灰平鏈霉菌Streptomyces griseoplanus S501產外切菊粉酶的分離純化及酶學性質研究*
2015-12-16 08:06:32于春于基成張春紅劉秋陳嬌
食品與發(fā)酵工業(yè) 2015年8期
關鍵詞:影響
于春,于基成,張春紅,劉秋,陳嬌
1(沈陽農業(yè)大學食品學院,遼寧沈陽,110866)2(大連民族大學生命科學學院,遼寧大連,116600)
菊粉是由多個D-果糖分子通過β-2,1果糖糖苷鍵連接而成的多聚果糖,末端有1個葡萄糖殘基以α-2,1糖苷鍵與之相連,呈直鏈結構,聚合度為2~60[1]。菊粉酶又稱果聚糖酶(β-2,1-D-果聚糖水解酶),其催化水解菊粉中β-2,1呋喃果糖苷鍵,酶解產物為低聚果糖和果糖,在工業(yè)上有重要的應用價值[2]。直接利用粗酶液來生產低聚果糖或果糖時,常常由于菊粉酶性質不穩(wěn)定或酶活力較低而影響低聚果糖或果糖產率。此外,菊粉酶粗酶液在使用過程中還常存在易變質和不易保存等問題[3-6]。針對這些問題,研究者通常采用酶的固定化來解決。但是在酶的固定化過程中,由于一些雜蛋白的存在而影響酶的活力和酶的固定化效果[7-8]。因此,研究菊粉酶的分離純化并獲得較高純度的菊粉酶,對進一步了解其酶學性質和研究其固定化方法具有重要意義。
本實驗以從分離自鴨綠江濱海濕地環(huán)境中的1株產酶活性高、穩(wěn)定性好的灰平鏈霉菌S.griseoplanus S501為研究對象,采用固態(tài)發(fā)酵法制備菊粉酶粗酶液,結合不同的分離純化方法獲得純酶組分,并對其酶學性質進行了研究。
1 材料與方法
1.1 材料
菊粉(大連佐源集團公司提供);灰平鏈霉菌S.griseoplanus S501(CGMCC No.8185)由大連民族大學功能微生物項目組保藏;所用試劑均為分析純。
1.2 固態(tài)發(fā)酵培養(yǎng)基
麩皮:833.33 g/L,營養(yǎng)液:KNO30.83 g/L,NaCl 0.42 g/L,MgSO4·7H2O 0.42 g/L,K2HPO4·3H2O 0.42 g/L,FeSO4·7H2O 0.009 g/L,去離子水1 L,pH值自然。
1.3 粗酶液的制備
菌株S501經固態(tài)發(fā)酵后,按1∶5(g∶mL,以發(fā)酵基質干重計)加入去離子水,振蕩提取1 h,4℃ 8 000 r/min離心20 min,上清液即為粗酶液。
1.4 菊粉酶活力的測定
[9]方法測定。
1.5 菊粉酶的分離純化
1.5.1 硫酸銨分級沉淀
將粗酶液置于冰水浴中,在磁力攪拌下,緩緩加入硫酸銨至質量濃度為200 g/L,4℃靜置3~5 h,8 000 r/min離心20 min,除雜蛋白。上清液再添加硫酸銨至濃度為900 g/L,4℃ 8 000 r/min離心20 min,沉淀用0.1 mol/L pH5.6的醋酸緩沖液溶解,4℃對去離子水透析,每12 h換透析液1次,共6次。透析后,冷凍干燥即為粗酶。
1.5.2 Sephadex G-75凝膠層析
