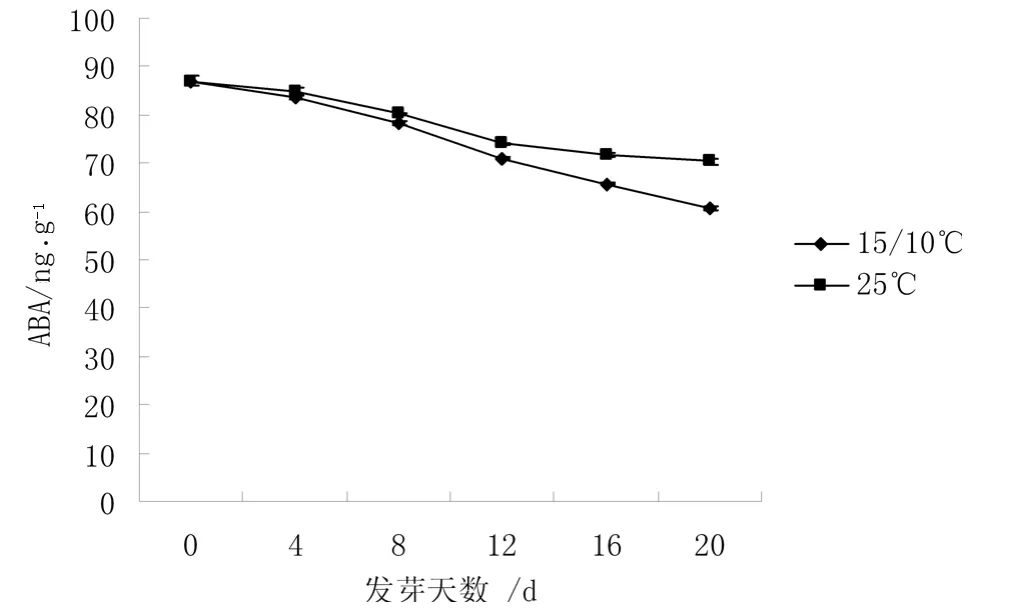
圖2 不同溫度下ABA 含量的變化
從圖2 可知,不同萌發溫度下,ABA 的變化趨勢基本一致,都是持續降低的。但是15 ℃/10 ℃下種子中ABA 含量始終低于25 ℃下種子中ABA 含量。萌發至16 d 時15 ℃/10 ℃和25 ℃下種子中ABA 含量分別比 0 d 降低了 24.52%和 17.44%。在整個種子萌發過程中,25 ℃下種子中ABA 含量始終高于15 ℃/10 ℃,表明種子發芽難易與內源ABA 含量有很大關系。
2.1.3 IAA 含量的變化
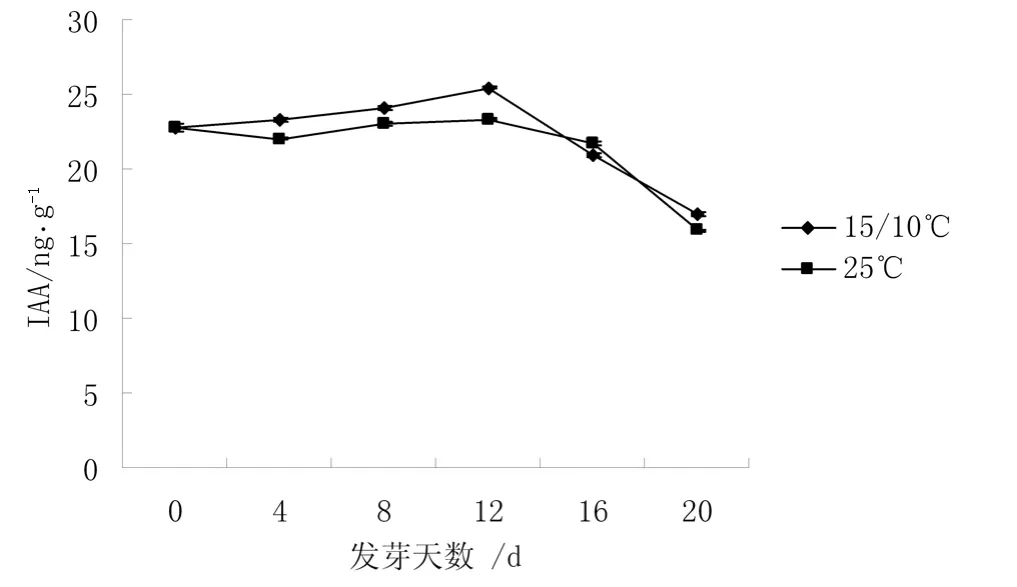
圖3 不同溫度下IAA 含量的變化
從圖3 可知,不同溫度下種子中IAA 含量變化趨勢基本一致,均呈先上升,再下降的趨勢。二者在12 d 時達到一個小高峰,且高峰值分別是0 d 時含量的 1.11 倍和 1.02 倍,而25 ℃下種子中 IAA 含量一直低于15 ℃/10 ℃,12 d 后二者都呈下降趨勢。高溫下IAA 含量降低,這可能是高溫下種子中ABA含量較高,ABA 與IAA 相拮抗有關。
2.2 不同溫度下激素比值的變化
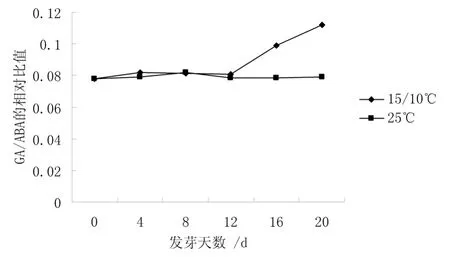
圖4 不同溫度下GA/ABA 比值的變化
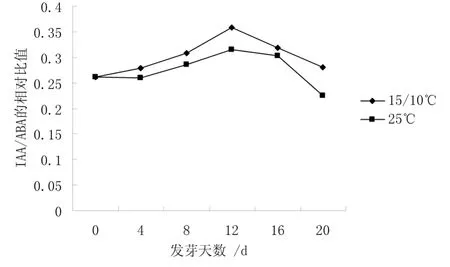
圖5 不同溫度下IAA/ABA 比值的變化
從圖4 可知,25 ℃下種子中GA/ABA 的相對比值基本不變,而15 ℃/10 ℃下種子中GA/ABA 的相對比值則以12 d 為轉折點,呈上升趨勢,20 d 時是0 d 的1.43 倍。說明 GA/ABA 比值的升高,能夠解除種子休眠,啟動種子萌發。從圖5 可知,25 ℃和15℃/10 ℃下種子中IAA/ABA 的比值變化趨勢非常一致,都是經歷了先上升后下降的過程。并以12 d為轉折點,15 ℃/10 ℃和25 ℃下種子中IAA/ABA的比值是 0 d 時的 1.36 倍和 1.20 倍。
3 結論與討論
許多植物種子中,休眠與萌發就是一個激素平衡的調節過程[7]。當種子在不適宜條件萌發時,內源ABA 含量保持較高水平,有ABA 的生物合成,萌發受到抑制[8,9]。本試驗結果表明,相對于適宜萌發溫度,25 ℃時ABA 含量增加,說明高溫脅迫對種子產生影響,會產生逆境脅迫信號物質ABA,其含量是萌發的關鍵因素。
IAA、GA 是種子體內重要的促進種子萌發的激素,因此高溫下,這類激素含量會發生變化,以協調種子的生理活動,適應環境的變化。高溫下,種子中GA、IAA 含量下降,造成種子萌發過程中,生長素類促進細胞建成和細胞的延伸,赤霉素類促進淀粉的分解及細胞的生長等一系列種子內部生理活動受到不同程度的抑制。植物內源激素在發揮其生理作用時并不是孤立的,而是相互聯系,相互制約的。雖然GA 含量比是解除種子休眠的啟動性因素,然而IAA與ABA 之間的關系對種子休眠解除和種子萌發也有重要的影響。本試驗表明,高溫下,使得種子中GA/ABA、IAA/ABA 的比值都顯著低于適溫(15 ℃/10 ℃),也說明當抑制型激素ABA 的含量占優勢時,種子的發芽便會受到抑制,處于休眠狀態。
