RNAi介導大鼠腺苷A1和A2A受體基因沉默對乙醛誘導的肝星狀細胞活化增殖的影響
2016-01-11 11:14:20王琪,王和,饒玲等
中國藥理學通報 2015年1期
關鍵詞:細胞增殖
網絡出版時間:2014-12-4 13:45網絡出版地址:http://www.cnki.net/kcms/doi/10.3969/j.issn.1001-1978.2015.01.012.html
RNAi介導大鼠腺苷A1和A2A受體基因沉默對乙醛誘導的肝星狀細胞活化增殖的影響
王琪,王和,饒玲,趙晗,楊鳳,楊巖,呂雄文,李俊
(安徽醫科大學藥學院,安徽醫科大學肝病研究所,安徽 合肥230032)
中國圖書分類號:R-332;R322.47;R329.24;R392.11;R575.2
摘要:目的研究大鼠腺苷A1受體(A1R)和腺苷A2A(A2AR)受體的siRNA分別轉染大鼠肝星狀細胞(HSC)對乙醛誘導的HSC活化增殖的影響。 方法采用乙醛誘導HSC-T6建立離體的大鼠酒精性肝纖維化HSC模型,設計并合成A1R和A2AR小干擾RNA(small interfering RNA, siRNA)序列,通過脂質體Lipofectamine`(TM) 2000瞬時轉染至HSC-T6細胞內,熒光倒置顯微鏡觀察細胞的轉染效率,用四甲基偶氮唑鹽(MTT)法檢測HSC-T6細胞增殖變化;利用Real-Time qPCR及Western blot法分別檢測HSC-T6的A1R、A2AR、α-SMA、Collagen I mRNA及蛋白表達。 結果將A1R和A2AR siRNA轉染至HSC-T6細胞內,A1R和A2AR基因及蛋白的表達水平明顯降低;同時α-SMA、Collagen I mRNA及蛋白表達水平亦明顯降低;靶向封閉A1R或A2AR基因的表達可明顯抑制HSC-T6細胞的活化增殖。 結論靶向封閉A1R或A2AR基因的表達可明顯抑制HSC-T6細胞的活化增殖,A1R和A2AR可能是潛在的酒精性肝纖維化的治療靶點。
關鍵詞:腺苷A1受體;腺苷A2A受體;肝星狀細胞;細胞增殖;乙醛;酒精性肝纖維化
doi:10.3969/j.issn.1001-1978.2015.01.012
文章編號:
文獻標志碼:A1001-1978(2015)01-0050-06
收稿日期:2014-09-03,修回日期:2014-11-22
基金項目:國家自然科學基金資助項目(No 81270498);安徽省自然科學基金項目(No 11040606M194);安徽高校省級科學研究重點項目(No KJ2012A148);國家級大學生創新創業訓練計劃項目(No 201310366032)
作者簡介:王琪(1992-),女,碩士生,研究方向:抗炎免疫藥理學, E-mail:leodd1992@126.com;
通訊作者呂雄文(1970-),男,博士,教授,碩士生導師,研究方向:抗炎免疫藥理學,,Tel: 0551-65161115,E-mail:lxw31288@aliyun.com
Abstract:AimTo investigate the influence of down-regulating adenosine A1 receptor and adenosine A2A receptor gene expression on proliferation and activation of acetaldehyde-induced hepatic stellate cell-T6 cells through siRNA. MethodsAlcoholic liver fibrosis in vitro model was constructed by inducing HSC-T6 cells with acetaldehyde. siRNA targeting A1R and A2AR were designed and synthesized according to its mRNA. The siRNA was transfected into rat HSC-T6 cells by liposome Lipofectamine`(TM) 2000. HSC cell proliferation was measured by MTT. The mRNA levels of A1R, A2AR, α-SMA, Collagen I in the supernatant of the cell culture were measured by Quantitative Real-Time PCR. The protein levels of A1R, A2AR, α-SMA, Collagen I were measured by Western blot. ResultsA1R and A2AR siRNA effectively inhibited the cell proliferation, and they also significantly decreased the levels of A1R, A2AR, α-SMA, Collagen I, suggesting that A1R and A2AR might be potential target genes in the alcoholic liver fibrosis. ConclusionsSilencing A1R or A2AR by RNAi can significantly inhibit the HSC proliferation, A1R and A2AR may be potential therapeutic target genes for alcoholic liver fibrosis.
