家蠅精氨酸激酶基因克隆及其在害蟲防治上的應用
2015-07-24 05:52:12于雪曹新茹高一夫唐婷柳峰松
河北大學學報(自然科學版) 2015年1期
于雪,曹新茹,高一夫,唐婷,柳峰松
(河北大學 生命科學學院,河北 保定 071002)
精氨酸激酶(arginine kinase,ATP:N-phosphotransferase EC2.7.3.3)廣泛存在于無脊椎動物如昆蟲、甲殼動物和軟體動物體內,是無脊椎動物能量代謝最重要的酶類之一.當動物體內的ATP充足時,可以將能量儲存在磷酸精氨酸的高能磷酸鍵中;當動物消耗大量的ATP時,精氨酸激酶催化磷酸精氨酸分解生成ATP.精氨酸激酶在無脊椎動物生命活動中發揮重要作用,是無脊椎動物特有的磷酸原激酶,該酶及其能量代謝途徑與哺乳動物不同,因此,精氨酸激酶可以作為防控害蟲的一個有效而安全的靶點.
RNA 干擾(RNA interference,RNAi)是指外源或內源雙鏈RNA(double strand RNA,dsRNA)介導的特異性降解靶向mRNA,導致轉錄后水平的基因沉默(post-transcriptional gene silencing,PTGS)[1].RNA干擾具有抑制轉座子活動、抗病毒入侵、調控基因表達等作用,是重要的監控機制.Guo等[2]利用反義RNA手段探索線蟲(Caenorhabditis elegans)par1基因的功能時發現了RNAi現象.Fire等[3]詳盡地解釋了這種現象是由于制備反義RNA 時混入了微量的dsRNA 引起的,認為dsRNA 是引發RNA 沉默的關鍵因子,并把這種現象稱為RNAi.目前,RNAi技術已經在多種昆蟲上得到應用,dsRNA 進入昆蟲體內的方法主要有投喂、浸泡、注射、轉基因和病毒介導等,這些方法各有特點,投喂法最具前景[4-8].
家蠅(Musca domestica)俗稱蒼蠅,其繁殖能力強,生長速度快,世界各地廣泛分布,與人類的關系密切.家蠅攜帶多種病原菌,并能將病原菌傳給人類或牲畜,危害性較大[9].本研究以家蠅精氨酸激酶基因為靶點,通過投喂長dsRNA 來抑制家蠅體內精氨酸激酶的表達水平,大大影響了家蠅的存活率,以此揭示精氨酸激酶在家蠅體內的生物學功能,并為害蟲防治提供了一種新思路.
1 材料與方法
1.1 試蟲與試劑
1.1.1 供試昆蟲
家蠅(Musca domestica)由本實驗室保種和飼養,飼養溫度為25 ℃,幼蟲餌料成分:麩皮55g,滅活酵母粉3g和150mL水.成蠅飼養成分:水、紅糖和奶粉.
1.1.2 菌株及質粒
大腸桿菌(Escherichia coli)DH5α菌株、HT115菌株以及L4440質粒.
1.2 主要試劑和試劑盒
RNAiso Plus,pMD18-T Vector,限制性內切酶,T4DNA 連接酶購自大連寶生物公司,M-MLV 反轉錄酶,SYBR Green,Taq DNA 聚合酶均為北京全式金生物技術有限公司產品,引物合成自蘇州金唯智生物技術有限公司.其他試劑均為國產或進口分析純.
1.3 總RNA 的提取及cDNA的合成
依照寶生物RNAiso Plus使用說明書,按步驟提取家蠅3齡幼蟲總RNA.核酸定量儀測定總RNA 純度和濃度,經瓊脂糖凝膠電泳檢驗完整性后,定總量為2μg的總RNA 為RT-PCR 模板,用通用引物Oligo(dT)反轉錄合成cDNA.