將1.5.1中制得粗酶采用Sephadex G-75凝膠層析柱(2.0 cm×30.0 cm)分離。上樣量2.0 mL,洗脫液為0.02 mol/L pH5.6的醋酸緩沖液,流速0.2 mL/min,監(jiān)測波長280 nm,每30 min收集1管洗脫液。測定各管收集液活性,將具有酶活性峰洗脫液收集備用。
1.5.3 DEAE-Sepharose CL-6B離子交換色譜
將1.5.2收集到的具有酶活性的收集液采用DEAE-Sepharose CL-6B離子交換色譜柱(2.0 cm×40.0 cm)分離。上樣量2.0 mL,采用0.0~0.5 mol/L NaCl和0.02 mol/L pH5.6的醋酸緩沖液進行線性梯度洗脫,流速為1.0 mL/min,監(jiān)測波長280 nm,每管收集5.0 mL。同時測定其活力,收集具有酶活性峰洗脫液備用。
1.5.4 菊粉酶蛋白純度和分子量測定
采用SDS聚丙烯酰胺凝膠電泳(SDS-PAGE)測定分離獲得酶的純度和分子量。分離膠和濃縮膠濃度分別為15%和5%。
1.6 菊粉酶酶學性質研究
1.6.1 菊粉酶的最適溫度及熱穩(wěn)定性
最適溫度測定:以20 g/L菊粉溶液為底物,定量加入酶液,分別在 20、30、40、50、60、70、80 ℃下反應30 min后測定酶活。以最高酶活性組為100%,以相對酶活考察菊粉酶的最適溫度。
熱穩(wěn)定性測定:酶液在不同溫度下預保溫1 h后,迅速冷卻到室溫后,分別測定殘余酶的相對活力,以未經保溫處理的酶液活性為100%。計算其相對酶活。

1.6.2 菊粉酶的最適pH值及酸堿穩(wěn)定性
最適pH值測定:分別以pH 3.8~5.8的0.2 mol/L的醋酸緩沖溶液配制2%的菊粉底物溶液,定量加入酶液,50℃下反應30 min后測定酶活,計算相對酶活。
酸堿穩(wěn)定性測定:采用不同pH值緩沖溶液定量稀釋酶液,4℃放置過夜,測定其殘余酶的相對酶活。
1.6.3 底物濃度對菊粉酶活力的影響
用0.2 mol/L pH5.0的醋酸緩沖液分別配制濃度為20、40、60、80 和100 g/L 的菊粉溶液,定量加入酶液,50℃下反應30 min后,測定酶活,計算相對酶活。
1.6.4 金屬離子對菊粉酶活力的影響
用0.2 mol/L pH5.0的醋酸緩沖液分別配制5.0 mmol/L 的 Ba2+、Cu2+、Zn2+、K+、I-、Mg2+、Al3+、Ca2+、Li+、Ag+、Fe2+、Fe3+溶液,以 40 g/L 的菊粉溶液為底物,定量加入酶液,50℃下反應30 min后測定酶活,以未加金屬離子的底物溶液為空白,考察金屬離子對菊粉酶活力的影響。
1.6.5 反應時間對菊粉酶活力的影響
以4%的菊粉溶液為底物,定量加入酶液,50℃下分別反應 5、10、15、20、25 和 30 min 后,測定其的酶活,以酶解產物還原糖含量對反應時間作圖。
1.6.6 菊粉酶的動力學常數測定
分別以不同濃度的菊粉溶液為底物,定量加入酶液,50 ℃下,分別反應 5、10、15、20、25 和 30 min,計算底物濃度[s]和反應速度[v],以 1/v對 1/[s]作圖,求算以菊粉為底物時菊粉酶的動力學常數Km和最大反應速率vmax。
2 結果與討論
2.1 菊粉酶的分離純化
