新疆阿勒泰山地蜱傳腦炎疫源地調(diào)查
2015-06-15 19:16:37張桂林劉曉明楊靚靚
中國(guó)人獸共患病學(xué)報(bào) 2015年12期
孫 響,張桂林,劉 然,劉曉明,楊靚靚,鄭 重,趙 焱
新疆阿勒泰山地蜱傳腦炎疫源地調(diào)查
孫 響1,張桂林1,劉 然1,劉曉明1,楊靚靚2,鄭 重1,趙 焱1
目的 調(diào)查新疆阿勒泰山地蜱傳腦炎疫源地特征,分離鑒定蜱傳腦炎病毒。方法 通過(guò)家畜體表?yè)旆ú杉纳纾焕瞄g接免疫熒光法檢測(cè)當(dāng)?shù)亟】等巳貉逯序鐐髂X炎病毒IgG抗體;通過(guò)將蜱研磨液接種實(shí)驗(yàn)小鼠進(jìn)行病原動(dòng)物分離;通過(guò)接種BHK-21細(xì)胞對(duì)蜱傳腦炎病毒進(jìn)行分離培養(yǎng);利用RT-PCR方法對(duì)病毒E蛋白基因片段進(jìn)行擴(kuò)增和測(cè)序,通過(guò)序列分析明確病毒系統(tǒng)進(jìn)化特征。結(jié)果 新疆阿勒泰山地白哈巴地區(qū)分布有2種蜱,森林革蜱為優(yōu)勢(shì)種(55.6%),其次為邊緣革蜱(44.4%);當(dāng)?shù)厝巳候鐐髂X炎IgG抗體陽(yáng)性率5.31%(6/113);通過(guò)動(dòng)物試驗(yàn)和細(xì)胞分離培養(yǎng),從森林革蜱中分離出一株森林腦炎病毒;對(duì)病毒E蛋白基因序列的系統(tǒng)進(jìn)化分析表明,蜱傳腦炎病毒阿勒泰分離株屬于遠(yuǎn)東亞型。結(jié)論 首次從病原學(xué)上證實(shí)新疆阿勒泰山地存在蜱傳腦炎疫源地,媒介為森林革蜱,病毒流行株為遠(yuǎn)東亞型。
蜱傳腦炎;蜱傳腦炎病毒;疫源地;阿勒泰
蜱傳腦炎(Tick-borne encephalitis,TBE)在我國(guó)又被稱(chēng)為森林腦炎,是由蜱傳腦炎病毒(TBEV)引起的以中樞神經(jīng)系統(tǒng)病變?yōu)橹鞯淖匀灰咴葱约膊 ER床上多表現(xiàn)為高熱、劇烈頭痛、惡心、嘔吐、嗜睡、意識(shí)障礙及腦膜刺激征、肌張力減弱或消失等神經(jīng)系統(tǒng)病征,重癥患者病死率較高。蜱傳腦炎經(jīng)蜱叮咬傳播,蜱既是蜱傳腦炎病毒的傳播媒介,也是其宿主。蜱傳腦炎分布橫貫亞-歐大陸,根據(jù)地理分離株的基因組序列變異性可分3個(gè)亞型(Sub-types),包括為歐洲型、西伯利亞型與遠(yuǎn)東型[1]。亞型間病毒毒力差異大,歐洲亞型和西伯利亞型的致死率分別為0.5%~2%與6%~8%,而遠(yuǎn)東型致死率最高,為5%~20%[2]。