酒精性肝病是指長期大量飲用各種含乙醇的飲料所導致的肝臟損害性病變[1]。在酒精性肝纖維化發生過程中,乙醇可通過代謝毒性產物、氧化應激、細胞因子相互作用等多種因素激活肝星狀細胞(hepatic stellate cell,HSC)。乙醇在酒精脫氫酶的作用下轉化成乙醛,接著在乙醛脫氫酶作用下轉化成醋酸,最后分解為二氧化碳和水。根據文獻報道,乙醛在酒精性肝纖維化的形成中起到關鍵作用。HSC的活化增殖是肝纖維化的核心環節,肝臟受損時HSC活化增殖轉化為肌樣成纖維化細胞,α-SMA合成與分泌細胞外基質(extracellular matrix, ECM)增多,導致ECM合成與降解失衡[2-3]。腺苷受體(adenosine receptors,ARs)家族包括A1、A2A、A2B和A3受體,屬于G蛋白偶聯受體超家族,廣泛存在于HSC、枯否細胞(kupffer cell, KC)和肝細胞中[4]。在酒精性脂肪肝以及肝纖維化發展過程中,腺苷和ARs激活的作用受到眾多研究者的關注。其中A1受體(A1 receptor,A1R)和A2A受體(A2A receptor,A2AR)為高表達受體,也是4種ARs中最主要的2種亞型[5]。有研究表明,在肝纖維化的發病機制中,腺苷受體活化扮演著重要的角色,有望成為預防和治療肝纖維化的一個新穎的靶點[6]。但ARs在酒精性肝纖維化中的重要調節功能仍未完全明確。本研究在采用乙醛誘導HSC-T6細胞建立離體大鼠酒精性肝纖維化HSC模型的基礎上,運用RNA干擾技術,通過構建針對A1R和A2AR基因的小干擾RNA,特異性降低A1R和A2AR的表達,觀察靶向抑制A1R和A2AR基因的表達對HSC活化增殖及對α-SMA、Collagen I表達的影響,為酒精性肝纖維化的防治提供新的靶點。
1材料
1.1實驗細胞株大鼠肝星狀細胞株(HSC-T6),購自中國科學院上海細胞研究所。
1.2試劑和儀器新生牛血清(杭州四季青生物工程材料有限公司,中國),DMEM培養液、TRIzol Reagent RNA 提取試劑(Gibco公司,美國),脂質體轉染試劑LipofectamineTM2000 (Invitrogen公司,美國),乙醛(質量分數40%,天津大茂化學試劑廠,中國),胰蛋白酶、RIPA、PMSF(碧云天公司,中國),逆轉錄試劑盒(Fermentas 公司,美國),SYBR Green,ECL化學發光試劑盒(Thermo公司,美國),A1R、A2AR抗體(安博公司,中國),α-SMA、Collagen I(博奧森公司,中國),β-actin抗體(中山金橋公司,中國),HRP標記的辣根過氧化物酶二抗(中山金橋公司,中國),引物(捷瑞公司,中國),siRNA序列(上海吉瑪公司,中國)。Thermo forma 3111型培養箱(Thermo公司,美國),倒置熒光顯微鏡(OLYMPUS公司,日本),酶標儀(BiotekEL,美國)
2方法
2.1HSC-T6細胞的培養大鼠肝星狀細胞系(HSC-T6)以10%小牛血清DMEM培養基培養,另外加抗生素(1×105U·L-1青霉素、100 mg·L-1鏈霉素)。置于CO2培養箱(5%CO2、37℃)中,1~2 d換液,2~3 d細胞傳代,換液用PBS沖洗3次,傳代時用0.25%的胰酶消化細胞1~2 min。
2.2siRNA的設計與合成A1R和A2AR的siRNA序列由上海吉瑪公司設計并合成,該序列經BLAST查詢,確認所設計siRNA序列的唯一性。另合成一對帶FAM標記的陰性對照siRNASRC序列。(A1R siRNA上游引物序列:5′-CCA GCA UUC UGA UCU ACA UTT -3′,下游引物為:5′-AUG UAG AUC AGA AUG CUG GTT -3′。 A2AR siRNA 上游引物序列:5′-GUG GCU GUA UGA AGU UGA ATT-3′,下游引物為:5′-UUC AAC UUC AUA CAG CCA CTT-3′。siRNASCR 上游引物序列:5′-UUC UCC GAA CGU GUC ACG UTT-3′,下游引物為:5′-ACG UGA CAC GUU CGG AGA ATT-3′)。
