鉬對(duì)蘋果砧木平邑甜茶幼苗硝態(tài)氮吸收、轉(zhuǎn)化及分配的影響
2015-06-15 19:18:51歐志鋒姜遠(yuǎn)茂魏紹沖
植物營(yíng)養(yǎng)與肥料學(xué)報(bào) 2015年3期
劉 利, 歐志鋒, 姜遠(yuǎn)茂, 魏紹沖
(作物生物學(xué)國家重點(diǎn)實(shí)驗(yàn)室,山東果蔬優(yōu)質(zhì)高效生產(chǎn)協(xié)同創(chuàng)新中心,山東農(nóng)業(yè)大學(xué)園藝科學(xué)與工程學(xué)院,山東泰安 271018)
鉬對(duì)蘋果砧木平邑甜茶幼苗硝態(tài)氮吸收、轉(zhuǎn)化及分配的影響
劉 利, 歐志鋒, 姜遠(yuǎn)茂, 魏紹沖*
(作物生物學(xué)國家重點(diǎn)實(shí)驗(yàn)室,山東果蔬優(yōu)質(zhì)高效生產(chǎn)協(xié)同創(chuàng)新中心,山東農(nóng)業(yè)大學(xué)園藝科學(xué)與工程學(xué)院,山東泰安 271018)
【目的】氮肥利用率低是制約果樹產(chǎn)業(yè)可持續(xù)發(fā)展的重要因素之一。過量施用氮肥以及不當(dāng)?shù)霓r(nóng)業(yè)措施造成當(dāng)前多數(shù)蘋果果園發(fā)生不同程度的土壤酸化,而鉬在酸性土壤中有效含量的降低會(huì)影響氮素吸收利用效率。本試驗(yàn)旨在探索能提高氮肥吸收利用效率的適宜鉬用量,了解鉬對(duì)蘋果砧木平邑甜茶(MalushupehensisRehd.) 幼苗硝態(tài)氮吸收、轉(zhuǎn)化和分配利用特性的影響,為蘋果生產(chǎn)中鉬肥與氮肥的合理施用提供科學(xué)數(shù)據(jù)。【方法】以平邑甜茶幼苗為試驗(yàn)材料,采用全硝態(tài)氮霍格蘭營(yíng)養(yǎng)液進(jìn)行培養(yǎng)。試驗(yàn)設(shè)置5個(gè)供鉬水平:營(yíng)養(yǎng)液不含鉬(CK);營(yíng)養(yǎng)液含0.25 μmol/L鉬酸(M1);營(yíng)養(yǎng)液含0.5 μmol/L鉬酸(M2);營(yíng)養(yǎng)液含1.5 μmol/L鉬酸(M3);葉片噴0.04%鉬酸,營(yíng)養(yǎng)液不含鉬(M4)。運(yùn)用15N同位素示蹤技術(shù),研究不同供鉬水平對(duì)幼苗15N吸收量、全氮量、分配率和利用率的影響,同時(shí)測(cè)定鉬對(duì)不同時(shí)期幼苗葉片和根系硝酸還原酶活性的影響。【結(jié)果】供鉬處理幼苗的根系活力不同程度地高于不施鉬處理(對(duì)照),其中營(yíng)養(yǎng)液含0.5 μmol/L鉬酸的處理(M2)效果最佳。培養(yǎng)8 d內(nèi)M2處理的平邑甜茶幼苗的根系硝酸還原酶活性一直高于其他處理,且與對(duì)照差異顯著;培養(yǎng)4 d時(shí)各種處理的葉片硝酸還原酶活性上升至最高值,隨后下降,8 d后又出現(xiàn)上升和下降的趨勢(shì),但上升幅度明顯小于培養(yǎng)4 d。 M1、M2、M3和M4處理的植株總干重分別比CK提高3.88%、21.12%、12.38% 和19.90%。與對(duì)照相比,0.5 μmol/L鉬酸處理幼苗的15N吸收量增加的比例最大,全氮量表現(xiàn)出相同的趨勢(shì)。