棘托竹蓀提取液對單增李斯特菌的抑菌機理初步研究
2015-11-07 09:28:15藍蔚青潘迎捷孫曉紅
食品工業科技 2015年14期
關鍵詞:李斯特
藍蔚青,曹 奕,陳 燕,潘迎捷,孫曉紅
(上海海洋大學食品學院,上海水產品加工及貯藏工程技術研究中心,上海201306)
棘托竹蓀提取液對單增李斯特菌的抑菌機理初步研究
藍蔚青,曹奕,陳燕,潘迎捷,孫曉紅*
(上海海洋大學食品學院,上海水產品加工及貯藏工程技術研究中心,上海201306)
在細胞超微結構觀察的基礎上,采用實時熒光定量PCR技術初步分析棘托竹蓀提取液對單增李斯特菌的抑菌機理。使用最小抑菌濃度(15.0mg/mL)的棘托竹蓀提取液分別對單增李斯特菌菌液進行0.5、1.0、1.5h處理,提取對照組與處理組菌株的總RNA,通過反轉錄獲得各自cDNA,并選擇相關基因合成引物作Real-time PCR分析。以本菌的16S rRNA作為內參基因,以prfA、ActA、Iap為轉錄過程與表達蛋白基因,分析提取液對單增李斯特菌相關基因表達量的影響。由細胞超微結構觀察可知,單增李斯特菌經提取液處理后,菌體細胞的細胞壁變薄,細胞膜的完整性發生改變,內容物從胞內釋放出來。由熒光定量PCR法對不同處理時間下的相關基因相對表達量變化情況分析得出:處理組的prfA基因在0.5h時表達量有所上調,后期表達量上調速度減慢。1h后表達量基本停止上調,1.5h后,表達量轉為下調,說明提取液能在轉錄階段對單增李斯特菌的生命活動構成影響。ActA和Iap基因在處理0.5h后基因表達量上調,處理1h后上調量最大,ActA基因在1.5h后表達量上調量有所降低,Iap基因表達量出現下調,說明提取液能在轉錄水平上調控單增李斯特菌的生命活動,特別是多種毒力因子的表達過程。由此可知,棘托竹蓀提取液能對單增李斯特菌細胞的轉錄過程和抗性產生顯著影響,使菌體的細胞膜受損,細胞質溶出,從而導致菌體細胞死亡。
棘托竹蓀提取液,單增李斯特菌,抑菌機理
棘托竹蓀(Dictyophora echinovlvata)為鬼筆菌目鬼筆科竹蓀屬真菌,主要分布于云南、貴州、湖南、四川等地[1]。其子實體較小,菌蓋近鐘形,菌裙白色,菌柄較長,菌托白色或淺灰色,以7~8月為生長高峰期。棘托竹蓀因其營養豐富、香味濃郁、滋味鮮美等特點,長期以來作為我國珍貴特產之一,其價等黃金。近年來,國內外少數學者先后圍繞竹蓀的抗氧化[2]、免疫調節[3]、降壓降脂[4]、凝集保肝[5-6]、抑制癌細胞[7-8]等方面開展了部分研究工作,也對其抑菌活性進行了初步研究。其中檀東飛等[9-11]通過不同溶劑的竹蓀子實體浸提實驗,并采用水蒸氣蒸餾法提取其揮發油,用牛津杯法測定了其抑菌活性。盧惠妮等[12]通過瓊脂平板打孔法進行抑菌活性的測定,結果得出棘托竹蓀提取液能在中性乃至偏堿性條件下保持其抑菌活性,但關于棘托竹蓀抑菌機理方面的研究尚未開展。