1.4 家蠅MdAK 基因的克隆
從本實驗室測得的家蠅轉錄組數據庫中搜索到一段長3 140bp的EST 序列(CL 939.Contig 1),通過Blastn程序在GenBank數據庫中對該序列進行同源性分析,發現與昆蟲精氨酸激酶基因相似性較高.根據此EST 序列用Primer premier 5.0設計正反向引物MdAK-F和MdAK-R(表1),以家蠅cDNA 為模板,進行PCR 擴增,PCR 反應條件為:94℃預變性5min;94℃變性30s,55℃退火40s,72℃延伸3min,30個循環;72 ℃延伸10min.PCR 產物經瓊脂糖凝膠電泳分離、切膠回收后,連接pMD18-T 載體,轉化至感受態細胞DH5α中,經菌落PCR 篩選陽性克隆后測序.
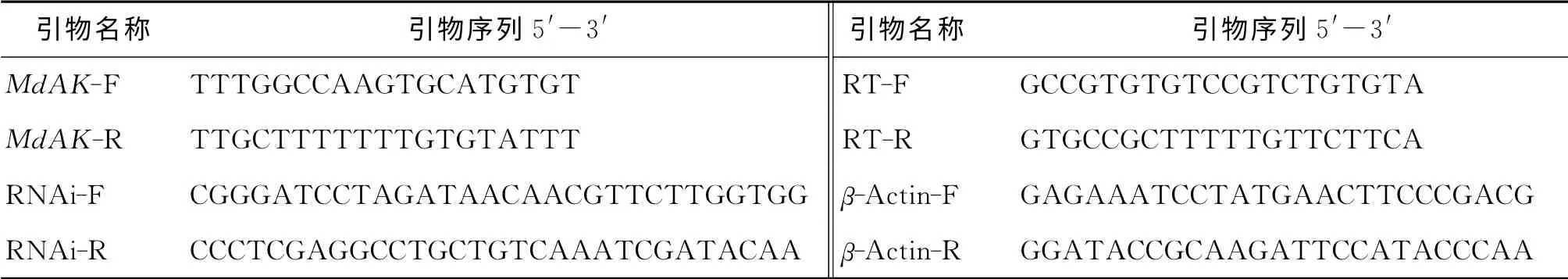
表1 PCR 引物Tab.1 Primers for PCR
1.5 MdAK 基因序列的生物信息學分析
應用Bioedit軟件搜索MdAKcDNA 序列開放閱讀框(open reading frame,ORF),并將核酸序列翻譯成氨基酸序列.在ExPASy:ProtParam 程序中(http://au.expasy.org/tools/protparam.html)分析MdAK的分子質量和等電點.利用Blastp程序在NCBI網站上進行同源序列查找,對獲得的昆蟲精氨酸激酶的氨基酸序列利用Clustal軟件進行多重序列比對.
1.6 L4440載體的構建和dsRNA的誘導
根據家蠅MdAK 的cDNA 序列,利用軟件Primer設計出上下游引物RNAi-F和RNAi-R,引物5'末端分別加入BamHI和XhoI限制性酶切位點,將擴增片段與L4440載體經過雙酶切回收后,連接獲得重組質粒L4440-MdAK,將其轉化至DH5α感受態細胞中,經過菌落PCR 篩選陽性克隆菌,提取小量質粒,重復雙酶切,檢驗后測序驗證.將構建好的干擾載體轉化至HT115宿主,通過菌落PCR 篩選陽性轉化子.陽性轉化子接種100mL含有氨芐青霉素(終質量濃度60μg/mL)和四環素(終質量濃度50μg/mL)的LB(Lysogeny Broth)新鮮液體培養基,37 ℃搖床200r/min培養至OD600=0.6~0.8時加入2mol/L IPTG(isopropyl-β-D-thiogalactoside)至終濃度為0.4mmol/L,37 ℃,200r/min誘導3h.同時,誘導表達含空載L4440質粒的菌作為空白對照.提取菌體RNA 進行電泳檢測,觀察dsRNA 的表達情況.
1.7 RNAi實驗
分別將經IPTG 誘導的含有L4440-MdAK 重組質粒和L4440 空載質粒的HT115 菌液稀釋至OD600=0.1,用以配置家蠅幼蟲餌料,80mL菌懸液加入60g麩皮,均勻攪拌,分別裝在玻璃瓶中.向每個玻璃瓶中投放200只剛剛孵化的家蠅1齡幼蟲,25 ℃培養箱中培養.之后每天觀察幼蟲數量變化并統計存活率,最后觀察幼蟲化蛹和羽化情況.每組實驗重復3次.