蜱傳腦炎在我國(guó)主要分布在東北林區(qū)(內(nèi)蒙古、黑龍江和吉林)和新疆地區(qū)[3]。新疆蜱傳腦炎疫源地分布在北天山一帶以及阿拉套山南坡[4-5]。阿勒泰山通過(guò)血清學(xué)證實(shí)存在人群蜱傳腦炎感染,但未分離出病原體。為進(jìn)一步調(diào)查阿勒泰山地蜱傳腦炎疫源地的分布、傳播媒介以及流行的病毒株亞型,我們于2011-2013年進(jìn)行了蜱傳腦炎調(diào)查。
1 材料與方法
1.1 調(diào)查地點(diǎn) 白哈巴地區(qū)(東經(jīng)86°47′,北緯48°41′)地處新疆阿勒泰地區(qū)阿爾泰山南麓,平均海拔1 300 m,屬北溫帶大陸性氣候。該地區(qū)與哈薩克斯坦接壤。為山地森林草原生境,主要是新疆針葉松、西伯利亞落葉松、云杉、冷杉等針葉林,間或與白樺等闊葉林混雜。
1.2 標(biāo)本采集
1.2.1 蜱標(biāo)本采集 2011年和2012年4-5月,通過(guò)家畜體表?yè)旆ú杉倚篌w外寄生蜱,通過(guò)布旗法采集游離蜱。蜱分類(lèi)鑒定后,死蜱用70%酒精保存,活蜱液氮低溫保存。
1.2.2 人群血清標(biāo)本采集 隨機(jī)選擇當(dāng)?shù)亟】等巳海忪o脈采血3~5 mL,分離血清,-20 ℃保存。
1.3 血清TBE IgG抗體檢測(cè) 采用IFA方法。血清1∶10稀釋。TBEV IgG抗體IFA檢測(cè)試劑盒由歐蒙醫(yī)學(xué)實(shí)驗(yàn)診斷股份公司(EUROIMMUN Medizinische Labordiagnostika AG)生產(chǎn),產(chǎn)品編號(hào)No.FI2661-1005G,試劑盒內(nèi)包括抗原片、羊抗人二抗、陽(yáng)性對(duì)照、陰性對(duì)照等。按照使用說(shuō)明操作。抗原片平衡至室溫,加樣板每個(gè)反應(yīng)區(qū)加入30 μL稀釋血清,將抗原片覆有生物薄片的一面朝下,蓋在加樣板對(duì)應(yīng)的凹槽里,室溫(18~25 ℃)溫育30 min。PBS-Tween緩沖液沖洗抗原片1 s,浸洗5 min。在另一潔凈加樣板的每個(gè)反應(yīng)區(qū)加入25 μL FITC標(biāo)記羊抗人IgG抗體,將抗原片蓋在加樣板的凹槽里,室溫避光溫育30 min;用PBS-Tween緩沖液流水沖洗抗原片1 s,浸洗5 min。甘油/PBS封片。每次實(shí)驗(yàn)按說(shuō)明書(shū)加入陽(yáng)性和陰性對(duì)照。熒光信號(hào)通過(guò)熒光顯微鏡(Nikon ECLIPSE 80i)觀察。陽(yáng)性參考值血清稀釋度≥ 1∶10。