單增生李斯特菌(Listeria monocytogenes)為食源性致病菌之一,在自然界和各類食品中經常被發現,具有高致病率的特點,且對外界環境具有很強的適應性[13],被WHO列為20世紀90年代主要食品致病菌,是21世紀對中國人衛生健康具有重大影響的12種病原微生物之一[14]。因此,采用合適的處理方式控制海產品中的致病菌,尤其是單增李斯特菌對于保證水產品的安全性具有重要的研究意義。目前,國內外已有相關學者對抑菌物質作用于菌體細胞的方式開展了相關方面的研究工作,在抑菌機理方面的表現主要有損傷細胞壁、改變細胞的透性、改變蛋白質和核酸分子、抑制酶的作用、作為抗代謝物與抑制核酸的合成等幾種方式[15]。本文在掃描電子顯微鏡(SEM)和透射電子顯微鏡(TEM)觀察提取液對細菌形態和結構作用效果的基礎上,通過實時熒光定量PCR法研究棘托竹蓀提取液對單增李斯特菌生命活動相關基因表達變化的影響,初步評價棘托竹蓀提取液對食源性致病菌的抑菌機理,以期為棘托竹蓀的后期開發利用提供理論依據。
1 材料與方法
1.1材料與儀器
棘托竹蓀子實體購自四川綿陽食用菌所,保存于干燥避光處;單增李斯特菌菌株編號:ATCC19115,購自中科院微生物研究所;胰蛋白胨大豆肉湯(TSB) 北京陸橋有限責任公司;TRIzol美國Invitrogen公司;酚:氯仿:異戊醇生工生物工程(上海)有限公司;異丙醇國藥化學試劑有限公司;DRR047A反轉錄試劑盒日本TaKaRa株式會社;Super Real Pre-mix熒光定量試劑盒天根生化科技有限公司。
E-1010離子濺射裝置、S3400N掃描電子顯微鏡、JEM2010型透射電子顯微鏡Hitachi日本株式會社;PCR儀、電泳儀美國Bio-Rad有限公司;5424離心機美國eppendorf公司;7500型Fast Real-Time PCR System美國Applied Biosystems公司;236HK高速冷凍離心機上海湘儀離心機儀器有限公司;超聲波細胞粉碎機寧波新芝生物科技股份有限公司。
1.2實驗方法
1.2.1棘托竹蓀提取液采用超臨界CO2萃取法(萃取壓力20MPa,萃取溫度35℃,萃取時間120min)獲得棘托竹蓀提取液,將其稀釋至15.0mg/mL(采用微量肉湯稀釋法結合平板涂布法確定其MIC,菌數保持在106CFU/mL,細菌生長受到明顯抑制的提取液最低濃度為MIC,微量肉湯稀釋法的MIC值為肉眼發現無明顯濁度變化的提取液最低濃度[16]),保存于-20℃冰箱備用。
1.2.2單增李斯特菌的活化取甘油管保藏的單增李斯特菌菌株,TSB活化兩次后用相應培養液進行稀釋,使菌液濃度達106CFU/mL。
1.2.3細胞超微結構觀察