供鉬處理的氮肥利用率均明顯高于對(duì)照,其中以0.5 μmol/L鉬酸處理的效果最顯著,利用率為13.97%,比對(duì)照高48.92%。施鉬處理對(duì)幼苗的15N分配率有一定的影響,0.5 μmol/L鉬酸處理(M2)和葉片噴施0.04%鉬酸處理(M4)的莖和葉片的15N分配率明顯高于對(duì)照,對(duì)照根系中15N分配率最高。0.5 μmol/L鉬酸處理葉片、莖和根的Ndff均達(dá)到最高,分別為對(duì)照同一部位的1.59倍、1.56倍和1.33倍。以上結(jié)果表明適量供鉬可促進(jìn)幼苗對(duì)肥料15N 的吸收征調(diào)利用。【結(jié)論】供鉬可以提高蘋果砧木平邑甜茶幼苗的生物量、根系活力、15N吸收量、全氮量和15N利用率,其中經(jīng)過0.5 μmol/L鉬酸處理(M2),其對(duì)平邑甜茶幼苗生長(zhǎng)及硝態(tài)氮吸收、轉(zhuǎn)化和分配利用的影響最顯著。
鉬; 平邑甜茶;15N; 吸收; 分配; 利用
鉬是高等植物正常生長(zhǎng)發(fā)育和生理代謝所必需的微量元素之一,其作用主要通過含鉬酶來實(shí)現(xiàn)[1-2]。目前能證明存在于高等植物體內(nèi)的有硝酸還原酶(Nitrate reductase,NR)、黃嘌呤脫氫酶(Xanthine dehydrogenase,XDH)、醛氧化酶(Aldehyde oxidase,AO)、亞硫酸鹽氧化酶(Sulfite oxidase,SO)4種[3-5]。其中硝酸還原酶是氮素同化過程中的關(guān)鍵限速酶,鉬是其活性成分,參與硝態(tài)氮還原為氨的過程,在植物的氮素代謝過程中發(fā)揮著重要作用[6-7]。研究還發(fā)現(xiàn),鉬可影響冬小麥葉綠素合成產(chǎn)物在葉綠體基質(zhì)內(nèi)的轉(zhuǎn)化,施鉬可促進(jìn)葉綠素的合成,進(jìn)而提高產(chǎn)量[8]。
植物氮素代謝中,銨態(tài)氮與硝態(tài)氮是兩個(gè)互相依賴的氮素形態(tài),它們之間保持著一定的平衡,平衡的協(xié)調(diào)程度與鉬供應(yīng)有關(guān)[9]。鉬可增強(qiáng)大豆和花生的根系活力和根瘤生物量,降低葉片硝態(tài)氮的含量,增強(qiáng)硝酸還原酶的活性,從而促進(jìn)植株氮素吸收利用以及氮素向籽粒的轉(zhuǎn)移[10-11]。在非豆科植物中,施鉬對(duì)小麥葉片硝酸還原酶有激活作用,能促進(jìn)小麥的氮代謝,降低體內(nèi)硝酸鹽的含量,增加銨態(tài)氮的含量,進(jìn)而提高氮肥利用效率[12-13]。鉬和鎳配合施用可以增加水稻葉片、莖和籽粒中的全氮含量,提高氨基酸和蛋白質(zhì)含量[14]。用鉬酸銨作基肥或噴施鉬酸銨增產(chǎn)顯著,亦可明顯降低花椰菜花球硝酸鹽含量,提高維生素C和可溶性糖含量[15]。對(duì)缺鉬植株葉面噴鉬可以明顯提高美樂葡萄坐果率和果穗重量,降低畸形果粒的比例[16]。可見,合理施用鉬肥可以提高硝態(tài)氮轉(zhuǎn)化利用效率以及有效地改善果實(shí)品質(zhì)。