2.3A1R及A2AR siRNA轉染 取對數生長期HSC-T6細胞,用含10%新生牛血清不含抗生素的DMEM接種于培養瓶中,調整細胞濃度至每毫升4×105個。參照脂質體LipofectamineTM2000說明書進行siRNA轉染。首先轉染FAM標記陰性對照siRNA并在熒光顯微鏡下觀察轉染效率。實驗分為正常組、模型組、轉染組、陰性對照組。正常組不經任何處理,模型組、轉染組、陰性對照組加入終濃度為200 μmol·L-1的乙醛,并且每12 h補充1次。轉染組轉染A1R或A2AR siRNA,A1R及A2AR siRNA終濃度均為200 nmol·L-1。陰性對照組轉染FAM標記的陰性對照siRNA。按照上海吉瑪制藥技術有限公司siRNA說明書進行操作,用Opti-MEM(Invitrogen) 脂質體轉染試劑進行轉染,于37℃、5%CO2細胞培養箱中轉染6 h后每組將轉染培養基換為含10%血清培養基,繼續培養不同時間后收集。采用熒光倒置顯微鏡觀察各組轉染后細胞形態及其形態學變化,判斷轉染效果。
2.4細胞增殖實驗收集對數生長期細胞,胰酶消化后進行細胞計數,調整細胞濃度至每毫升1×105個接種于96孔板,每孔終體積100 μL,分為4組:正常組(常規培養);模型組(200 μmol·L-1乙醛刺激);siRNA組(200 μmol·L-1乙醛+siRNA 200 nmol·L-1);陰性對照組(200 μmol·L-1乙醛+siRNASCR),進行細胞轉染,6 h后棄去轉染培養基,每孔加入200 μL含10%血清培養基置37℃、5%CO2培養箱中培養48 h后加MTT(5 g·L-1)20 μL/孔,繼續培養4 h,棄上清,加DMSO 100 μL/孔,避光振蕩10 min,使結晶充分溶解后,待結晶充分溶解后,使用酶聯儀在492 nm波長處測各孔的吸光度。
2.5Real-time qPCR法檢測A1R、A2AR、α-SMA、Collagen I mRNA表達收集培養48 h的細胞,采用TRIzol法提取HSC總RNA,采用分光光度法測定其含量及純度(A260/A280>1.8)。取2 μg總RNA,參照Fermentas公司的RNA PCR Kit試劑盒說明書進行反轉錄合成cDNA。10 μL PCR反應體系,用PIKO REAL 96 進行實時定量PCR擴增,以β-actin作為內參。下列引物(Tab 1)均由上海捷瑞生物技術服務有限公司合成。

Tab 1 Sequences of primers used for real-time
2.6Western blot 檢測A1R、A2AR、α-SMA、Collagen Ⅰ蛋白表達收集培養48 h的細胞,每組加1 mL細胞裂解液(含10 μL PMSF),于冰上裂解30 min后置于4℃離心機上12 000×g,離心30 min。取上清,用BCA試劑盒進行蛋白定量。加入上樣緩沖液,100℃,加熱10 min使蛋白變性。每孔上樣20 μL總蛋白,進行SDS-聚丙烯酰胺凝膠電泳(濃縮膠電壓80V,分離膠120V)。電泳結束后用BioRAD濕轉儀器將蛋白轉移到PVDF膜上。膜在室溫下用含5%脫脂奶粉的TBST緩沖液封閉3 h后,用TBST清洗3次,每次15 min。加入第一抗體(1 ∶500),4 ℃中孵育過夜,再與HRP偶聯第二抗體(1 ∶10 000)室溫孵育1 h,ECL發光試劑盒顯影,以β-actin為內參,分別以相應蛋白與β-actin光密度比值表示該蛋白相對表達水平。以上實驗結果至少重復3次。

3結果
3.1熒光顯微鏡下觀察細胞轉染效率FAM標記siRNA轉染HSC-T6細胞48 h后熒光顯微鏡下觀察,可見轉染成功的HSC-T6細胞內有綠色熒光,而未轉染成功的細胞則無熒光,在同一位置經普通倒置顯微鏡和熒光顯微鏡對比觀察,轉染48 h后轉染效率可達70%以上,見Fig 1。

Fig 1 Transfection efficiency of siRNA on HSC-T6 cells
A, B: A1R; C, D: A2AR.