1.2.3.1掃描電子顯微鏡(SEM)觀察取活化好的菌液,加入15.0mg/mL竹蓀提取液處理6h,以未作任何處理的菌液為對照組,兩組樣品同時進行處理。隨后,分別轉移至1.5mL離心管,4℃、7000r/min離心5min,棄上清,反復如上操作,多次離心獲得菌體,菌體沉淀用磷酸緩沖液洗滌3次后,用戊二醛固定、清洗、鋨酸固定12h,使用30%、50%、70%、90%的乙醇梯度脫水,將樣品涂至金屬箔片上,采用CO2臨界點干燥法置換乙酸乙酯作臨界點干燥固定、噴金,并置于掃描電鏡觀察。
1.2.3.2透射電子顯微鏡(TEM)觀察樣品前處理同上操作至戊二醛固定,取4℃預冷固定液1.5mL加入到每個離心管中,4℃固定10h以上,后續清洗、包埋、切片與制樣工作由華東師范大學電鏡中心協助完成。
1.2.4細菌相關基因表達量變化分析
1.2.4.1細菌總RNA提取參考鄒曉蕾等[17]采用Trizol法進行細菌總RNA的提取。收集菌液,以未做任何處理的菌液為對照組,采用15.0mg/mL棘托竹蓀提取液分別處理菌株0.5、1.0、1.5h。將處理組與對照組分別在4℃、12000r/min離心5min(沉淀加入3.0mg/mL的溶菌酶TE緩沖液100μL孵育10min),棄上清,加1.0mL Trizol,漩渦振蕩器振蕩15min,室溫靜置20min,加入0.2mL酚∶氯仿∶異戊醇(25∶24∶1),劇烈振蕩15s,靜置2min;4℃、12000r/min離心15min,獲取上清液;加入等體積預冷的異丙醇,4℃、12000r/min離心10min,棄上清。液體混勻,-20℃靜置20min;加入1.0mL、-20℃預冷的75%乙醇,洗滌沉淀。4℃,10000r/min離心5min,棄上清;晾干后加入30μL的焦磷酸二乙酯(DEPC)H2O溶解,分裝8.0μL用于后續操作,放置于-20℃,其余RNA放置在-80℃冰箱,測定其RNA濃度與純度。
1.2.4.2反轉錄將RNA濃度調至200ng/μL進行DNA去除及反轉錄,基因組DNA除去反應的配制體系為5×gDNA Eraser Buffer 2.0μL,gDNA Eraser 2.0μL,Total RNA 1.0μL,RNase Free dH2O 5.0μL,于PCR儀上進行DNA去除,隨后進行反轉錄。向10.0μL反應液中依次加入RNase Free dH2O 5.0μL和5×PrimeScript Buffer 2(for Real Time)3.0μL,混勻后再添加RT Primer Mix 1.0μL、PrimeScript RT Enzyme Mix I 1.0μL,并于PCR儀上進行反轉錄反應(反應條件:37℃、15min,85℃、5s,之后溫度維持在4℃),得到cDNA保存在-20℃冰箱中[17]。