1.4 動(dòng)物接種分離病原 將液氮保存的100只森林革蜱置于75%酒精中浸泡30 min,滅菌生理鹽水漂洗3次,加入含青霉素的PBS研磨破碎,3 000 r/min離心,上清液經(jīng)0.22 μm 濾器(Pall)過(guò)濾除菌。按0.5 mL/只劑量腹腔接種5~8周齡的BALB/c小鼠(購(gòu)于新疆醫(yī)科大學(xué)實(shí)驗(yàn)動(dòng)物中心),共接種5只。接種后每日觀察小鼠發(fā)病情況,發(fā)病小鼠瀕死前無(wú)菌解剖,取肝脾和腦組織,液氮保存。取腦組織研磨上清液繼續(xù)接種小鼠傳代。連續(xù)觀察3周。
1.5 病毒細(xì)胞培養(yǎng) 取發(fā)病小鼠腦組織研磨,離心,過(guò)濾,取0.5 mL研磨上清液接種BHK-21細(xì)胞,37 ℃孵育1 h,培養(yǎng)7~10 d,反復(fù)凍融3次,按3 000 r/min離心收集細(xì)胞培養(yǎng)上清,再次接種細(xì)胞,盲傳3代后觀察細(xì)胞病變現(xiàn)象。BHK-21細(xì)胞(Baby hamster kiney-21)經(jīng)含10%(v/v)胎牛血清(杭州四青)的DMEM培養(yǎng)基(Life technology)傳代培養(yǎng)分種于48孔細(xì)胞培養(yǎng)板。
1.6 病毒分子生物學(xué)鑒定
1.6.1 RNA提取及RT-PCR檢測(cè) 利用Trizol法對(duì)400 μL病變細(xì)胞培養(yǎng)上清液進(jìn)行病毒RNA提取,提取物溶于40 μL DEPC處理的去離子水,置于-70 ℃度冷藏備用。按照一步法RT-PCR試劑盒(Takara)說(shuō)明書(shū)操作,擴(kuò)增體系為25 μL,其中包括5 μL上清RNA提取物,蜱傳腦炎病毒E蛋白特異引物5′-TGG AGC TYG ACA AGA CCT CA-3′與 5′-TCC CAC YAG GAT CTT GGG CAA-3′)各1 μL(10 μmol/L)。擴(kuò)增條件如下:42 ℃反轉(zhuǎn)錄30 min。95 ℃熱變性5 min。30輪熱循環(huán)中,每輪循環(huán)95 ℃熱變性30 s,57 ℃退火30 s,72 ℃擴(kuò)增90 s,最后72 ℃延伸10 min。取5 μL擴(kuò)增產(chǎn)物進(jìn)行1.7%(m/v)瓊脂糖凝膠電泳檢測(cè)[6]。