[1]穆立薔,鄒琦,楊國亭,等.東北地區不同緯度紫椴枝葉解剖構造比較分析[J].東北林業大學學報,2007,35(4):25 -27
[2]王九齡,楊建平.紫鍛種子休眠原因的初步研究[J].林業科學,1981(3):317 -324
[3]劉艷秋.紫鍛種子休眠原因的初步研究[J].科技信息,2010(34):409
[4]Baskin Carol C,Baskin Jerry M.Seeds:ecology,biogeography,and evolution of dormancy and germination[J].Academic Press,2001:63 -64
[5]AI-Taisan Wafaa A.Comparative effects of drought and salt stress on germination and seedling growth of Pennisetum divisum (Gmel.)Henr.[J].American Journal of Applied Sciences,2010,7(5):640-646
[6]段加玉,勾天兵,謝朋,等.溫度對紫椴種子萌發及萌發過程中物質轉化的影響.中國農學通報,2013,29(19):31 -34
[7]Ramon G,Leon-Gonzalez.Genetic physiological characterization of seed dormancy regulation in common waterhemp〔Amaranthus tuberculatus (Moq.)sauer.〕[D].Iowa State University Graduation Thesis,2005
[8]Yoshioka T,Bndo T,Satoh S.Restoration of seed germination at superoptimal temperature by fluridone,an inhibitor of abscisic acid synthesis[J].Plant Cell Physiology,1998,39(7):307 -312
[9]Grappin P,Bouinot D,Sotta B,et al.Control of seed dormancy in Nicotiana plumbaginifolia:post-imbition abscisic acid synthesis imposes dormancy maintenance[J].Planta,2000,210(2):279 -285