當(dāng)前,不當(dāng)?shù)霓r(nóng)業(yè)措施及化肥過量施用等造成當(dāng)前山東省多數(shù)蘋果果園土壤發(fā)生不同程度的酸化,而鉬在酸性條件下易與土壤中的游離鐵、鋁生成沉淀[17],使得有效鉬含量降低,進(jìn)而影響氮素吸收利用效率,因此提高肥料利用率對(duì)保護(hù)生態(tài)環(huán)境和農(nóng)業(yè)可持續(xù)發(fā)展具有重要的意義。本試驗(yàn)以霍格蘭完全營(yíng)養(yǎng)液為培養(yǎng)基質(zhì),運(yùn)用同位素示蹤技術(shù),研究不同供鉬水平對(duì)平邑甜茶幼苗硝態(tài)15N吸收、轉(zhuǎn)化及分配特性的影響,為蘋果生產(chǎn)中鉬肥與氮肥的合理施用提供科學(xué)數(shù)據(jù)。
1 材料與方法
1.1 材料與試驗(yàn)設(shè)計(jì)

1.2 分析項(xiàng)目和方法
根系活力采用TTC法測(cè)定。硝酸還原酶(NR)活性參照李合生[18]的方法測(cè)定。植株解析參照文獻(xiàn)[19]進(jìn)行,分為根、莖和葉片3部分,放入烘箱中經(jīng)105℃殺青30 min,80℃烘干至恒重,稱重,研磨粉碎后過0.25 mm篩,裝袋備用。植株全氮用凱氏定氮法測(cè)定[20]。15N豐度在中國農(nóng)業(yè)科學(xué)院原子能利用研究所用MAT-251質(zhì)譜儀測(cè)定。
1.3 數(shù)據(jù)處理
Ndff (%) = (植物樣品中15N豐度%-15N自然豐度%)/(肥料中15N豐度%-15N自然豐度%)×100
器官15N 吸收量(g) = Ndff×器官全氮量(g)
器官全氮量(g)= 器官生物量(g)×氮含量(%)
氮肥利用率(%)= 器官15N 吸收量/施肥量(g)×100
氮肥分配率(%)= 各器官從氮肥中吸收的氮量(g)/總吸收氮量(g)×100
試驗(yàn)數(shù)據(jù)采用DPS7.05軟件進(jìn)行單因素方差分析,LSD 法進(jìn)行差異顯著性檢驗(yàn),應(yīng)用Microsoft Excel 2003繪制圖表。
2 結(jié)果與分析
2.1 不同供鉬水平對(duì)平邑甜茶幼苗根系活力的影響
根系活力水平的高低直接影響植物礦質(zhì)元素的吸收和地上部的生長(zhǎng)[21-22]。由圖1可以看出,各種供鉬幼苗的根系活力水平一直高于同期對(duì)照的水平。除對(duì)照在第4 d根系活力達(dá)到最大值之外,其余各處理均在第8 d達(dá)到最大值,M1、M2、M3和M4處理分別為CK的1.76倍、2.11倍、1.49倍和2.04倍,隨后各處理的根系活力開始下降。
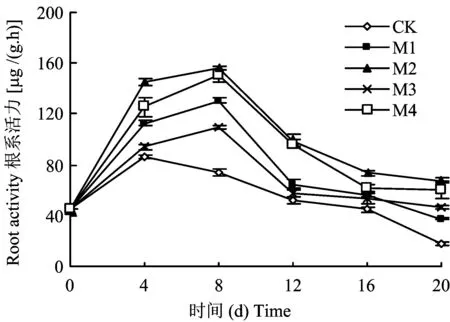
圖1 不同鉬水平對(duì)平邑甜茶幼苗根系活力的影響Fig.1 Effect of different Mo levels on root activity of Malus hupehensis Rehd. seedlings
2.2 不同供鉬水平對(duì)平邑甜茶幼苗根系硝酸還原酶(NR)活性的影響
硝酸鹽饑餓處理后的平邑甜茶幼苗體內(nèi)的硝態(tài)氮含量較低,當(dāng)轉(zhuǎn)移到全硝態(tài)氮營(yíng)養(yǎng)液后大量地吸收硝態(tài)氮,從而誘導(dǎo)根系硝酸還原酶活性的提高。因此硝酸還原酶活性4 d時(shí)快速上升,并于8 d時(shí)各處理均達(dá)到最大值(圖2)。培養(yǎng)8 d內(nèi),M2處理的幼苗根系硝酸還原酶活性一直高于其他處理,且與CK有顯著差異。培養(yǎng)至12 d時(shí),CK處理的幼苗根系硝酸還原酶活性高于其他處理,16 d后各種處理間差異不顯著。