1.6.2 序列比對(duì)分析 陽(yáng)性擴(kuò)增產(chǎn)物交予上海生工公司進(jìn)行測(cè)序。利用DNAstar 7.0進(jìn)行測(cè)序比對(duì),陽(yáng)性測(cè)序結(jié)果提交GenBank。利用MEGA 6軟件按Clustal-W方法進(jìn)行同源分析。
2 結(jié) 果
2.1 蜱種類(lèi)組成 在阿勒泰山地白哈巴地區(qū)共采集成蜱2種2 160只,森林革蜱(Dermacentorsilvarum)為優(yōu)勢(shì)種,占蜱數(shù)量組成的55.6%(1 200/2 160),其次為邊緣革蜱(Dermacentormarginatus),占蜱數(shù)量組成的44.4%(960/2 160)。4月中下旬至5月中旬,馬體上寄生有大量的蜱。
2.2 人群血清學(xué)調(diào)查 共檢測(cè)血清113份,其中男性104份,女性9份,年齡18~69歲,平均年齡25歲。陽(yáng)性血清6份,陽(yáng)性率5.31%(6/113)。陽(yáng)性者均為男性,年齡分布在20至28歲,職業(yè)為林區(qū)工作人員,16名哈薩克族牧民血清檢測(cè)無(wú)陽(yáng)性。
2.3 動(dòng)物病原分離實(shí)驗(yàn)結(jié)果 一組森林革蜱研磨液接種5只BALB/c小鼠,其中4只發(fā)病,發(fā)病率80%。1只觀察21 d未發(fā)病。小鼠接種后7~8 d開(kāi)始發(fā)病,初始癥狀為拒食、不活潑、毛聳、蜷縮、消瘦,后發(fā)展為后肢麻痹、癱瘓、瀕死。發(fā)病小鼠解剖后發(fā)現(xiàn),肝脾腫大,顱內(nèi)出血。取發(fā)病小鼠腦組織研磨液接種2代BALB/c小鼠,共接種7只。其中發(fā)病4只,發(fā)病率57.1%;未發(fā)病3只。發(fā)病癥狀與一代發(fā)病小鼠一致。取二代發(fā)病小鼠腦組織提取RNA后進(jìn)行一步法RT-PCR,黃病毒通用引物及TBEV E基因特異引物均擴(kuò)增出陽(yáng)性條帶,分別為250 bp及750 bp左右(見(jiàn)圖2)。
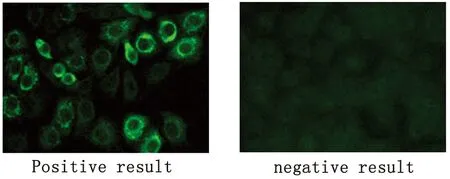
圖1 人血清TBE IgG抗體的間接免疫熒光檢測(cè)
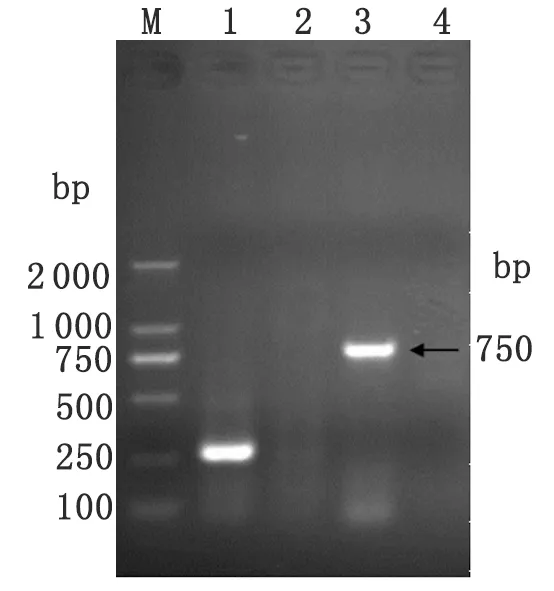
1: Positive result of flavivirus universal primers (250 bp); 2: Negative result; 3: Positive result of E protein gene primers (750 bp); 4: Negative control; M: DL2000 Marker.