3.2siRNA對HSC-T6細胞增殖的作用 轉染siRNA 48 h后,細胞增長明顯減慢,MTT比色法結果顯示:與正常組比較,模型組和陰性對照組顯示,200 μmol·L-1的乙醛對HSC-T6細胞有明顯的增殖作用(P<0.05),siRNA組抑制率與模型組比較差異具有統計學意義(P<0.05)。表明A1和A2A受體siRNA對乙醛誘導的HSC細胞的增殖具有明顯抑制作用,見Fig 2。

Fig 2 Proliferation inhibitory effect of siRNA

#P<0.05vsnormal group;*P<0.05vsmodel group.
3.3Real-time qPCR檢測siRNA轉染后HSC-T6細胞A1R、A2AR、α-SMA、Collagen I mRNA表達變化采用相對定量法測定轉染siRNA后的HSC-T6細胞內A1R、A2AR基因相對于β-actin內參基因的表達變化。與正常組比較,乙醛作用48 h后,A1R、A2AR、α-SMA、Collagen I mRNA的表達明顯增強(P<0.01)。瞬時轉染A1R、A2AR siRNA 48h后,A1R、A2AR mRNA表達降低(P<0.05),與模型組相比差異有顯著性(P<0.05);同時,siRNA組α-SMA、Collagen I mRNA表達也降低(P<0.05),與模型組相比差異有顯著性(P<0.05) 模型組與陰性對照組之間的差異無顯著性(P>0.05),見Fig 3、4。

Fig 3 mRNA expression levels of A1R, α-SMA,

#P<0.05,##P<0.01vsnormal group;*P<0.05vsmodel group.

Fig 4 mRNA expression levels of A2AR, α-SMA, Collagen I

##P<0.01vsnormal group;*P<0.05vsmodel group.
3.4Western blot檢測siRNA轉染后HSC-T6細胞A1R、A2AR、α-SMA、Collagen I 蛋白表達變化Western blot檢測結果表明,200 μmol·L-1的乙醛對HSC-T6細胞增殖具有促進作用,模型組的A1R、A2AR、α-SMA、Collagen I蛋白表達明顯增強,與正常組比較差異有顯著性(P<0.05)。瞬時轉染A1R、A2AR siRNA 48 h后,A1R、A2AR蛋白表達降低(P<0.05),與模型組相比差異有顯著性(P<0.05),同時,siRNA組α-SMA、Collagen Ⅰ蛋白表達也降低(P<0.05),與模型組相比差異有顯著性(P<0.05)。陰性對照組與空白對照組之間的差異無顯著性(P>0.05),見Fig 5、6。

Fig 5 Protein expression levels of A1R, α-SMA,

#P<0.05,##P<0.01vsnormal group;*P<0.05vsmodel group.