2.1.1 硫酸銨分級沉淀
由表1可以看出,粗酶液經不同飽和度的硫酸銨沉淀后,其上清液相對酶活性差異顯著。隨著硫酸銨飽和度的增加,上清液相對酶活性逐漸減低,當硫酸銨飽和度達到900 g/L時,上清液殘余酶活性為9.43%,說明超過90%的酶通過沉淀被分離;而當硫酸銨飽和度為200 g/L時,其上清液相對酶活性為92.35%,則表明大部分具有活性的菊粉酶仍留在上清液中。因此,菊粉酶粗提純時先采用飽和度為200 g/L硫酸銨進行沉淀去除雜蛋白后,將上清液中硫酸銨飽和度調至900 g/L,沉淀后可得粗酶。
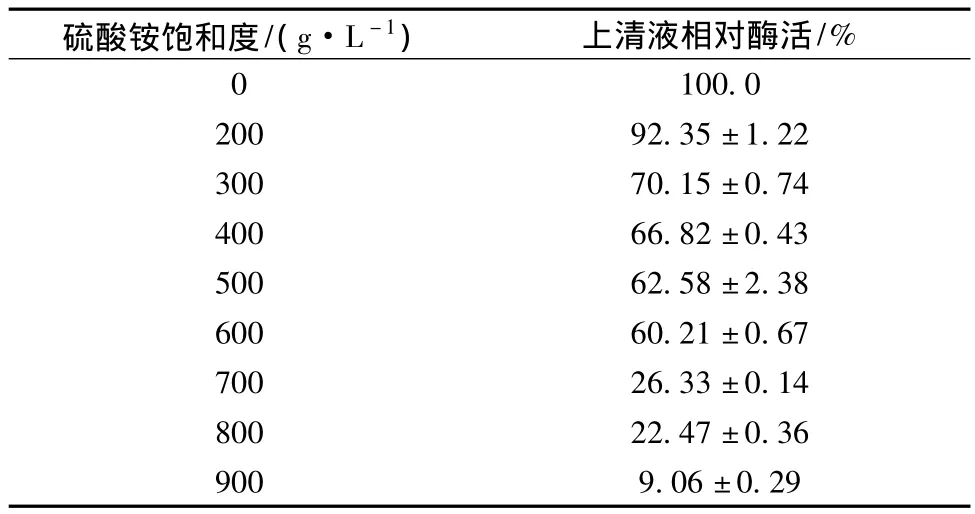
表1 不同濃度硫酸銨沉淀后上清液相對酶活(n=3)Table 1 The relative enzyme activity of supernatant after precipitated by different concentrations of ammonium sulfate(n=3)
2.1.2 Sephadex G-75凝膠層析
由洗脫曲線(圖1)可見,硫酸銨分級沉淀分離的粗酶經Sephadex G-75葡聚糖凝膠層析分離后,共獲得5個組分。收集各個組分并測定其酶活性發(fā)現,其中1號和2號峰具有酶活性,活性分別為57.5 U/mg和254.9 U/mg,合并二者酶液作為下一步離子交換色譜法分離和電泳實驗樣品。
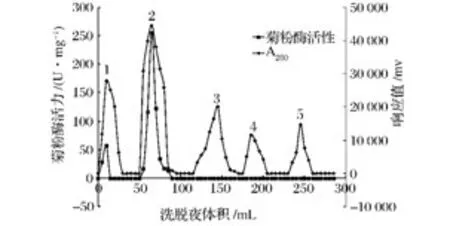
圖1 菊粉酶的Sephadex G-75葡聚糖凝膠柱層析圖Fig.1 Gel filtration chromatography of inulinase on Sephadex G-75 column
2.1.3 粗酶液的DEAE-Sepharose CL-6B離子交換色譜分離
據文獻報道,菊粉酶的等電點一般為4.0~5.0[10-18],因此,選用陰離子交換柱對其進行進一步的純化。由圖2可以看出,經DEAE-Sepharose CL-6B離子交換色譜分離后,共獲得3個組分。雖然組分2和組分3并沒有完全分離,但經活性測定后發(fā)現,這2個組分峰收集液無酶活性。而組分1則具有較強的外切菊粉酶活性,活性達到483 U/mg,因此,收集組分1峰濃縮備用。
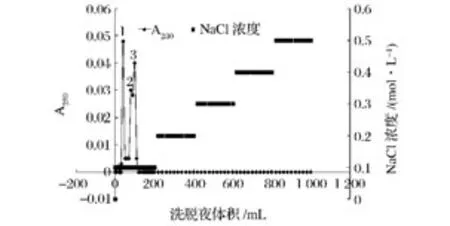
圖2 菊粉酶的DEAE-Sepharose CL-6B離子交換色譜層析圖Fig.2 Ion exchange chromatography of inulinase on DEAE-Sepharose CL-6B
2.1.4 不同純化方法獲得酶組分的酶活和比酶活測定
菌株S501產外切菊粉酶的分離純化結果見表2。由表2可知,發(fā)酵液經過硫酸銨沉淀、Sephadex G-75凝膠柱、DEAE-Sepharose CL-6B離子柱分離純化后,比酶活和純化倍數逐漸增加。3種方法獲得酶組分活性和純化倍數分別為27.48、46.11、249.5 U/mg和 1.15、1.93、10.42。