1.2.4.3Real-Time PCR參照孫文爍等[18]選取20μL的反應體系進行Real-time PCR分析。其中,熒光染料10.0μL、正向引物(10μmol/L)1.5μL、反向引物(10μmol/L)1.5μL、cDNA模板2.0μL與RNase-free ddH2O 5.0μL,輕柔混勻并短暫離心。將反應體系置于熒光定量PCR儀中進行Real-Time PCR兩步法反應,PCR反應條件為,第一階段:95℃、15min;第二階段:95℃、10s與60℃、32s,進行40個循環,所需引物序列如表1所示。
熒光定量PCR反應結果以2-△△Ct值作為衡量表達量變化情況的指標,其代表實驗組目的基因的表達相對于對照組的變化倍數,使用這一方法便可直接得到目的基因相對于內參基因的量。若該值大于1,則說明特征基因表達量上調;若該值小于1,則說明特征基因表達量下調[22]。
1.2.4.4數據處理實驗數據均采用3次平行實驗的平均值,數據用軟件origin(Pro)7.5繪圖,數據間的差異通過統計軟件SPSS 13.0中的Duncan新復極差法進行方差分析與多重比較,結果以平均值±標準偏差表示。

表1 Real-time PCR所需引物序列Table 1 Primers sequences used for Real-time PCR
2 結果與分析
2.1細胞超微結構觀察結果
2.1.1掃描電子顯微鏡(SEM)觀察結果從圖1(a)中可以看到單增李斯特菌對照組的細菌細胞形態正常飽滿,細胞膜結構完整;圖1(b)中對照組的細胞正在進行分裂繁殖,輪廓清晰可見。由圖1(c)可看出:經棘托竹蓀提取液處理后,視野內殘留的菌體細胞變少且不能維持細胞的原有形態;從圖1(d)可見,處理后的菌體細胞形狀變得松散,胞內物質發生泄漏。說明在該菌經MIC竹蓀提取液處理后,已對其正常生長產生明顯的抑制作用。
2.1.2透射電子顯微鏡(TEM)觀察結果從圖2(a)發現,同掃描電子顯微鏡對照組的情況一致,透射電鏡視野下對照組的菌體細胞較多,形態完整,內容物充實,可明顯看出細胞正進行分裂而形成的縊痕。從圖2(b)中可以看到,經提取液處理后,細胞的細胞壁變薄,幾乎觀察不到完整的細胞膜,且有內容物從胞內滲透出來。

圖1 單增李斯特菌掃描電子顯微鏡觀察結果Fig.1 Scanning electron microscopy micrographs of Listeria monocytogenes
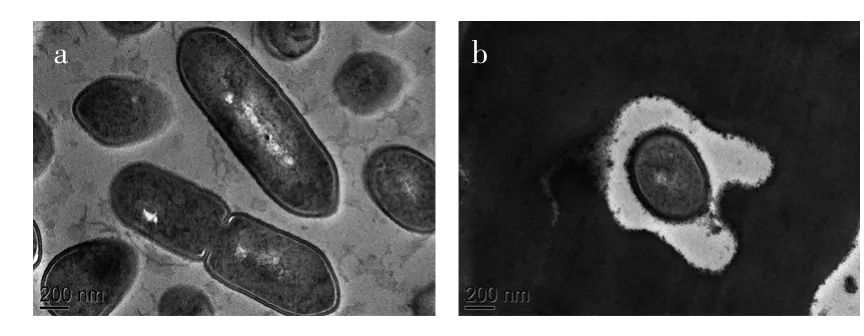
圖2 單增李斯特菌透射電子顯微鏡觀察結果Fig.2 Transmission electron microscopy micrographs of Listeria monocytogenes