圖2 森林腦炎病毒RT-PCR E蛋白基因片段擴(kuò)增結(jié)果
Fig.2 RT-PCR to E protein gene fragment of TBEV
陽(yáng)性結(jié)果測(cè)序后序列BLAST分析顯示,與我們從新疆博爾塔拉蒙古自治州夏爾希里自然保護(hù)區(qū)全溝硬蜱和森林革蜱中分離出的TBEV病毒株(JX534167)E蛋白基因核苷酸序列完全相同(100%,737/737)。
2.4 病毒細(xì)胞分離及分子生物學(xué)鑒定 將發(fā)病小鼠腦組織研磨、離心、過(guò)濾,上清接種BHK-21細(xì)胞,盲傳3代后,細(xì)胞出現(xiàn)圓縮、脫落等病變現(xiàn)象。病變細(xì)胞培養(yǎng)上清液進(jìn)行病毒RNA提取,利用RT-PCR對(duì)蜱傳腦炎病毒E蛋白基因片段進(jìn)行擴(kuò)增,擴(kuò)增出特異性條帶(約為750 bp)。測(cè)序后,與動(dòng)物實(shí)驗(yàn)發(fā)病小鼠腦組織RNA擴(kuò)增陽(yáng)性序列進(jìn)行比對(duì),兩者核苷酸序列完全相同,為同一條序列。由此,初步認(rèn)為經(jīng)過(guò)動(dòng)物接種純化及細(xì)胞分離培養(yǎng),已分離出阿勒泰白哈巴地區(qū)TBEV病毒株。
經(jīng)比對(duì)分析,擴(kuò)增序列與我們從新疆博爾塔拉蒙古自治州夏爾希里自然保護(hù)區(qū)全溝硬蜱和森林革蜱中分離出的TBEV病毒株(JX534167)E蛋白基因核苷酸序列相同。與我國(guó)東北分離毒株MDJ-01株(JQ650522)、武漢分離株WH2012(KJ755186)序列相似度為99%,與森張株(JQ650523)序列相似度為98%,表明阿勒泰分離毒株(Altai-01)為T(mén)BEV遠(yuǎn)東亞型。
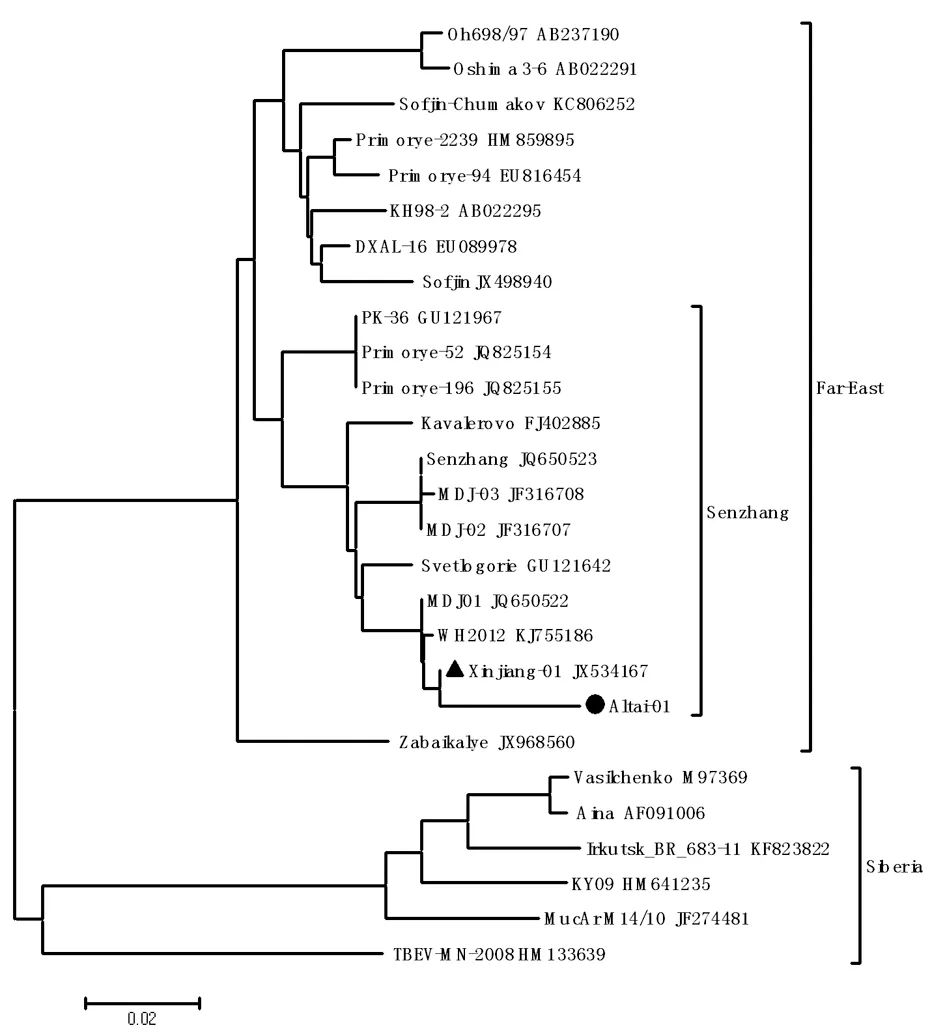
▲Xinjiang-01為分離自夏爾希里自然保護(hù)區(qū)全溝硬蜱的TBEV病毒株,●Altay-01為本研究分離自阿勒泰白哈巴地區(qū)森林革蜱的TBEV病毒株.
▲Xinjiang-01 was TBEV isolated fromIxodespersuleatusfrom Charles Hilary nature reserve;●Altay-01 was TBEV isolated fromDermacentorsilvarumfrom Baihaba area of Altai Mountains.