1.8 MdAK 基因敲低效果分析
投喂表達MdAK dsRNA 的菌液后,于6,24,36h后各取6只家蠅幼蟲,提取總RNA 并反轉錄,以βactin基因作為內參,進行實時熒光定量PCR 檢測MdAK mRNA 表達量變化.設計目的基因定量引物MdAK-RT-F/R;內參基因定量引物β-actin-F/R.實時熒光定量PCR 程序:95 ℃2 min;95 ℃20s,60 ℃30s,30個循環.實時熒光定量結果用相對定量2-ΔΔCt法進行分析,用t檢驗來比較實驗組和對照組的差異,P>0.05則為無顯著差異,0.01<P<0.05為顯著差異,P<0.01為極顯著差異.
2 實驗結果
2.1 MdAK 序列分析
通過RT-PCR 克隆得到一段全長為2 007bp的MdAKcDNA 序列,包含1 071bp的開放閱讀框,可編碼356個氨基酸殘基(圖1).利用ExPASy:ProtParam 程序分析,MdAK 蛋白理論分子質量為40.0ku,理論等電點為5.91.利用Blastp程序在NCBI網站上進行同源搜索,發現家蠅精氨酸激酶與黑腹果蠅(Drosophila melanogaster)的精氨酸激酶氨基酸序列相似性較高(identity=91%),于是將此基因命名為MdAK(Musca domesticaarginine kinase).精氨酸激酶氨基酸序列多重比對結果表明:不同物種的精氨酸激酶氨基酸中第271~277位的序列CPTNLGT,62 位天冬氨酸(Asp)和193 位精氨酸(Arg)保守性較高(圖2).CPTNLGT 是精氨酸激酶的活性中心序列,271位的半胱氨酸(Cys)是活性中心必需氨基酸.
2.2 RNAi處理后精氨酸激酶基因的表達變化
將PCR 擴增得到的MdAK 基因片段克隆到L4440載體,由于克隆位點兩側各含有1個T7啟動子,經IPTG 誘導后,插入片段被雙向轉錄并在大腸桿菌HT115中形成dsRNA.提取細菌總RNA 進行電泳檢測,發現L4440-MdAK 轉化菌株較L4440 轉化菌株明顯多出1 條亮帶(圖3),證明MdAK 雙鏈RNA 誘導成功.
利用投喂法對家蠅1齡幼蟲MdAK 基因進行了RNAi實驗,將能表達MdAKdsRNA 的工程菌和空載對照菌分別添加到幼蟲餌料中進行投喂.為了檢測MdAK 基因的敲低效果,對進行RNAi處理6,24,36h后的家蠅幼蟲體內MdAK 基因進行定量研究,結果發現RNAi處理組MdAK 基因表達被有效抑制,且在處理后36h的敲低程度達到80%以上(圖4).

圖2 昆蟲精氨酸激酶同源序列比對分析Fig.2 Multiple alignment of arginine kinases from insects
2.3 RNAi處理后家蠅存活率統計
RNAi處理家蠅幼蟲1,2,3,4,5,6d后,統計家蠅的數量(圖5).結果顯示,實驗組家蠅幼蟲在RNAi處理后第4天起開始大幅死亡,第6天家蠅的死亡率達到80%以上,而對照組的死亡率較低,不到10%.同時發現RNAi處理后實驗組家蠅化蛹率和羽化率也低于對照組,即使幼蟲存活至羽化為成蠅,生長速度也要比對照組慢很多,個體明顯要小于對照組,幼蟲尾部常常發黑,活動能力很差.