2.2熒光定量PCR反應結果分析
prfA是單增李斯特菌中的重要基因之一,其所表達的轉錄激活因子能夠激活細胞內多類毒力因子的表達[23],如ActA蛋白和Iap基因編碼的p60蛋白,其表達量的上調表示單增李斯特菌細胞生命活動旺盛。單增李斯特菌的特征基因相對表達量變化結果如圖3所示。
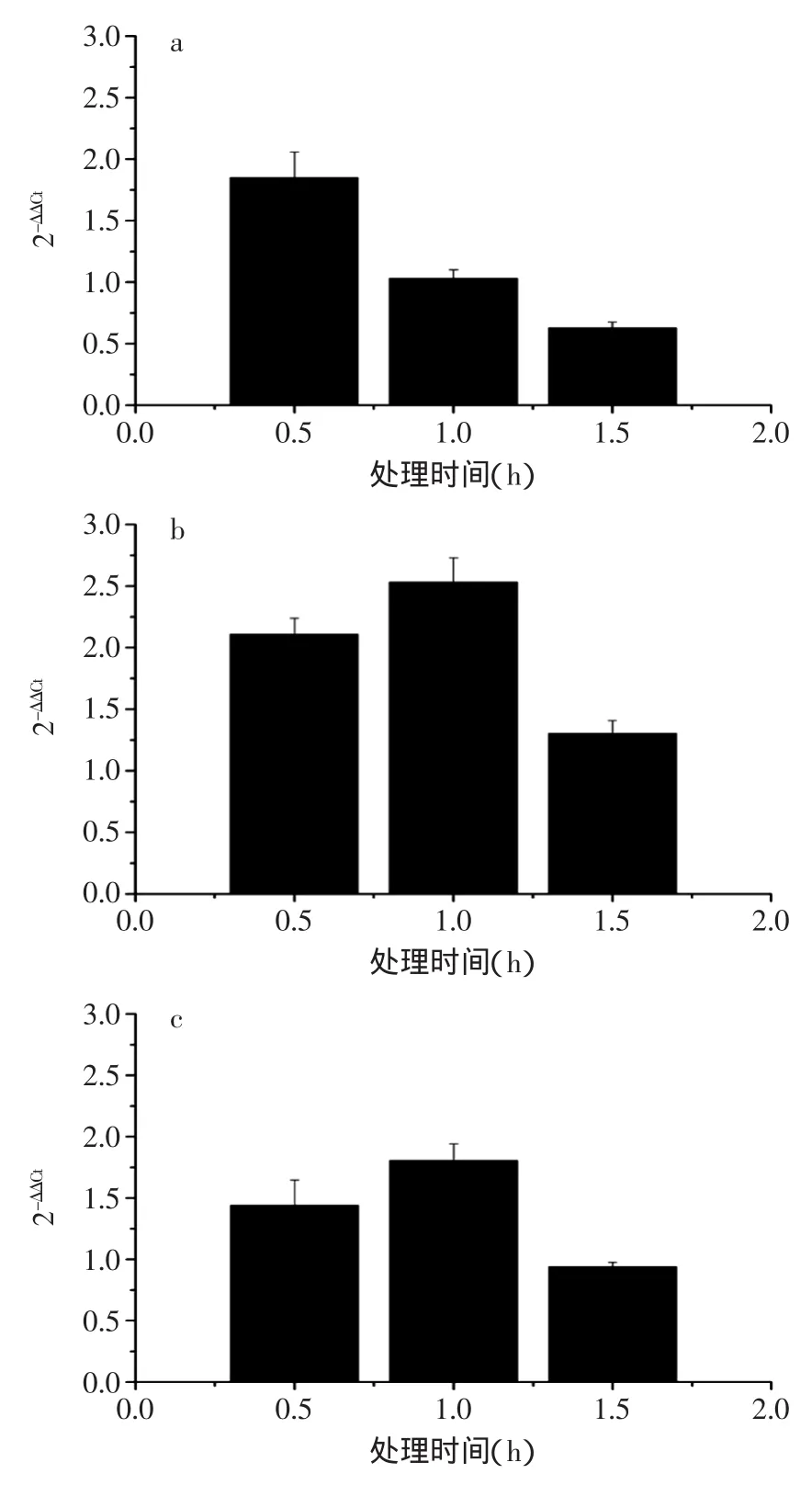
圖3 單增李斯特菌相關基因相對表達量變化Fig.3 Relative expression levels of several related genes resistance of Listeria monocytogenes
從圖3中可以看出,prfA基因在0.5h時表達量有所上調,隨著處理時間延長,其表達量上調速度減慢。處理1h后表達量基本停止上調,處理1.5h后,其表達量轉為下調。ActA基因在處理0.5h后較處理前基因表達量上調,處理1h后上調量達到最大,處理1.5h后表達量上調量有所降低。Iap基因在0.5h與1h的基因表達量變化與ActA基因基本相似,但在1.5h后表達量呈下調趨勢。可見,單增李斯特菌經棘托竹蓀提取液處理后,已對菌體的基因表達產生影響,對其生長產生抑制。而表達量的下調則說明竹蓀提取液能通過在轉錄水平上調控單增李斯特菌的生命活動,抑制菌體的活性。但竹蓀提取液的作用發揮仍需一段時間,因此才會出現prfA表達量在0.5h時仍處于上調的現象。
3 結論
通過進行細胞超微結構觀察結果顯示,提取液處理后,視野內殘留的菌體細胞已經很少,不能維持細胞的原有形態,菌體細胞形狀變得松散,細胞的細胞壁變薄,幾乎觀察不到完整的細胞膜,胞內物質泄漏。說明在該菌經MIC竹蓀提取液處理后,已對其正常生長產生明顯的抑制作用。單增李斯特菌經棘托竹蓀提取液處理后,prfA基因表達量的上調速度逐漸減慢,在1h后由上調轉為下調,說明提取液對其基因表達產生影響,對其正常生長產生抑制作用。而在其表達量轉為下調后,ActA和Iap基因表達量的上調速度也迅速下降,說明竹蓀提取液能夠通過在轉錄水平上調控單增李斯特菌的生命活動,特別是多種毒力因子的表達過程。然而,竹蓀提取液從處理到開始發揮作用,需要一段時間,細胞可能會對其產生一定抗性,從而使prfA表達量在0.5h時處于上調階段。
由實驗可知,棘托竹蓀提取液對單增李斯特菌細菌轉錄水平和能量代謝的影響尤為明顯,對細胞膜的破壞能力也較強。其特性與同為植物源天然產物的烏梅提取液[24]、藍莓提取物[25]較為類似,這也可在一定程度上說明了植物源天然產物的抑菌作用機制基本相同。課題組還將在后期進一步開展研究工作,以深入探究棘托竹蓀提取液對食源性致病菌的抑菌機理。
[1]劉振祥,張勝.食用菌栽培技術[M].北京:化學工業出版社,2007:122-127.