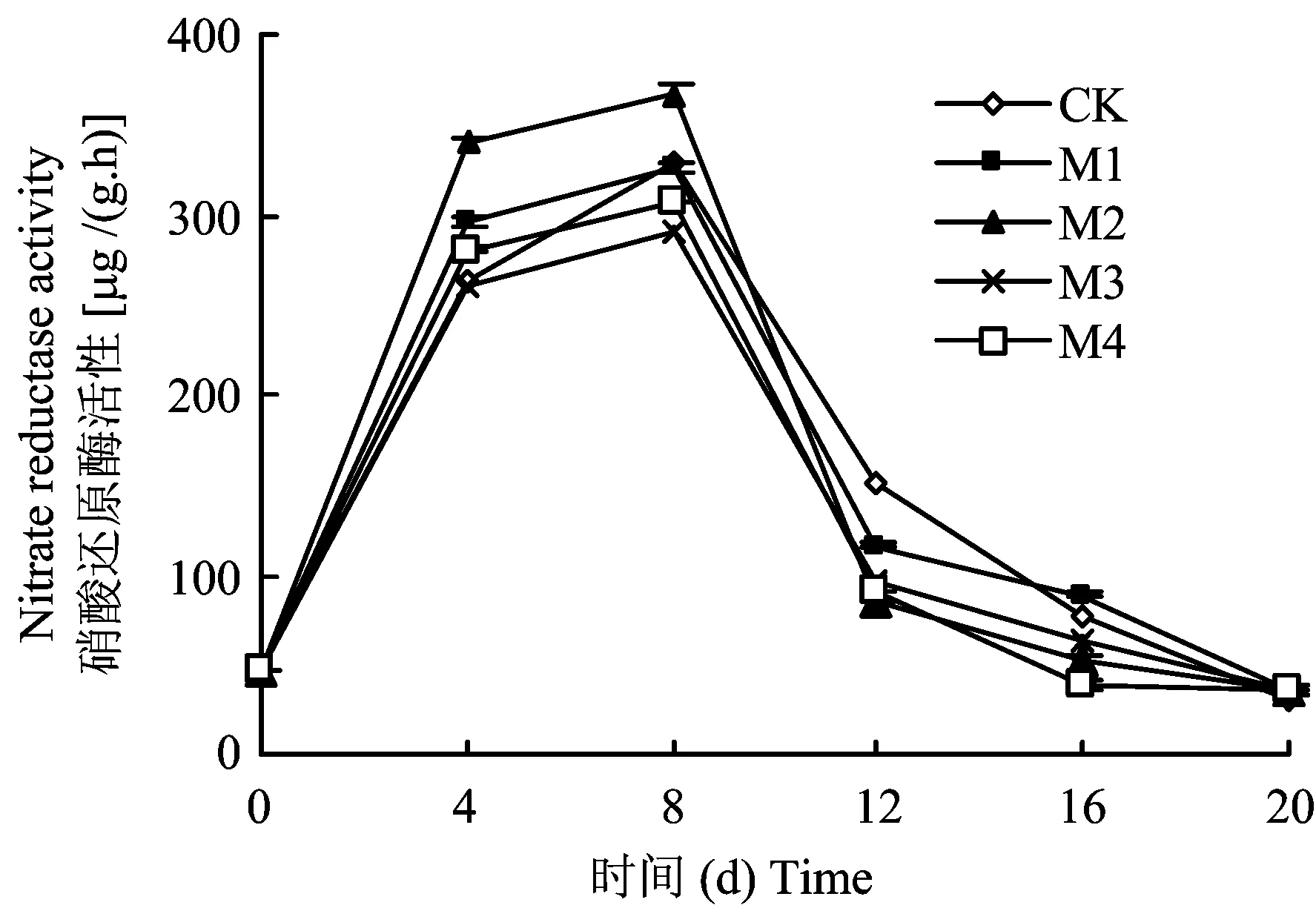
圖2 不同鉬水平對(duì)平邑甜茶幼苗根系硝酸還原酶活性的影響Fig.2 Effect of different Mo levels on nitrate reductase acitivity in roots of Malus hupehensis Rehd. seedlings
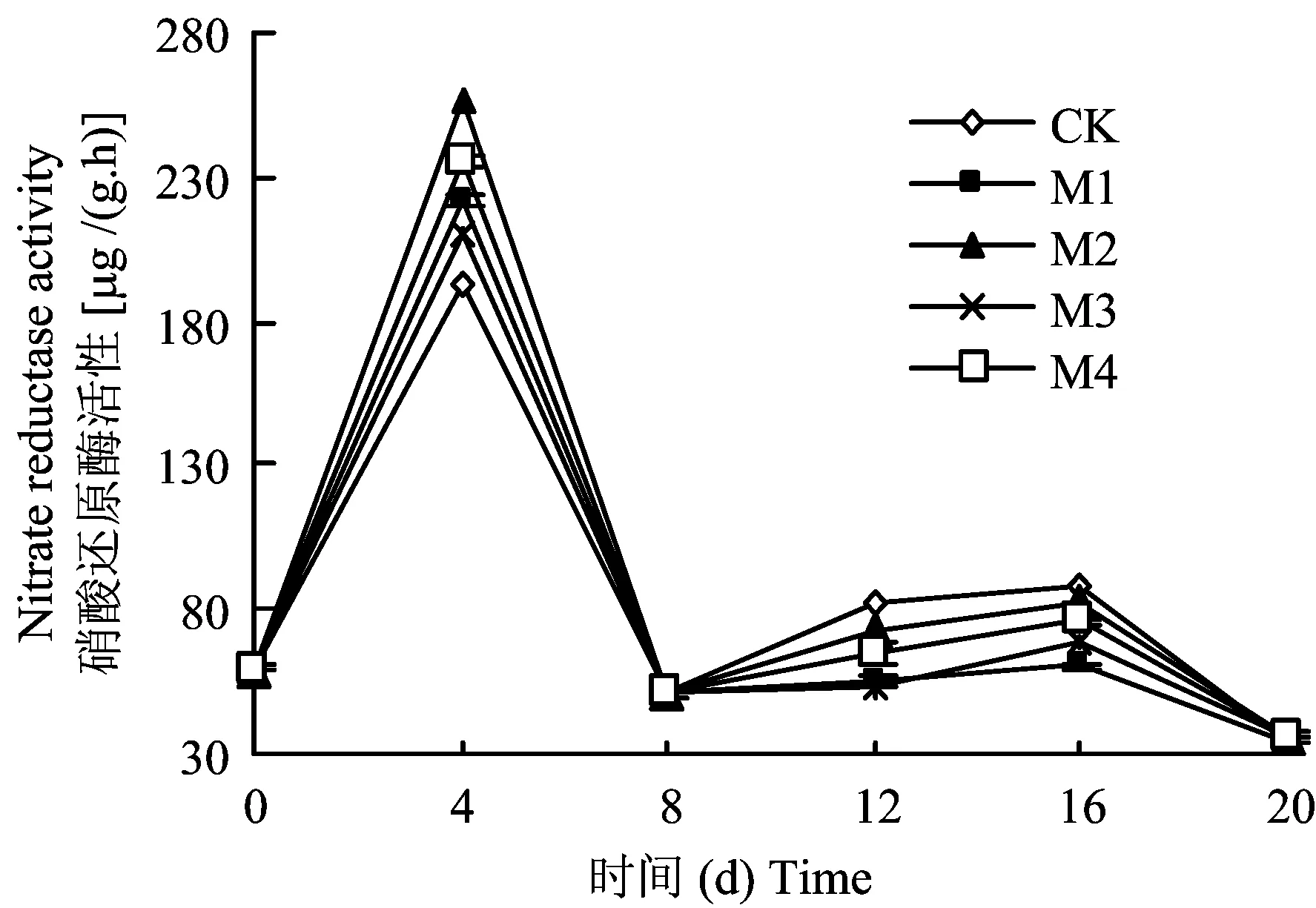
圖3 不同鉬水平對(duì)平邑甜茶幼苗葉片硝酸還原酶活性的影響Fig.3 Effect of different Mo levels on nitrate reductase acitivity in leaves of Malus hupehensis Rehd. seedlings
2.3 不同供鉬水平對(duì)平邑甜茶幼苗葉片硝酸還原酶(NR)活性的影響
平邑甜茶幼苗經(jīng)硝酸鹽饑餓處理后轉(zhuǎn)入到全硝態(tài)氮營(yíng)養(yǎng)液進(jìn)行培養(yǎng),4 d時(shí)各處理葉片的硝酸還原酶活性上升至最高值,然后下降,8 d后又出現(xiàn)上升和下降的趨勢(shì),但上升幅度明顯小于第4 d(圖3)。第4 d時(shí),M2處理的葉片硝酸還原酶活性比CK提高33.79%,而M3處理僅提高9.94%。第8 d時(shí),各處理葉片硝酸還原酶活性均降到50 μg/(g·h)左右。培養(yǎng)至12 d時(shí),對(duì)照處理的硝酸還原酶活性上升幅度最大。方差分析顯示,到20 d時(shí)各處理間幼苗葉片硝酸還原酶活性無顯著差異(P>0.05)。
2.4 不同供鉬水平對(duì)植株生物量的影響
由表1可以看出,M1、M2、M3和M4處理的總干重分別比對(duì)照提高3.88%、21.12%、12.38%和19.90%。葉片干重M4和M2處理提高的幅度最大。M2、M3和M4處理的根部干重也顯著高于對(duì)照,但這3個(gè)處理間差異不顯著。

表1 不同鉬水平下的植株生物量(g/plant)
注(Note): 同列數(shù)據(jù)后不同字母表示處理間差異達(dá)5%顯著水平 Values followed by different letters in a column are significant among treatments at the 5% level.
2.5 不同供鉬水平下植株各器官的Ndff
Ndff是指植株器官從肥料15N中吸收分配到的15N量對(duì)該器官全氮量的貢獻(xiàn)率,它反映了植株器官對(duì)肥料15N的吸收征調(diào)能力[23]。不同處理平邑甜茶幼苗各種器官的Ndff具有較大差異(表2)。M2處理葉片、莖和根的Ndff均達(dá)到最高,分別為對(duì)照的1.59倍、1.56倍和1.33倍。方差分析表明,M2和M4處理各種器官的Ndff均顯著高于對(duì)照(P<0.05)。
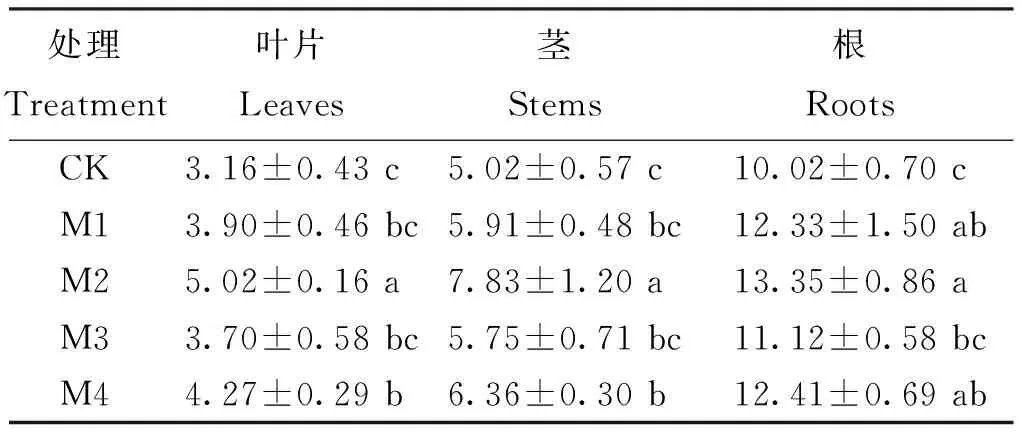
表2 不同鉬水平下植株各器官的Ndff(%)
注(Note): 同列數(shù)據(jù)后不同字母表示處理間差異達(dá)5%顯著水平 Values followed by different letters in a column are significant among treatments at the 5% level.