表2 菌株S501產外切菊粉酶的分離純化結果(n=3)Table 2 Summary of inulinase purification(n=3)
2.1.5 菊粉酶純度和分子量測定
采用SDS-PAGE分別測定了粗酶液和硫酸銨分級沉淀法、Sephadex G-75凝膠層析法、DEAE-Sepharose CL-6B離子交換色譜法分離后的粗酶純度和分子量。由圖3可見,未經處理粗酶液約有10個條帶,經Sephadex G-75分離獲得5條組分帶,而經DEAESepharose CL-6B離子交換色譜柱分離后的酶液僅有1條條帶,說明采用DEAE-Sepharose CL-6B柱分離后已獲得純度較高的外切菊粉酶。經計算其分子質量約為23.442 kDa。與文獻報道比較,該酶的分子質量較小,這可能與不同菌株所產的菊粉酶在分子結構和組成方面存在差異有關;另一方面,SDS-PAGE技術只能粗略估算蛋白分子量。
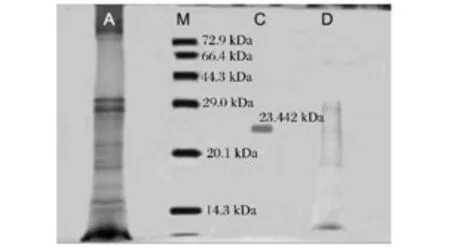
圖3 菊粉酶的SDS-PAGE電泳圖Fig.3 Electrophoretogram of SDS-PAGE of inulinase
2.2 菊粉酶的酶學性質
2.2.1 菊粉酶的最適溫度及熱穩(wěn)定性
溫度對酶反應速度的影響非常復雜。溫度既影響到酶蛋白的構象,參與酶促反應功能團的解離狀態(tài),也影響酶與底物的親和力,酶-底物絡合物的分解;甚至還影響酶與激活劑、抑制劑的親和力等[19]。
由圖4可以看出,隨著反應溫度的增加,菊粉酶的活性先增加后減小,在50℃時達到最高(464.0 U/mg),說明該菊粉酶的最適溫度為50℃。這一結果與張國青等人[20-24]報道結果相符合。而圖5結果進一步證明,將菊粉酶在不同溫度下預保溫1 h,迅速冷卻至室溫,于50℃下反應30 min后測定其酶活,其酶活力在50℃時達到最高。

圖4 溫度對菊粉酶活力的影響Fig.4 Effects of different temperature on inulinase activity
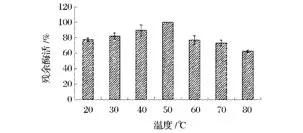
圖5 菊粉酶的熱穩(wěn)定性Fig.5 Effects of different temperature on inulinase stability
2.2.2 菊粉酶的最適pH值及酸堿熱穩(wěn)定性
最適pH是酶的一個很重要的特征,對酶的催化反應速率影響很大,主要表現為:(1)通過影響酶的空間結構來影響酶的穩(wěn)定性;(2)通過影響酶活性中心的催化基團的解離來影響酶與底物的結合;(3)通過影響底物的解離來影響其與酶的結合。酶的最適pH與酶的種類、底物濃度及種類、緩沖液種類等條件有關,它并不是一個常數,會隨著反應條件的變化而變化。因此,酶的最適pH只是在某一特定條件下的一個確定的數值。pH的穩(wěn)定性則是表明酶液在不同pH 下的耐受程度[25-26]。
由圖6可以看出,隨著體系pH值的增加菊粉酶的活性變化較為顯著,在pH5.0時酶活達到最高(448 U/mg)。為了進一步驗證這一實驗結果,研究中采用不同pH值的0.2 mol/L醋酸緩沖溶液配制同濃度酶液,室溫下放置1 h后,測定其殘余酶的相對活性。結果發(fā)現,該菊粉酶活性仍在pH=5.0時活性最高(圖7),因此,說明該菊粉酶的最適pH值為5.0。