圖3 基于E蛋白基因的Neighbor-joining系統(tǒng)發(fā)生樹(shù)
Fig.3 Phylogenetic analysis of TBEV strain based on E protein gene with neighbor-joining method
3 討 論
既往對(duì)阿勒泰地區(qū)的研究在血清學(xué)上證實(shí)該地區(qū)人群中存在蜱傳腦炎感染,但未分離出病原體。本研究首次從阿勒泰山白哈巴地區(qū)的森林革蜱中分離到TBEV病毒株,從流行病學(xué)、血清學(xué)、病原學(xué)上證實(shí)了阿勒泰山地蜱傳腦炎疫源地的存在,并證實(shí)其病原體為T(mén)BEV遠(yuǎn)東亞型。
蜱傳腦炎的分布與媒介蜱的分布密切相關(guān)。我國(guó)北方地區(qū)蜱傳腦炎的主要媒介為全溝硬蜱(Ixodespersuleatus),南方地區(qū)主要媒介為卵形硬蜱(Ixodesovatus),在少數(shù)地區(qū)森林革蜱(Dermacentorsilvarum)也可作為媒介[7]。新疆北天山以及阿拉套山蜱傳腦炎疫源地雖然從全溝硬蜱和森林革蜱中都分離出了蜱傳腦炎病毒,但全溝硬蜱為疫源地優(yōu)勢(shì)蜱種,因此主要傳播媒介為全溝硬蜱。阿勒泰山地蜱傳腦炎疫源地與新疆其它疫源地不同,森林革蜱是優(yōu)勢(shì)蜱種,未發(fā)現(xiàn)全溝硬蜱,因此主要傳播媒介為森林革蜱。
血清學(xué)檢測(cè)陽(yáng)性者均為男性青壯年,在當(dāng)?shù)赜?~5年生活史,職業(yè)均為外來(lái)林區(qū)工作人員,表明該職業(yè)人群為T(mén)BE感染高危人群。而當(dāng)?shù)亻L(zhǎng)期生活的16名哈薩克族牧民血清檢測(cè)未發(fā)現(xiàn)陽(yáng)性。調(diào)查中發(fā)現(xiàn),16名牧民中僅1人有蜱叮咬史。不同人群血清陽(yáng)性率的差異可能與蜱的叮咬習(xí)性、病毒致病性、人群敏感性及人群防護(hù)措施的差異有關(guān),需要進(jìn)一步的流行病學(xué)調(diào)查分析加以證實(shí)。
在新疆地區(qū)境內(nèi),除了本研究從阿勒泰山地白哈巴地區(qū)分離到TBEV病毒株外,我們還曾從夏爾希里自然保護(hù)區(qū)分離到4株TBEV病毒株[4]。兩地區(qū)在地理位置、生境、媒介蜱種等方面均存在差異。白哈巴地區(qū)屬阿勒泰山脈,主要生境為山地森林草原,森林革蜱為優(yōu)勢(shì)蜱種,而夏爾希里自然保護(hù)區(qū)地處北天山西段阿拉套山南坡,屬天山山脈,主要生境為針闊葉混交林,全溝硬蜱為優(yōu)勢(shì)蜱種。但基于E蛋白基因的序列比對(duì)分析顯示,阿勒泰白哈巴分離株與夏爾希里分離株E蛋白基因片段無(wú)堿基差異。提示新疆境內(nèi)分布流行的TBEV遺傳基因可能比較穩(wěn)定,地區(qū)間未發(fā)生變異。
從全球來(lái)說(shuō),TBEV主要在一個(gè)帶狀區(qū)域內(nèi)分布流行,這個(gè)區(qū)域從西歐向東跨越俄羅斯、中國(guó)、蒙古直至日本。我國(guó)的新疆、內(nèi)蒙古和東北地區(qū)均位于這個(gè)流行帶內(nèi)[8-9]。通過(guò)對(duì)系統(tǒng)發(fā)育樹(shù)分析,新疆Altai-01 TBEV分離株與東北分離株(GenBank注冊(cè)號(hào)JQ650522)的相似度達(dá)99%,與蒙古TBEV-MN-2008分離株(HM133639)序列相似度僅為88%,但均為T(mén)BEV遠(yuǎn)東亞型[10];而新疆周邊地區(qū)如俄羅斯新西伯利亞(Novosibirsk)Vasilchenko分離株(M97369)、伊爾庫(kù)茨克市(Irkutsk) Aina分離株(AF091006)、蒙古國(guó)境內(nèi)MucAr M14/10 (JF274481)分離株、吉爾吉斯斯坦境內(nèi)KY09分離株(HM641235)均為T(mén)BEV西伯利亞亞型[1,10-12]。由此可見(jiàn)新疆TBEV病毒株與地理距離較遠(yuǎn)的東北TBEV病毒株有共同的祖先,而與地理距離相對(duì)較近的俄羅斯、吉爾吉斯斯坦、蒙古地區(qū)TBEV病毒株卻并不同源;提示新疆地區(qū)與東北地區(qū)之間存在著TBEV傳播感染的自然循環(huán),而鳥(niǎo)類(lèi)的遷徙在病毒的遠(yuǎn)距離傳播中可能起到了重要作用[13]。
[1]Ecker M, Allison SI, Meixner T, et al. Sequence analysis and genetic classification of tick-borne encephalitis viruses from Europe and Asia[J]. J Gen Virol, 1999, 80: 179-185. DOI: 10.1099/0022-1317-80-1-179
[2]Gristsun TS, Frolova TV, Zhankov AI, et al. Characterization of a Siberian virus isolated from a patient with progressive chronic tick-borne encephalitis[J]. J Virol, 2003, 77: 25-36.DOI: 10.1128/JVI.77.1.25 36.2003
[3]Bi WM, Deng HP, Bu XY. Study on the partition of natural foci of tick-borne encephalitis[J]. J Capital Normal Univ (Nat Sci Ed), 1997, 18(2): 100-107. (in Chinese) 畢維銘,鄧厚培,卜笑妍.蜱傳腦炎自然疫源地區(qū)劃研究[J].首都師范大學(xué)學(xué)報(bào):自然科學(xué)版,1997,18(2):100-107.