2.6 不同供鉬水平的植株各器官的全氮量和15N吸收總量
表3顯示,隨著營(yíng)養(yǎng)液供鉬水平的提高,平邑甜茶幼苗的全氮量和15N吸收總量呈先上升后下降的趨勢(shì)。與對(duì)照相比,M2處理的全氮量增加的比例最大,其次是M4和M3處理,M1處理最小。植株15N吸收總量也表現(xiàn)出同樣的趨勢(shì)。
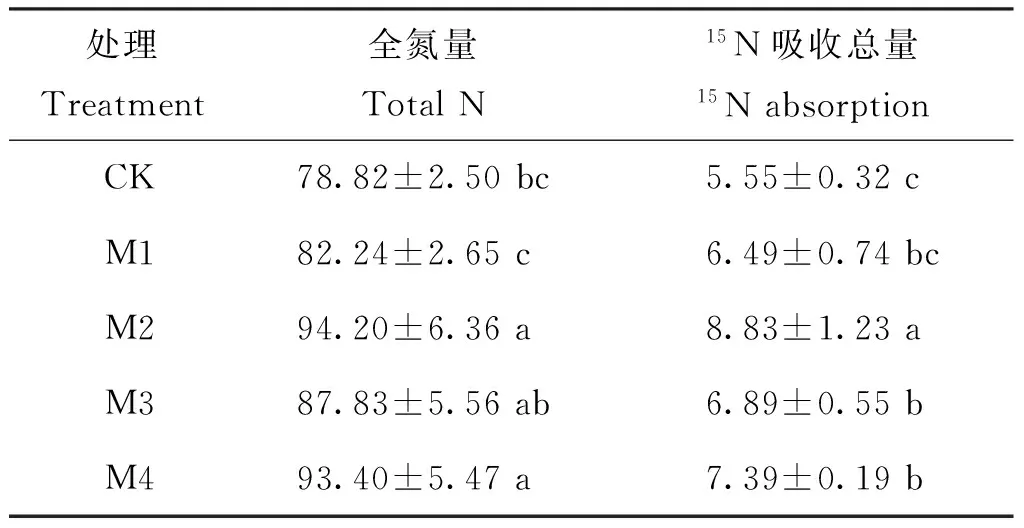
表3 不同鉬水平下植株的全氮量和15N吸收總量(mg)
注(Note): 同列數(shù)據(jù)后不同字母表示處理間差異達(dá)5%顯著水平 Values followed by different letters in a column are significant among treatments at the 5% level.
2.7 不同供鉬水平的植株15N利用率
不同供鉬水平之間氮肥利用率差異顯著,其中M2處理高達(dá)13.97%,而對(duì)照僅為9.38%(圖4)。 M1、M2、M3和M4處理分別比對(duì)照提高21.26%、48.92%、11.96%和38.44%。

圖4 不同鉬水平下植株的15N利用率(%)Fig.4 15N Use efficiency of seedlings under different Mo levels[注(Note): 柱上不同字母表示處理間差異達(dá)5% Different letters above the bars mean significant among treatments at the 5% level.]
2.8 不同供鉬水平的植株各器官15N分配率
器官中15N占全株15N總量的百分率反映了肥料氮在樹體內(nèi)的分布及在各器官遷移的規(guī)律[24]。15N主要分配在根系中,其次是莖,葉片最少(表4)。
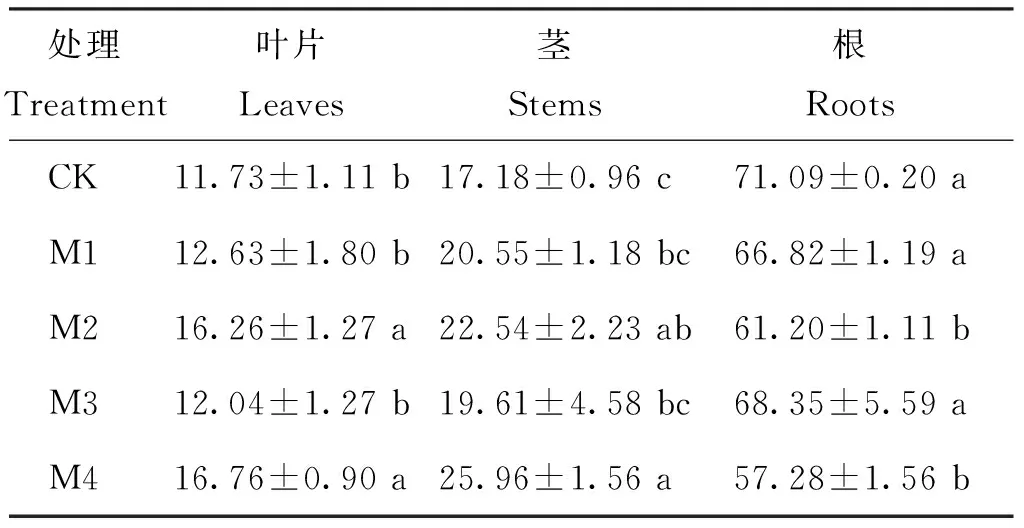
表4 不同鉬水平下植株各器官的15N 分配率(%)
注(Note): 同列數(shù)據(jù)后不同字母表示處理間差異達(dá)5%顯著水平 Values followed by different letters in a column are significant among treatments at the 5% level.