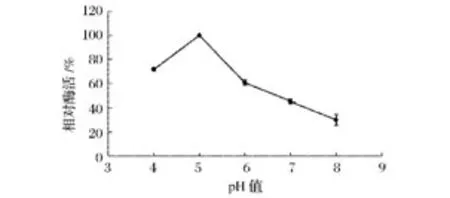
圖6 不同pH值對菊粉酶活力的影響Fig.6 Effect ofdifferent pH on inulinase activity
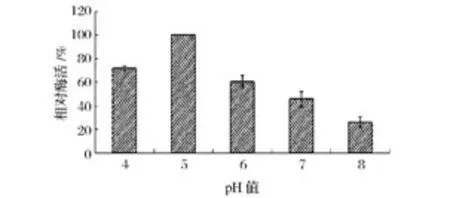
圖7 菊粉酶的酸堿穩(wěn)定性Fig.7 Effects of different pH on inulinase stability
2.2.3 底物濃度對菊粉酶活力的影響
底物濃度對酶促反應具有顯著的影響。當底物濃度較低時,提高底物濃度可以提高反應速度;但當底物濃度較高時,底物濃度進一步提高,反應速度的增加變得緩慢甚至不再增加。由圖8可以看出,隨著底物濃度的增加,菊粉酶活力先增加后趨于平穩(wěn),說明當酶濃度一定時,繼續(xù)增加底物濃度對產物影響不顯著。也進一步說明在酶促反應中酶濃度與底物濃度互相影響的關系。因此,研究中將菊粉酶的最適底物濃度設定為40 g/L。
2.2.4 金屬離子對菊粉酶活力的影響
金屬離子主要影響酶活性中心部位,從而影響酶的反應速率。金屬離子對酶活性的影響表現為一為活化或激活作用,一為抑制作用[27]。圖9結果表明,Mg2+、I-、Li+、Fe3+、Al3+、K+對該菊粉酶有很強的抑制作用,Cu2+、Ca2+、Ag+對該菊粉酶活性起促進作用,而Fe2+、Ba2+對該菊粉酶活性影響很小。
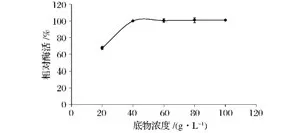
圖8 底物(菊粉)濃度對菊粉酶活力的影響Fig.8 Effect of inulin concentration on inulinase stability
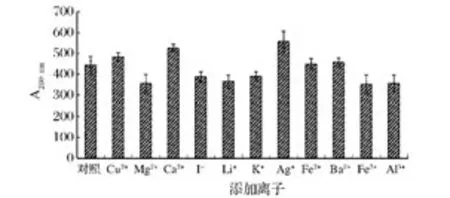
圖9 金屬離子對菊粉酶活力的影響Fig.9 Effect of various metallic ions on inulinase activity
2.2.5 反應時間對菊粉酶活力的影響
由圖10可見,菊粉酶在0~30 min反應時間與生成物濃度呈線性關系,此反應速度是恒定的。因此,前30 min內所測的反應速度可以認為是酶的初反應速度。所以,確定測定該酶活性的最適反應時間為30 min。
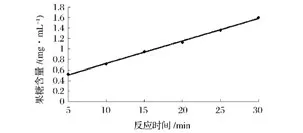
圖10 反應時間對菊粉酶活力的影響Fig.10 Effect ofreaction time on inulinase activity
2.2.6 菊粉酶的酶促反應動力學
酶反應動力學是研究酶反應速度規(guī)律以及各種因素對酶反應速度影響的科學。研究酶反應動力學規(guī)律對于基礎研究及生產應用都有十分重要的意義。酶反應動力學關系式,一般用米氏方程來表示。米氏方程中的Km是一個特征常數,它部分地描述了酶反應性質、反應條件對酶反應速度的影響。Km值越大,說明酶與底物的親和力越小;反之,可認為酶與底物的親和力越大[28]。根據圖11,由擬合直線的斜率和縱截距之比可求得米氏常數(Km),菊粉酶催化底物菊粉的酶促反應的米氏常數Km=3.83 mg/mL,最大反應速度(vmax)為4.64 mg/(mL·min)。