4討論
HSC是位于大鼠肝Disses間隙內,緊貼肝竇內皮細胞與肝細胞,是肝內的貯脂細胞,正常情況下HSC表現為富含脂滴的靜止狀態,但當肝臟受到炎癥或機械刺激損傷時,HSC可被活化,HSC的激活是肝纖維化發生發展的中心環節[7]。酒精在生物體內經乙醇脫氫酶可代謝為乙醛,再經乙醛脫氫酶代謝為乙酸。乙醛作為乙醇的直接代謝產物,是激活HSC并導致酒精性肝纖維化發生的關鍵分子[8]。根據本課題組前期研究結果,選用200 μmol·L-1的乙醛作用48 h作為體外大鼠酒精性肝纖維化的細胞模型是較為理想的[9-11]。
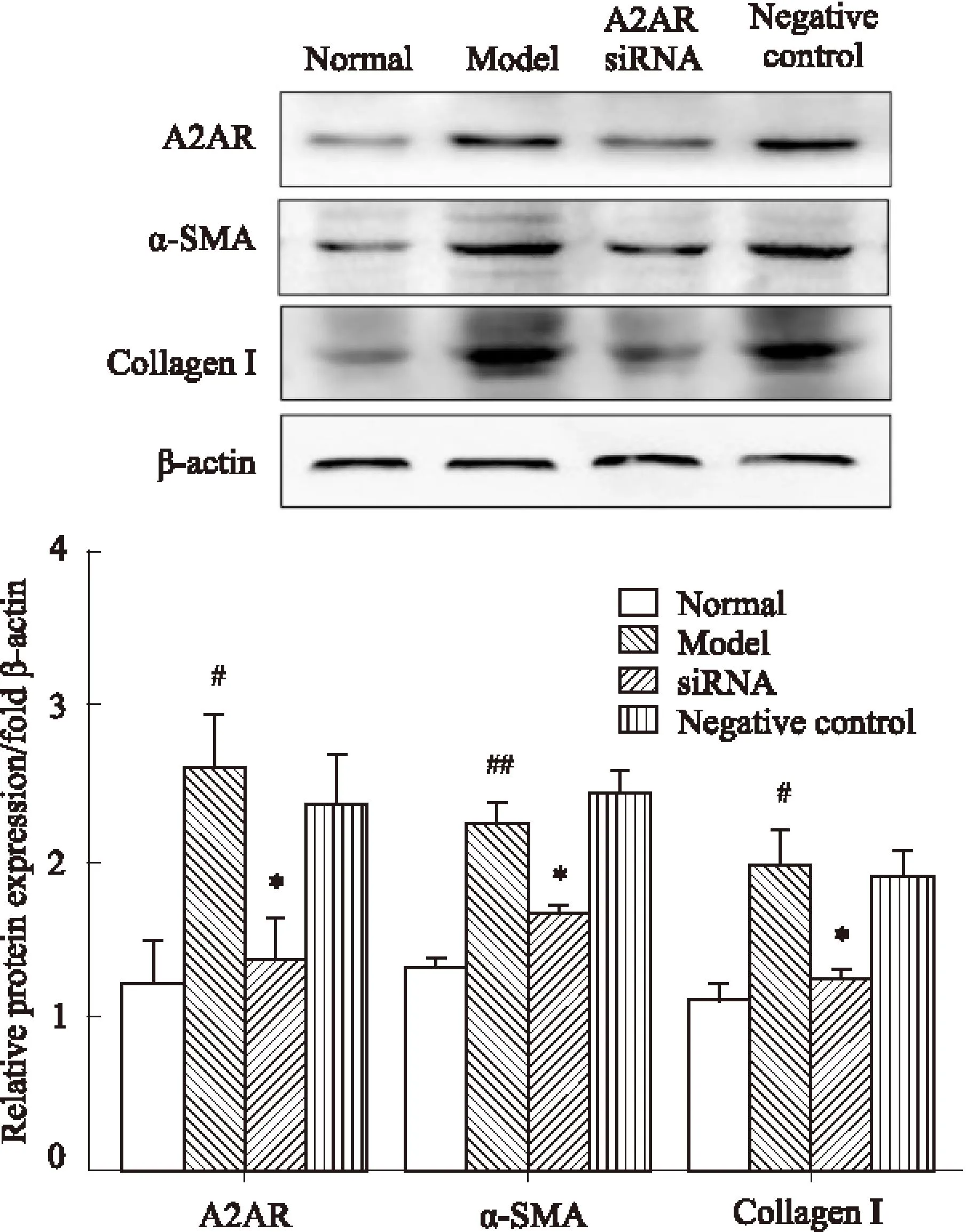
Fig 6 Protein expression levels of A2AR, α-SMA,

#P<0.05,##P<0.01vsnormal group;*P<0.05vsmodel group.