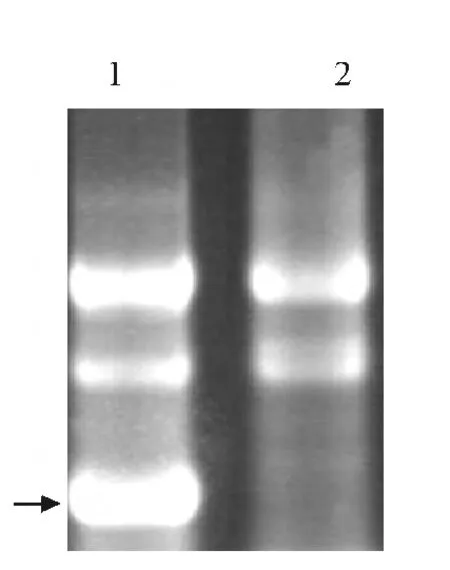
圖3 HT115總RNA電泳結果Fig.3 Total RNA electrophoretogram of HT115
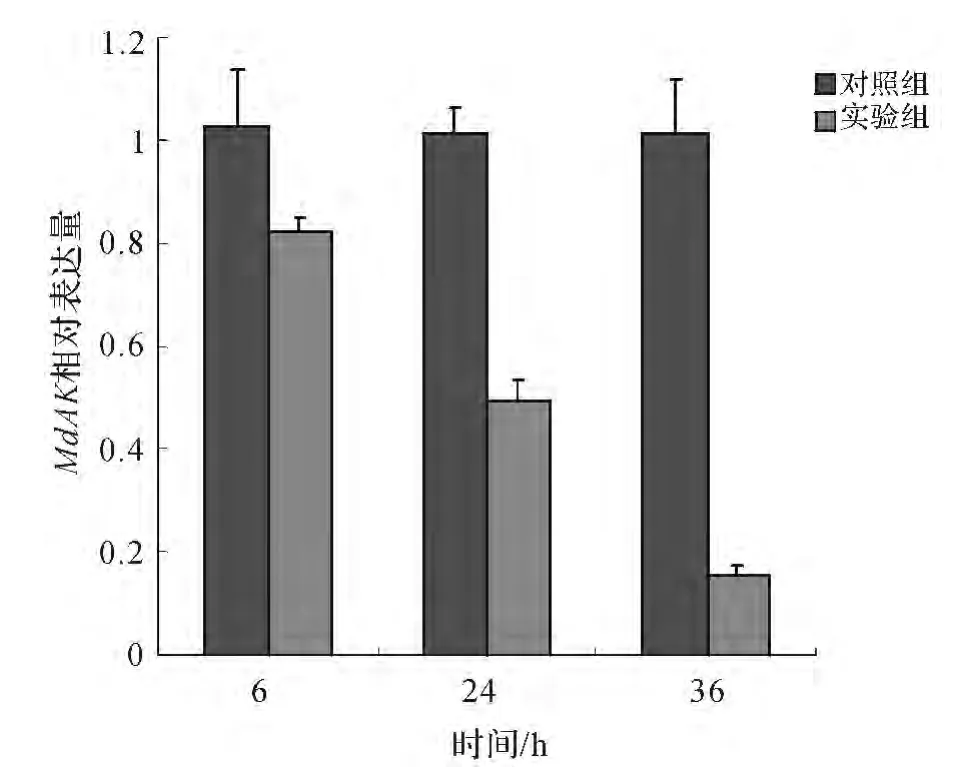
圖4 RNAi干擾對精氨酸激酶基因mRNA表達的影響Fig.4 Relative expression level of MdAK mRNA at different time points after RNAi treatment

圖5 RNAi處理后家蠅幼蟲的數量變化Fig.5 Changes in mortality rate of housefly after RNAi treatment
3 討論
精氨酸激酶是無脊椎動物體內主要的磷酸原激酶,其在無脊椎動物能量代謝上的重要作用已得到廣泛證實.本研究利用RT-PCR 技術獲得了長度為2 007bp的家蠅精氨酸激酶cDNA,編碼356個氨基酸.不同昆蟲的精氨酸激酶的氨基酸序列雖有不同,但有較高的同源性,相似性都在70%以上,這表明精氨酸激酶在進化上高度保守,同時也提示其功能的重要性.