M2和M4處理地上部15N分配率明顯高于對(duì)照,葉片分別提高38.80%和42.72%。方差分析表明,M1和M3處理15N分配率和對(duì)照無顯著差異。
3 討論
適宜濃度鉬供應(yīng)不但可以有效地降低小麥硝態(tài)氮的累積,促進(jìn)銨態(tài)氮形成,而且能促進(jìn)小麥吸收更多的氮素,增加小麥有機(jī)氮含量[9]。單施鉬肥明顯降低大白菜硝酸鹽的含量,促進(jìn)氮素的吸收利用,鉬鋅與復(fù)合生物肥配施可提高大白菜品質(zhì),增加產(chǎn)量[30]。本試驗(yàn)研究表明,當(dāng)鉬濃度為0.5 μmol/L鉬酸時(shí)平邑甜茶幼苗的氮肥利用率為13.97%,而對(duì)照僅為9.38%,其比對(duì)照提升達(dá)48.92%,鉬濃度為0.25 μmol/L和1.5 μmol/L鉬酸處理的氮肥利用率均明顯低于0.5 μmol/L鉬酸處理,葉片噴鉬也可以起到提高氮肥利用率的效果。0.5 μmol/L鉬酸處理各種器官Ndff值均顯著高于對(duì)照,1.5 μmol/L鉬酸處理各種器官中的Ndff出現(xiàn)下降,與對(duì)照無顯著差異。0.5 μmol/L鉬酸處理根系活力較高,莖和葉片的15N分配率高于對(duì)照,但0.25 μmol/L和1.5 μmol/L鉬酸處理葉片和根部15N分配率和對(duì)照差異并不顯著,表明合適濃度的鉬供應(yīng)可能更有利于地上部的生長(zhǎng)發(fā)育,提高植株氮素吸收利用效率。由于葉片中的鉬可能難于向根系運(yùn)輸,因此噴鉬主要提高葉片硝酸還原酶活性,對(duì)根系硝酸還原酶活性影響不大。噴鉬處理的葉片15N分配率最高,而0.5 μmol/L鉬酸處理根系吸收的鉬則可以通過蒸騰作用運(yùn)輸?shù)饺~片,因此葉片和根系的硝酸還原酶活性都有較大程度提高(圖2,圖3)。
喻敏等[31]研究表明,鉬高效冬小麥品種97003和鉬低效冬小麥品種97014在相同缺鉬逆境下,97003鉬利用率較高,可獲得較高的產(chǎn)量,而97014則表現(xiàn)出嚴(yán)重的缺鉬癥狀,產(chǎn)量不到施鉬處理的一半。因此,鉬高效蘋果砧木品種篩選將對(duì)蘋果產(chǎn)業(yè)的良性發(fā)展具有重要意義。
4 結(jié)論
施鉬可不同程度地提高蘋果砧木平邑甜茶幼苗的根系活力水平及根系和葉片的硝酸還原酶活性,其中以營(yíng)養(yǎng)液濃度為0.5 μmol/L鉬酸處理的效果最好。0.5 μmol/L鉬酸處理的植株15N利用率最高,各種器官的Ndff值也均最大。供鉬可以提高植株莖和葉片的15N分配率,葉面噴施0.04%鉬酸提高的幅度最大,其次為0.5 μmol/L鉬酸處理。
[1] Mendel R R, Hansch R. Molybdoenzymes and molybdenum cofactor in plants[J]. Journal of Experimental Botany, 2002, 53(375): 1689-1698.
[2] Nivol C L, Leimkühler S. Molybdenum enzymes, their maturation and molybdenum cofactor biosynthesis inEscherichiacoli.[J]. Biochimica et Biophysica Acta, 2013, 1827(8-9): 1086-1101.
[3] Mcgratha S P, Micóa(chǎn) C, Curdy R, Zhao F J. Predicting molybdenum toxicity to higher plants: Influence of soil properties[J]. Environmental Pollution, 2010, 158: 3095-3102.
[4] Hille R, Nishino T, Bittner F. Molybdenum enzymes in higher organisms[J]. Coordination Chemistry Reviews, 2011, 255(9): 1179-1205.
[5] Zdunek Z E, Lips H S. Plant molybdoenzymes and their response to stress[J]. Acta Physiologiae Plantarum, 2003, 25(4): 437-452.
[6] Kaiser B N, Gridley K L, Brady J Netal. The role of molybdenum in agricultural plant production[J]. Annals of Botany, 2005, 96(5): 745-754.
[7] Mendel R R, Kruse T. Cell biology of molybdenum in plants and humans[J]. Biochimica et Biophysica Acta, 2012, 1823(9): 1568-1579.
[8] 喻敏, 胡承孝, 王運(yùn)華. 低溫條件下鉬對(duì)冬小麥葉綠素合成前體的影響[J]. 中國農(nóng)業(yè)科學(xué), 2006, 39(4): 702-708. Yu M, Hu C X, Wang Y H. Effect of molybdenum on the precursors of chlorophyll biosynthesis in winter wheat cultivars under low temperature[J]. Scientia Agricultura Sinica, 2006, 39(4): 702-708.
[9] 門中華, 李生秀. 鉬對(duì)冬小麥硝態(tài)氮代謝的影響[J]. 植物營(yíng)養(yǎng)與肥料學(xué)報(bào), 2005, 11(2): 205-210. Men Z H, Li S H. Effects of molybdenum on nitrate metabolism of winter wheat[J]. Plant Nutrition and Fertilizer Science, 2005, 11(2): 205-210.
[10] 劉鵬, 楊玉愛. 鉬、硼對(duì)大豆氮代謝的影響[J]. 植物營(yíng)養(yǎng)與肥料學(xué)報(bào), 1999, 5(4): 347-351. Liu P, Yang Y I. Effect of molybdenum and boron on nitrogen metabolism of soybean[J]. Plant Nutrition and Fertilizer Science, 1999, 5(4): 347-351.