[2]江玉姬,王宏雨,謝寶貴,等.食用菌的抗氧化活性及竹蓀抗氧化物質提取工藝的優化[J].熱帶作物學報,2011,32(1):76-78.
[3]MalaguarneraL,FerlitoL,LmbesiRM, etal. Immunosenescence:a review[J].Archives of Gerontology and Geriatrics,2001,32(1):1-14.
[4]林海紅,林浪.長裙竹蓀對大鼠血脂的影響[J].福建農業大學學報,2000,29(2):238-241.
[5]林玉滿,蘇愛華.棘托竹蓀凝集素的純化及其生化特性[J].植物資源與環境學報,2004,13(3):1-6.
[6]蔡美珠,唐礡.竹蓀托蓋液對小鼠肝、腎功能的影響[J].醫學理論與實踐,2004,17(5):497-498.
[7]杜昱光,白雪芳,卜宗式,等.竹蓀深層發酵菌絲體對小鼠免疫功能的影響及其抗腫瘤活性的影響[J].中國食用菌,1998,17(5):25-26.
[8]N Kodama,K Komuta,H Nanba.Effects of maitake(Grifola frondosa)D-fraction on the activation of NK cells in cancer patients[J].Journal of Medicinal Food,2003,6(4):371-377.
[9]檀東飛,黃儒珠,盧真,等.棘托竹蓀菌托的化學成分及抑菌活性研究(I)[J].菌物學報,2006,25(4):603-610.
[10]檀東飛,黃儒珠,盧真,等.棘托竹蓀菌蓋的化學成分及抑菌作用研究(II)[J].微生物學雜志,2007,27(6):8-12.
[11]檀東飛,黃儒珠,盧真,等.棘托竹蓀子實體鮮品的化學成分及抑菌活性研究[J].福建師范大學學報:自然科學版,2010,26(2):100-105.
[12]盧惠妮,潘迎捷,孫曉紅,等.棘托竹蓀子實體抑菌活性的研究[J].食品科學,2009,30(15):120-123.
[13]Vazquez-Boland JA,Kuhn M,Berche P,et al.Listeriapathogenesis and molecular virulence determinants[J].Clinical Microbiology Reviews,2001,14(3):584-640.
[14]孫會芳.單核細胞增生性李斯特桿菌單克隆抗體的制備[D].蘭州:西北民族大學,2007.
[15]小佩爾扎MJ,里德RD,詹ECS.微生物學[M].武漢大學生物系微生物教研室譯.北京:科學出版社,1987:359-361.
[16]曹奕,孫曉紅,陳燕,等.微量肉湯稀釋法測定長裙竹蓀多種提取液對食品中常見細菌的抑制效果[J].食品工業科技,2013(1):16-20.
[17]鄒曉蕾,劉禮崔,羅立新.細菌總RNA提取方法的比較[J].現代食品科技,2013,30(7):1948-1954.
[18]孫文爍,靳夢曈,王敬敬,等.運用Real time PCR建立即食蝦中副溶血性弧菌分子預測模型[J].現代食品科技,2014,30(7):142-148.
[19]Maria A O,Eliana G R,Alzira M B,et al.Quantification of Listeria monocytogenes in minimally processed leafy vegetables using a combined method based on enrichment and 16SrRNA real-time PCR[J].Food Microbiology,2010(27):19-23.
[20]Peter R,Martina K,Martin W,et al.Detection of Listeria monocytogenes in food using a combined enrichment/real-time PCR method targeting the prfA gene[J].Research in Microbiology,2006(157):763-771.
[21]Xiaohui Z,Xinan J.Polymerase chain reaction detection of Listeria monocytogenes using oligonucleotide primers targeting actA gene[J].Food Control,2005(16):125-130.