精氨酸激酶是無脊椎動物體內不可缺少的、調節能量代謝的酶,且精氨酸激酶僅存在于無脊椎動物體內,哺乳動物體內肌酸激酶參與的代謝途徑與無脊椎動物體內精氨酸激酶參與的途徑不同.所以,精氨酸激酶可作為害蟲防治的潛在靶點.家蠅分布廣,繁殖能力強,免疫力強,生長周期短,適于實驗室研究.本研究,以家蠅作為實驗對象,通過投喂表達dsRNA 的工程菌來敲低家蠅體內精氨酸激酶基因的表達水平.大腸桿菌HT115為RNaseⅢ缺失突變菌株,有助于dsRNA 的穩定表達.基因定量結果顯示,投喂表達MdAK dsRNA 的大腸桿菌后,精氨酸激酶mRNA 表達水平明顯低于對照組,同時發現家蠅的生長發育受到了很大的影響,且幼蟲存活率顯著降低.
dsRNA 分子較單鏈RNA 分子相對穩定,噴灑到植物葉片上的dsRNA 能存留幾天[10-11].這樣使得RNAi技術具有潛力成為害蟲防治的有力工具.本研究證實dsRNA 可有效干擾家蠅體內精氨酸激酶基因的表達,并能有效殺死家蠅,且該方法僅對節肢動物等含有精氨酸激酶的生物有作用,不會影響其他生物的正常生長和發育,具有較高的安全性.化學農藥的使用給環境和人類健康帶來了不可估量的影響,尋求新的害蟲防治手段具有重要的意義.隨著昆蟲系統生物學和基因工程等學科的快速發展,針對害蟲的特有基因設計特異性藥物已經成為了害蟲生物防治的重要研究方向,具有廣闊的應用前景.
[1] 王子見 吳秀萍 鄧洪寬,等.RNA 干擾技術在動物寄生線蟲研究中的局限性[J].中國寄生蟲學與寄生蟲病雜志,2008,26(6):476-479.WANG Zijian,WU Xiuping,DENG Hongkuan,et al.Limitations of the application of RNA interference in the study of animal parasitic nematodes[J].Chinese Journal of Parasitology and Parasitic Diseases,2008,26(6):476-479.
[2] GUO Su,KEMPHUES K J.Par-1,agene required for establishing polarity in C.elegans embryos,encodes a putative Ser/Thr kinase that is asymmetrically distributed[J].Cell,1995,81(4):611-620.
[3] FIRE A,XU Siqun,MONTGOMERY M K,et al.Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans[J].Nature,1998,391(6669):806-811.
[4] BAUM J A,BOGAERT T,CLINTON W,et al.Control of coleopteran insect pests through RNA interference[J].Nat Biotechnol,2007,25(11):1322-1326.
[5] DZITOYEVA S,DIMITRIJEVIC N,MANEV H.Intra-abdominal injection of double-stranded RNA into anesthetized adult Drosophila triggers RNA interference in the central nervous system[J].Mol Psychiatry,2001,6(6):665-670.
[6] EATON B A,FETTER R D,DAVIS G W.Dynactin is necessary for synapse stabilization[J].Neuron,2002,34(5):729-741.
[7] TURNER C T,DAVY M W,MACDIARMID R M,et al.RNA interference in the light brown apple moth,Epiphyas postvittana(Walker)induced by double-stranded RNA feeding[J].Insect Mol Biol,2006,15(3):383-391.
[8] UHLIROVA M,FOY B D,BEATY B J,et al.Use of Sindbis virus-mediated RNA interference to demonstrate a conserved role of Broad-Complex in insect metamorphosis[J].Proc Natl Acad Sci U S A,2003,100(26):15607-15612.
[9] SCOTT J G,LIU N,KRISTENSEN M,et al.A case for sequencing the genome of Musca domestica(Diptera:Muscidae)[J].J Med Entomol,2009,46(2):175-182.
[10] TENLLADO F,BARAJAS D,VARGAS M,et al.Transient expression of homologous hairpin RNA causes interference with plant virus infection and is overcome by a virus encoded suppressor of gene silencing[J].Mol Plant Microbe Interact,2003,16(2):149-158.
[11] TENLLADO F,MARTINEZ-GARCIA B,VARGAS M,et al.Crude extracts of bacterially expressed dsRNA can be used to protect plants against virus infections[J].BMC Biotechnol,2003,3:3.