[11] 張智猛, 萬書波, 戴良香, 等. 施氮水平對(duì)不同花生品種氮代謝及相關(guān)酶活性的影響[J]. 中國農(nóng)業(yè)科學(xué), 2011, 44(2): 280-290. Zhang Z M, Wan S B, Dai L Xetal. Effects of nitrogen application rates on nitrogen metabolism and related enzyme activities of two different peanut cultivars[J]. Scientia Agricultura Sinica, 2011, 44(2): 280-290.
[12] 孫學(xué)成, 胡承孝, 譚啟玲, 等. 施用鉬肥對(duì)冬小麥游離氨基酸,可溶性蛋白質(zhì)和糖含量的影響[J]. 華中農(nóng)業(yè)大學(xué)學(xué)報(bào), 2002, 21(1): 40-43. Sun X C, Hu C X, Tan Q Letal. Effects of molybdenum application on contents of free amino acid, soluble sugar and protein of winter wheat at different growth stages[J]. Journal of Huazhong Agricutural University, 2002, 21(1): 40-43.
[13] Yu M, Hu C X, Sun X C, Wang Y H. Influences of Mo on nitrate reductase, glutamine synthetase and nitrogen accumulation and utilization in Mo-efficient and Mo-inefficient winter wheat cultivars[J]. Scientia Agricultura Sinica, 2010, 9(3): 355-361.
[14] Moraes M F, Reis A R, Moraes L A Cetal. Effects of molybdenum, nickel and nitrogen sources on the mineral nutrition and growth of rice plants[J]. Communications in Soil Science and Plant Analysis, 2009, 40(21-22): 3238-3251.
[15] 朱鳳林. 鉬、 硼對(duì)花椰菜產(chǎn)量及品質(zhì)的影響[J]. 園藝學(xué)報(bào), 2005, 32(2): 310-313. Zhu F L. Effects of Mo and B on the yield and quality of cauliflower[J]. Acta Horticiturae Sinca, 2005, 32(2): 310-313.
[16] Longbottom M L, Dry P R, Sedgley M. Effects of sodium molybdate foliar sprays on molybdenum concentration in the vegetative and reproductive structures and on yield components ofVitisviniferacv. Merlot[J]. Austalian Journal of Grape Wine Research, 2010, 16(3): 477-490.
[17] 龔子同, 黃標(biāo), 歐陽洮. 我國土壤地球化學(xué)及其在農(nóng)業(yè)生產(chǎn)中的意義[J]. 地理科學(xué), 1998, 18(1): 1-9. Gong Z T, Huang B, Ouyang T. Pedogeochemistry of China and its significance in agriculture[J]. Scientia Geographica Sinica, 1998, 18(1): 1-9.
[18] 李合生. 植物生理生化實(shí)驗(yàn)原理和技術(shù)[M]. 北京: 高等教育出版社, 2000. 123-128. Li H S. Experimental principle and technique for plant physiology and biochemistry[M]. Beijing: Higher Education Press, 2000. 123-128.
[19] 王海寧, 葛順峰, 姜遠(yuǎn)茂, 等. 施氮水平對(duì)五種蘋果砧木生長(zhǎng)、以及氮素吸收、分配和利用特性的影響[J]. 植物營(yíng)養(yǎng)與肥料學(xué)報(bào), 2012,18(5): 1262-1268. Wang H N, Ge S F, Jiang Y Metal. Effects of nitrogen fertilization on growth characteristics and absorption, distribution and utilization of NH415NO3of five apple rootstocks[J]. Plant Nutrition and Fertilizer Science, 2012, 18(5): 1262-1268.
[20] 鮑士旦. 土壤農(nóng)化分析[M]. 北京: 中國農(nóng)業(yè)出版社, 2000. Bao S D. Analytical methods for soil agro-chemistry [M]. Beijing: China Agricultural Press, 2000.
[21] 生利霞, 束懷瑞. 低氧脅迫對(duì)平邑甜茶根系活力及氮代謝相關(guān)酶活性的影響[J]. 園藝學(xué)報(bào), 2008, 35(1): 7-12. Sheng L X, Shu H R. Effects of hypoxia on the root activity, respiratory rate and the activities of enzymes involved in nitrogen metabolism in roots ofMalushupehensisRehd.[J]. Acta Horticiturae Sinca, 2008, 35(1): 7-12.
[22] Li Z J, Xie X Y, Zhang S Q, Liang Y C. Negative effects of oxytetracycline on wheat(TriticumaestivumL.) growth, root activity, photosynthesis, and chlorophyll contents[J]. Scientia Agricultura Sinica, 2011, 10(10): 1545-1553.
[23] 顧曼如, 束懷瑞, 周宏偉. 蘋果氮素營(yíng)養(yǎng)研究Ⅴ. 貯藏N的運(yùn)轉(zhuǎn)、分配特性[J]. 園藝學(xué)報(bào), 1986, 13: 25-30. Gu M R, Shu H R, Zhou H W. A study on the nitrogen nutrition of apple trees Ⅴ. The characters of translocation and distribution of the reserved15N[J]. Acta Horticiturae Sinca, 1986, 13: 25-30.
[24] 徐季娥, 林裕益, 呂瑞江, 等. 鴨梨秋施15N-尿素的吸收與分配[J]. 園藝學(xué)報(bào), 1993, 20(2): 145-149. Xu J E, Lin Y Y, Lu R Jetal. Studies on the absorption and the distribution of15N-labelled urea to‘Yali’pear trees following autumn application[J]. Acta Horticiturae Sinca, 1993, 20(2): 145-149.
[25] 彭福田, 姜遠(yuǎn)茂, 顧曼如, 束懷瑞. 氮素對(duì)蘋果果實(shí)內(nèi)源激素變化動(dòng)態(tài)與發(fā)育進(jìn)程的影響[J]. 植物營(yíng)養(yǎng)與肥料學(xué)報(bào), 2003, 9(2): 208-213. Peng F T, Jiang Y M, Gu M R, Shu H R. Effect of nitrogen on apple fruit hormone changing trends and development[J]. Plant Nutrition and Fertilizer Science, 2003, 9(2): 208-213.