腺苷作為一種內源性嘌呤核苷,主要通過結合并激活與 G 蛋白偶聯的ARs,對機體的許多系統及組織發揮著重要的作用。目前已發現,ARs亞型包括 A1、A2A、A2B和A3受體,其中 A1受體和 A2A受體為高表達受體,也是 4種 ARs中最主要的2種亞型。 Peng等[12]2009年研究表明缺乏或阻斷A1R 可阻止小鼠酒精性脂肪肝的發生;Yang等[13]2010年研究報道長期注射 CCl4誘發的肝纖維化在 A1R 缺乏的小鼠體內減輕,且伴有膠原沉積和 HSC 活化明顯減少。亦有研究表明經乙醇誘導后的HSC通過激活A2AR,釋放大量腺苷,促進大鼠和人HSC膠原的合成,在Hashmi等[14]和Sohail等[15]的研究也證實了腺苷通過激活A2AR在HSC活化致肝纖維化中的作用。Che等[16]2007年也報道, A2AR通過其偶聯的Gs蛋白上調 cAMP-PKA 信號途徑,再分別經由其下游信號通路PKA-Src-ERK1/2 MAPK和p38 MAPK誘導LX-2人肝星形狀細胞株Ⅰ型膠原和Ⅲ型膠原的生成。
鑒于ARs的重要生理功能,本實驗采用特異性A1R和A2AR siRNA轉染HSC-T6細胞,并從多角度觀察轉染效果,成功建立A1R和A2AR siRNA轉染HSC-T6細胞的方法,為后續研究打下基礎。
觀察A1R和A2AR siRNA轉染后其mRNA和蛋白的表達,結果顯示不論是A1R或A2AR的mRNA及蛋白表達水平均明顯減少,說明轉染成功。200 μmol·L-1乙醛作用48 h可激活HSC-T6細胞,使α-SMA、Collagen Ⅰ mRNA和蛋白的表達明顯增高,但A1R或A2AR siRNA轉染對乙醛誘導的HSC-T6細胞的增殖具有抑制作用,轉染A1R或A2AR siRNA 48 h后,細胞增長明顯減慢,且轉染后可明顯抑制α-SMA、Collagen Ⅰ的mRNA和蛋白的表達,再次證明轉染成功,且表明沉默A1R或A2AR可使HSC的活化增殖受到抑制。
綜上所述,本研究通過利用腺苷A1R或A2AR siRNA下調A1R及A2AR基因的表達,可明顯抑制乙醛誘導的HSC-T6細胞的活化,抑制α-SMA,Collagen Ⅰ的產生。提示A1R和A2AR在肝纖維化的形成過程中發揮了重要作用,有望成為防治酒精性肝纖維化的靶點之一。但肝纖維化形成是一個復雜的多因素過程,ARs如何調控相關信號通路抑制HSC的活化和增殖,尚需進一步深入研究。
參考文獻:
[1]Reyes-Gordillo K, Shah R, Arellanes-Robledo J, et al. Mechanisms of action of Acetaldehyde in the up-regulation of the human α2 (I) collagen gene in hepatic stellate cells: Key roles of Ski, SMAD3, SMAD4, and SMAD7[J].AmJPathol, 2014, 184(5): 1458-67.
[2]Hernandez-Gea V, Friedman S L. Pathogenesis of liver fibrosis. Annual review of pathology [J].MechanDis, 2011, 6:425-56.
[3]陶輝, 黃成, 楊晶晶, 等. RNAi介導MeCP2基因沉默對HSC-T6細胞活化增殖的影響[J]. 中國藥理學通報, 2012, 28(3): 333-6.
[3]Tao H, Huang C, Yang J J, et al. Experimental study of proliferation and activation of HSC-T6 cells through RNA inference targeting MeCP2 [J].ChinPharmacolBull, 2012, 28(3):333-6.
[4]Sands W A, Palmer T M.Adenosine receptors and the control of endothelial cell function in inflammatory disease [J].ImmunolLett, 2005, 101(1):1-11.
[5]Gessi S, Merighi S, Varani K, et al. Adenosine receptors in health and disease [J].AdvPharmacol, 2011, 61:41-75.
[6]Chan E S, Cronstein B N. Adenosine in fibrosis [J].ModRheumatol, 2010, 20(2): 114-22.
[7]Cohen J, Nagy L E. Pathogenesis of alcoholic liver disease: interactions between parenchymal and non-parenchymal cells [J].JDigDis, 2011, 12(1):3-9.