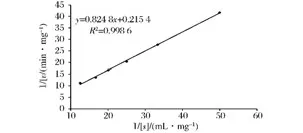
圖11 菊粉酶的Lineweave-Burk雙倒數曲線Fig.11 Lineweave-Burk double reciprocal curve of inulinase
3 結論
灰平鏈霉菌S.griseoplanus S501固態(tài)發(fā)酵液經采用硫酸銨分級沉淀、Sephadex G-7和DEAE-Sepharose CL-6B柱分離,獲得單組分菊粉酶,純化倍數為10.42倍,酶比活性249.5 U/mg蛋白。采用 SDSPAGE測定該菊粉酶組分的相對分子質量為23.442 kDa。酶學性質研究表明,該菊粉酶最適反應溫度和pH值分別為50℃和5.0。金屬離子Mg2+、I-、Li+、Fe3+、Al3+、K+對該菊粉酶有很強的抑制作用,Cu2+、Ca2+、Ag+對該菊粉酶活性起促進作用。以菊粉為底物時,該菊粉酶的Km為3.83 mg/mL,vmax為4.64 mg/(mL·min),表現為與菊粉底物具有較強的親和力。
參考文獻
[1] Roberfroid M B.Inulin-type fructans:functional food ingredients[J].The Journal of Nutrition,2007,137(11):2 493-2 502.
[2] 倪曉穎,李秉超,林榮峰,等.柱層析法分離純化菊粉酶的研究[J].沈陽農業(yè)大學學報,2010,41(4):484-487.
[3] 閆位娟,陳艷珍.菊粉酶分離純化方法的研究[J].食品與發(fā)酵工業(yè),2006,32(5):42-44.
[4] 周幗萍,沙濤.菊粉酶的研究與應用[J].食品與發(fā)酵工業(yè),2001,27(7):54-58.
[5] 王艷,金征宇.酵母菌C10產內切菊粉酶發(fā)酵條件的研究[J].生物技術,2001,11(6):25-27.
[6] 王艷,金征宇,徐學明,等.內切菊粉酶產生菌株的篩選及誘變選育[J].生物技術,2001,11(1):26-28.
[7] 陳曉明,朱太海.菊糖的功能及其在食品工業(yè)的應用[J].食品科技,2000(5):5-9.
[8] 張連富,李紅.內切菊粉酶法生產低聚果糖研究進展[J].中國食品添加劑,2000(1):20-25.
[9] 大連輕工學院,華南理工大學,鄭州輕工學院,等.食品分析[M].北京:中國輕工業(yè)出版社,2002:173.
[10] Gill P K,Sharma A D,Harchand R K,et al.Effect of media supplements and culture conditions on inulinase production by an actinomycete strain [J].Bioresource Technology,2003,87(3):359-362.