[26] Warner J, Zhang T Q, Hao X. Effects of nitrogen fertilization on fruit yield and quality of processing tomatoes[J]. Canadian Journal of Plant Science, 2004, 84(3): 865-871.
[27] Raese J T, Drake S R, Curry E A. Nitrogen fertilizer influences fruit quality, soil nutrients and cover crops, leaf color and nitrogen content, biennial bearing and cold hardiness of ‘Golden Delicious’[J]. Journal of Plant Nutrition, 2007, 30(10): 1585-1604.
[28] Mendel R R, Bittner F. Cell biology of molybdenum[J]. Biochimica et Biophysica Acta, 2006, 1763(7): 621-635.
[29] 孫學(xué)成, 胡承孝. 高等植物含鉬酶與鉬營(yíng)養(yǎng)[J]. 植物生理學(xué)通訊, 2005, 41(3): 395-399. Sun X C, Hu C X. Molybdoenzymes and molybdenum nutrition in higher plants[J]. Plant Physiology Communications, 2005, 41(3): 395-399.
[30] 段曉琴. 鉬鋅與不同肥料配施對(duì)大白菜硝酸鹽積累及品質(zhì)的影響[J]. 北方園藝, 2010,(22): 53-55. Duan X Q. Effects of Mo and Zn microelement fertilizer prescribe fertilizing on nitrate accumulation and qualities of Chinese cabbage[J]. Northern Horticulturae, 2010,(22): 53-55.
[31] 喻敏, 胡承孝, 王運(yùn)華. 不同鉬效率冬小麥品種鉬的吸收和分配[J]. 中國農(nóng)業(yè)科學(xué), 2004, 37(11): 1749-1753. Yu M, Hu C X, Wang Y H. Molybdenum efficiency in winter wheat cultivars as related to molybdenum uptake and distribution[J]. Scientia Agricultura Sinica, 2004, 37(11): 1749-1753.
Effect of molybdenum on absorption, transformation and distribution of nitrate-nitrogen of apple rootstockMalushupehensisRehd. seedlings
LIU Li, OU Zhi-feng, JIANG Yuan-mao, WEI Shao-chong*
(StateKeyLaboratoryofCropBiology/ShandongCollaborativeInnovationCenterforFruitandVegetableProductionwithHighQualityandEfficiency/CollegeofHorticultureScienceandEngineering,ShandongAgriculturalUniversity,Tai’an,Shandong271018,China)
【Objectives】 The low nitrogen fertilizer use efficiency restricts sustainable development of fruit industry. Excessive application of nitrogen fertilizer and improper agricultural measures lead to soil acidification with different degrees in current apple orchards in China, and soil acidification can cause low content of soil available molybdenum, which decreases nitrogen fertilizer use efficiency. The aim of the study was to explore appropriate molybdenum(Mo)level to enhance nitrogen fertilizer use efficiency ofMalushupehensisRehd. seedlings, and provide scientific data for utilization of nitrogen and molybdenum fertilizer in apple industry. 【Methods】MalushupehensisRehd. seedlings were used as experiment materials and the Hoagland solution with nitrate N was used as only N source in the study. The experiment included five different Mo levels: control(CK, no Mo in solution), M1(0.25 μmol/L molybdic acid in solution), M2(0.5 μmol/L molybdic acid in solution), M3(1.5 μmol/L molybdic acid in solution) and M4(spraying 0.04% molybdic acid,no Mo in solution). Effects of Mo on absorption, transformation, and distribution of nitrate-nitrogen in the seedlings were detected using the15N-labeled tracer method, meanwhile effects of Mo on nitrate reductase(NR) activity of leaves and roots inMalushupehensisRehd. seedlings were analysed in the study. 【Results】 Mo can enhance root activity of the seedlings with different degrees and the treatment with 0.5 μmol/L Mo in solution has the highest effect in the study. The NR activity in roots of the treatment with 0.5 μmol/L Mo is higher than that of the other treatments on the 8 d, and there is significant difference between the treatment with 0.5 μmol/L Mo and the control. The leaf NR activities in all the treatments reach the maxima on the 4 d and then decrease. The dry weight amounts of M1, M2, M3 and M4 treatments are higher than that of the control and are increased by 3.88%, 21.12%, 12.38% and 19.90%, respectively. The15N absorption of M2 treatment is the largest and as well as the total N. The15N utilization ratios of all Mo treatments are higher than that of the control, and the treatment with 0.5 μmol/L Mo has the highest ratio of 13.97%, which is 48.92% higher than that of the control. Mo also has obvious effect on the15N distribution ratios in the seedlings. The15N ratios in stems and leaves of M2 treatment and M4 treatment are significantly higher than those of the control, while the highest15N distribution ratio in roots is found in the control. The Ndff values of leaves, stems and roots of M2 treatment are the highest among all the treatments, which are 1.59-, 1.56-and 1.33-fold compared with the same part in the control respectively. These results show that appropriate Mo supply can enhance the15N absorption,15N distribution and15N utilization of the seedlings. 【Conclusions】 Mo can enhance biomass, root activity,15N absorption, the total N and the15N utilization ratio ofMalushupehensisRehd.seedlings. In short, the treatment with 0.5 μmol/L Mo is regarded as the most appropriate content to the growth, absorption, transformation, distribution and utilization of nitrate-nitrogen inMalushupehensisRehd. seedlings.
molybdenum;MalushupehensisRehd.;15N; absorption; utilization; distribution
2014-06-14 接受日期: 2014-06-30
現(xiàn)代農(nóng)業(yè)產(chǎn)業(yè)技術(shù)體系建設(shè)專項(xiàng)資金(CARS-28);公益性行業(yè)(農(nóng)業(yè))科研專項(xiàng)資金(201103003)資助。
劉利(1989— ),女,山東東平人,碩士研究生,主要從事果樹營(yíng)養(yǎng)診斷和施肥研究。E-mail: 15153871569@163.com * 通信作者 Tel: 0538-8242134,E-mail: scwei@sdau.edu.cn
S661.1.601; S143.7+1
A
1008-505X(2015)03-0727-07