[8]邱萍, 李相, 孔德松, 等. 酒精性肝病發病機制研究的新進展[J]. 中國藥理學通報, 2014, 30(2): 160-3.
[8]Qiu P, Li X, Kong D S, et al. Research progress on pathogenesis of alcoholic liver disease [J].ChinPharmacolBull, 2014, 30(2): 160-3.
[9]楊萬枝,呂雄文,余世春,等.cAMP-PKA-CREB信號通路在大鼠酒精性肝纖維 化星狀細胞模型中的作用[J].安徽醫藥, 2012, 16(6):729-31.
[9]Yang W Z, Lv X W, Yu S C, et al. Effect of cAMP-PKA-CREB signal pathway in the model of alcoholic hepatic fibrosis stellate cells isolated from rats [J].AnhuiMedPharmJ, 2012, 16(6):729-31.
[10]管文婕,呂雄文,楊萬枝,等.咖啡因對乙醛誘導的HSC-T6中TGF-β1,CTGF信號轉導通路的影響[J].安徽醫藥, 2012, 16(8):1060-3.
[10]Guan W J, Lv X W, Yang W Z, et al. Effect of caffeine on signaling transduction of TGF-1 and CTGF in hepatic stellate cell-T6 stimulated by acetaldehyde [J].AnhuiMedPharmJ, 2012, 16(8):1060-3.
[11]Wang H, Guan W, Yang W, et al. Caffeine inhibits the activation of hepatic stellate cells induced by acetaldehyde via adenosine A2A receptor mediated by the cAMP/PKA/SRC/ERK1/2/p38 MAPK signal pathway[J].PloSone, 2014, 9(3): e92482.
[12]Peng Z, Borea P A, Varani K, et al. Adenosine signaling contributes to ethanol-induced fatty liver in mice [J].JClinInvest, 2009, 119(3):582-94.
[13]Yang P, Han Z, Chen P, et al. A contradictory role of A1 adenosine receptor in carbon tetrachloride and bile duct ligation induced liver fibrosis in mice[J].JPharmacolExpTher, 2010, 332(3):747-54.
[14]Hashmi A Z, Hakim W, Kruglov E A, et al. Adenosine inhibits cytosolic calcium signals and chemotaxis in hepatic stellate cells [J].AmJPhysiolGastrointestLiverPhysiol, 2007, 292(1):395-401.
[15]Sohail M A, Hashmi A Z, Hakim W, et al. Adenosine induces loss of actin stress fibers and inhibits contraction in hepatic stellate cells via Rho inhibition [J].Hepatology, 2009, 49(1):185-94.
[16]Che J, Chan E S, Cronstein B N. Adenosine A2A receptor occupancy stimulates collagen expression by hepatic stellate cells via pathways involving protein kinase A, Src, and extracellular signal-regulated kinases 1/2 signaling cascade or p38 mitogen-activated protein kinase signaling pathway [J].MolPharmacol, 2007, 72: 1626-36.
Proliferation and activation of acetaldehyde-induced HSC-T6
cells through RNA inference targeting adenosine A1 and A2A receptors
WANG Qi, WANG He, RAO Ling, ZHAO Han, YANG Feng, YANG Yan, Lü Xiong-wen, LI Jun
(SchoolofPharmacy,InstituteofLiverDiseases,AnhuiMedicalUniversity,Hefei230032,China)
Key words: adenosine A1 receptor; adenosine A2A receptor;hepatic stellate cell;cell proliferation;acetaldehyde;alcoholic liver fibrosis
猜你喜歡
中國當代醫藥(2017年2期)2017-03-18 18:17:33
右江醫學(2016年4期)2017-01-05 16:26:48
中國民族民間醫藥·下半月(2016年6期)2016-11-02 20:27:22
中國醫藥科學(2016年9期)2016-07-25 23:04:03
湖南師范大學學報·自然科學版(2016年3期)2016-06-25 06:18:59
學周刊·下旬刊(2016年7期)2016-05-20 18:48:49
課程教育研究·學法教法研究(2016年6期)2016-04-26 10:06:57
新課程·中學(2015年9期)2015-10-26 19:00:20