[4]Zhang GL, Liu R, Sun X, et al. Investigation on the endemic foci of new emerged tick-borne encephalitis in Charles Hilary, Xinjiang[J]. Chin J Epidemiol, 2013, 34(5): 438-442. DOI:10.3760/j.issn.0254-6450.2013.05.006.(in Chinese). 張桂林,劉然,孫響,等.新疆夏爾希里自然保護(hù)區(qū)蜱傳腦炎疫源地調(diào)查[J].中華流行病學(xué)雜志,2013,34(5):438-442.DOI:10.3760/j.issn.0254-6450.2013.05.006
[5]Xie XC, Yu X, Zhang TX, et al. A survey report on the natural foci of Russian Spring Summer Encephalitis in the Mountainous Areas of Tianshan and Altay Mountain in Xinjiang[J]. Endem Dis Bull, 1991, 6(3): 109-114. (in Chinese). 謝杏初,于心,張挺秀,等.新疆天山、阿爾泰山山地蜱傳腦炎自然疫源地考察報(bào)告[J].地方病通報(bào),1991,6(3):109-114.
[6]Ruzek D,Stastn H,Kopecky J, et al. Rapid subtyping of tick-borne encephalitis virus isolates using multiplex RT-PCR[J]. J Virolog Methods, 2007, 144: 133-137.DOI:10.1016/j.jviromet.2007.04.010
[7]Lu Z, Broker M, Liang G. Tick-borne encephalitis in mainland China[J]. Vector Borne Zoonotic Dis, 2008, 8(5): 713-720.DOI:10.1089/vbz.2008.0028
[8]Lars L, Olli V. Tick-borne encephalitis[J]. Lancet, 2008, 371(9627): 1861-1871.DOI:10.1016/S0140-6736(08)60800-4
[9]Eckhardt P, Dieter G, Olaf Z. Tick-borne encephalitis (TBE) trends in epidemiology and current and future management[J]. Travel Med Infect Dis,2010,8:233-245.DOI:10.1016/j.tmaid.2010.08.001
[10]Stefan F, Ilona M, Damdin A, et al. Isolation, preliminary characterization, and full-genome analyses of tick-borne encephalitis virus from Mongolia[J]. Virus Genes, 2012, 45: 413-425.DOI:10.1007/s11262-012-0795-9
[11]Kovalev SY, Kokorev VS, Belyaeva IV. Distribution of far-eastern tick-borne encephalitis virus subtype strains in the former soviet union[J]. J Gen Virol, 2010, 91: 2941-2946.DOI:10.1099/vir.0.023879-0
[12]Briggs BJ, Atkinson B, Czechowski DM,et al. Tick-borne encephalitis virus, Kyrgyzstan[J]. Emerg Infect Dis, 2011, 17(5): 876-879.DOI:10.3201/eid1703.101183
[13]Waldenstrom J, Lundkvist A, Falk KI, et al. Migrating birds and tickborne encephalitis virus[J]. Emerg Infect Dis, 2007, 13(8): 1215-1218.DOI:10.3201/eid1308.061416
Tick-borne encephalitis endemic foci in Altai Mountains, Xinjiang, China
SUN Xiang1,ZHANG Gui-lin1,LIU Ran1,LIU Xiao-ming1,YANG Liang-liang2,ZHENG Zhong1, ZHAO Yan1
(1.CenterforDiseasePreventionandControlofXinjiangMilitaryCommandRegion,Urumuqi830011,China; 2.PLA16thHospital,Altai836599,China)