[22]Kim B R,Nam H Y,Kim S U,et al.Normalization of reverse transcription quantitative-PCR with housekeeping genes in rice[J].Biotechnology Letters,2003,25(21):1869-1872.
[23]王海艷,劉中學,石新華,等.單增李斯特菌及其表面蛋白的研究進展[J].檢驗檢疫科學,2006,16(2):76-80.
[24]耿飛,王偉,周濤.烏梅提取液對李斯特菌的抑菌機理[J].食品科學,2012,32(15):88-93.
[25]謝慶超.藍莓提取物的抑菌效果及其抑制副溶血性弧菌機理的研究[D].上海:上海海洋大學,2012.
Preliminary research on the antimicrobial mechanism of Dictyophora echinovolvata extracts against Listeria monocytogenes
LAN Wei-qing,CAO Yi,CHEN Yan,PAN Ying-jie,SUN Xiao-hong*
(College of Food Science and Technology Shanghai Ocean University,Shanghai Engineering Research Center of Aquatic Product Processing and Preservation,Shanghai 201306,China)
Based on the observation of cell ultrastructure,the method of real-time fluorescent quantitative PCR was used to evaluate the antimicrobial mechanism of Dictyophora echinovolvata extracts against Listeria monocytogenes.The strain was treated with 15.0mg/mL(minimum inhibitory concentration,MIC)of Dictyophora echinovolvata extracts under 0.5,1.0 and 1.5h.Total RNA of the control and treated group strains were extracted,cDNA were obtained by reverse transcription and the relevant gene primers were selected for Real-time PCR method respectively.The strains of 16S rRNA as reference gene,prfA,ActA and Iap as feature genes.It was showed that the cell wall become thinner,the integrity of cell membrane was changed and the contents released from intracellular when treated with Dictyophora echinovolvata extracts by the ultrastructure of cell observations.Compared with the control group,the level of gene expression in prfA was up-regulated at 0.5h,the speed of up-regulated became slower at 1h,the trend of up-regulation stopped at 1.5h with the prolong of treatment time,which proved that extracts could affect the activities of Listeria monocytogenes in transcription. While the level of gene expression in ActA and Iap were up-regulated in 0.5h,the amount of up-regulated expression was increased fast at 1h.The level of gene expression in ActA was decreased at 1.5h and that in Iap was down-regulated,which showed that extracts could regulate the level of transcription in Listeria monocytogenes,especially on the expression of virulence factors.Dictyophora echinovolvata extracts had a significant impact on Listeria monocytogenes for the process of transcription and the resistant of bacteria,which could make the damage of cell membrane and the dissolution of cytoplasm leading to the death of cell.
Dictyophora echinovolvata extracts;Listeria monocytogenes;antibacterial mechanism
TS201.1
A
1002-0306(2015)14-0152-05
10.13386/j.issn1002-0306.2015.14.023
2014-12-01
藍蔚青(1977-),男,博士,高級工程師,研究方向:水產品低溫保鮮技術研究。
孫曉紅(1978-),女,博士,副教授,研究方向:天然產物研究與開發。
上海市教育委員會科研創新項目(11YZ159);上海市科技興農重點攻關項目(滬農科攻字(2015)第4-12號);上海海洋大學科技發展專項基金(A2-0209-15-200061)。
猜你喜歡
藝術評鑒(2022年6期)2022-05-10 10:07:27
樂府新聲(2019年2期)2019-11-29 07:34:30
幼兒園(2019年7期)2019-09-05 17:49:18
北方音樂(2017年4期)2017-05-04 03:40:15
中國音樂教育(2015年8期)2015-05-16 03:22:06
小小說月刊(2013年6期)2013-05-14 14:55:19
食品科學(2013年23期)2013-03-11 18:30:07
食品科學(2013年23期)2013-03-11 18:30:05
食品科學(2013年15期)2013-03-11 18:25:40
食品科學(2013年15期)2013-03-11 18:25:25