[11] Cazetta M L,Martin P M,Montir,et al.Yaeon(Polymnia sanehifolia)extract as a substrate to produce inulinase by KIuyveromyces marxianus var.bulgaricus[J].Journal of Food Engineering,2005,66(3):301-305.
[12] 陳冠軍,孫忠東.黑曲霉菊糖酶的純化及性質[J].微生物學報,1997,37(5):362-367.
[13] Uzunova K,Vassileva A,Kambourova M,et al.Production and properties of a bacterial thermostable exo-inulinase[J].Z Naturforsch,2001,56c:1 022-1 028.
[14] 賈英民,趙學慧.黑曲霉M89菊粉酶的提純與性質[J].微生物學報,1998,38(2):120-125.
[15] Moriyama S,Akitmoto H,Suctsugu N,et al.Purification and properties of an exracellular exoinulinase from Penicillium sp.strain TN-88 and sequence analysis of the encoding gene[J].Bioscience,Biotechnology,and Bioehemistry,2002,66(9):1 887-1 896.
[16] Uhm T B,Chung M S,Lee S H,et al.Purification and characterization of Aspergillus ficuum endoinulinase [J].Bioscience,Biotechnology,and Bioehemistry,1999,63(1):146-151.
[17] Ferreira M S S,De Andrade A V M,Kennedy J F.Properties of a thermostable nonspecific fructofuranosidase produced by cladosporium cladosporioides cells for hydrolysis of Jerusalem artichoke extract[J].Applied Biochemistry and Biotechnology,1991,31(1):1-9.
[18] Hamdy,Hossam S.Purification and some important characters of extracellular inulinase of Alternaria alternate(Fr)Keissle[J].Indian Journal of Experimental Biology,2002,40(12):1 393-1 398.
[19] Elena G D,Rui C,Bruno S F,et al.Towards the development of a membrane reactor for enzymatic inulin hydrolysis[J].Journal of Membrane Science,2006,273(1-2):152-158.
[20] Xiao R,Tanida M,Takao S.Purification and some properties of endoinulinase from Chrysosporium pannorum[J].Journal of Fermentation and Bioengineering,1989,67(4):244-248.
[21] 張國青,崔福綿,楊秀清.毛殼霉內切菊粉酶的純化與性質[J].微生物學報,2004,44(6):785-788.
[22] Nakamura T,Shitara A,Matsuda S,et al.Production,purification and properties of an endoinulinase of Penicillium sp.TN-88 that liberates inulotriose[J].Journal of Fermentation and Bioengineering,1997,84(4):31-318.
[23] Cho Y J,Yun J W.Purification and characterization of an endoinulinase from Xanthomonas oryzae No.5[J].Process Biochemistry,2002,37(11):1 325-1 331.
[24] Kang S I,Chang Y J,Oh S J,et al.Purification and properties of an endo-inulinase from an Arthrobacter sp.[J].Biotechnology Letters,1998,20(10):983-986.
[25] 張菊芬.宇佐美曲霉產菊粉酶及其酶學特性研究[D].鎮(zhèn)江:江蘇大學,2007.
[26] 徐艷新.菊粉酶產生菌的選育及其酶學性質的初步研究[D].無錫:江南大學,2013.
[27] Singh RS,Sooch B S,Puri M.Optimization of medium and process parameters for the production of inulinase from a newly isolated Kluyveromyces marxianus YS-1[J].Bioresource Technology,2006,98(13):2 518-2525.
[28] 肖厚榮.煙草多酚氧化酶同工酶的分離純化、表征、酶學性質和應用研究[D].合肥:中國科學技術大學,1999.
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
家庭影院技術(2020年10期)2020-12-14 07:54:18
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
知識經濟·中國直銷(2016年3期)2016-02-27 16:15:49
現代檢驗醫(yī)學雜志(2014年6期)2014-02-02 03:02:04
閱讀與作文(小學低年級版)(2011年3期)2011-01-01 00:00:00