In order to investigate endemic region of tick-borne encephalitis in Altai Mountains of Northern Xinjiang and to isolate and characterize viral geographic strains, we collected 2 160 ticks from Baihaba area located in Altai Mountains with searching for hosts and 113 serum of local healthy residents. The detection of TBEV specific IgG antibodies from serum were performed by indirect fluorescent assay (IFA), and viruses were isolated from ticks samples by inoculating BALB/c mice and BHK-21 cells. The virus E protein gene fragments were amplified and sequenced using the RT-PCR method. Results showed thatDermacentorsilvarum(55.6%) were dominant species followed byDermacentormarginatus(44.4%). TBEV specific-IgG antibodies in serum were positive with the rate of 5.31% (6/113). ATBEV strain was isolated fromDermacentorsilvarumafter animal inoculation and cells culture. E protein gene alignment sequence and phylogenetic analyses showed that the virus was clustered into TBEV far-eastern sub-type. Results demonstrated a novel endemic foci of tick-borne encephalitis was firstly described in Altai Mountains, northern Xinjiang. The TBEV geographic isolates were belonged to TBEV far-eastern subtype.Dermacentorsilvarumplayed a crucial role for transmitting the disease.
tick-borne encephalitis; tick-borne encephalitis virus; endemic foci; Altai Mountains
Zhang Gui-lin, Email:xjglzhang@126.com
10.3969/j.issn.1002-2694.2015.12.021
國(guó)家自然科學(xué)基金項(xiàng)目(No.U1303104)
張桂林,Email:xjglzhang@126.com
1.新疆軍區(qū)疾病預(yù)防控制中心,烏魯木齊 830011; 2.新疆軍區(qū)第十六醫(yī)院,阿勒泰 836599
R384.4
A
1002-2694(2015)12-1189-04
2015-03-18;
2015-09-23
Supported by grants from the National Natural Science Fundation (No. U1303104)
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級(jí))(2025年8期)2025-08-18 00:00:00
中老年保健(2021年3期)2021-08-22 06:50:04
四川文學(xué)(2021年4期)2021-07-22 07:11:54
天津醫(yī)科大學(xué)學(xué)報(bào)(2021年2期)2021-03-29 05:31:08
現(xiàn)代臨床醫(yī)學(xué)(2021年1期)2021-01-26 00:56:02
學(xué)苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
絲綢之路(2014年9期)2015-01-22 04:24:46
云南中醫(yī)學(xué)院學(xué)報(bào)(2014年3期)2014-07-31 18:57:34
現(xiàn)代檢驗(yàn)醫(yī)學(xué)雜志(2014年4期)2014-02-02 02:44:59
七彩語(yǔ)文·畫(huà)刊(2012年3期)2012-04-29 00:00:00
 中國(guó)人獸共患病學(xué)報(bào)2015年12期
中國(guó)人獸共患病學(xué)報(bào)2015年12期
- 中國(guó)人獸共患病學(xué)報(bào)的其它文章
- 四川省鉤端螺旋體病高發(fā)縣報(bào)告病例的準(zhǔn)確性調(diào)查研究
- 皮內(nèi)變態(tài)反應(yīng)試驗(yàn)和IFN-γ試驗(yàn)在奶牛結(jié)核病檢測(cè)中的比較研究
- 牛源大腸桿菌O157∶H7分離與鑒定及其毒力基因檢測(cè)
- 以減毒沙門(mén)氏菌為載體的抗賈第蟲(chóng)腸溶疫苗制備及免疫效果檢測(cè)
- 羊口瘡病毒F1L和B2L基因真核表達(dá)質(zhì)粒構(gòu)建及其在MDBK細(xì)胞中的表達(dá)
- 結(jié)核分枝桿菌T7噬菌體展示基因組DNA文庫(kù)的構(gòu)建與鑒定