柏木低效林林窗改造初期邊界木細根形態和生物量變異
2016-01-12 08:37:40蘇宇,李賢偉,劉運科等
西北植物學報 2015年3期
關鍵詞:形態
*通信作者:李賢偉,教授,主要研究從事森林培育研究。E-mail:lxw@sicau.edu.cn
柏木低效林林窗改造初期邊界木細根形態和生物量變異
蘇宇1,李賢偉1,2*,劉運科1,李金鳳1,尹海峰1,王之南1
(1 四川農業大學 林學院,四川雅安 625014;2 長江上游林業生態工程四川省重點實驗室,四川雅安 625014)
摘要:為深入探究川中丘陵區柏木低效林林窗改造初期邊界木林細根的生態適應,利用土柱法對34年生柏木低效林在人造小面積開窗(50 m2、100 m2、150 m2)處理1年后的林窗邊界木1~5級細根直徑、根長、比根長、根長密度和生物量變異進行研究。結果顯示:(1)柏木1~5級細根平均直徑、根長隨著根序的上升而增加,比根長、根長密度隨著根序的上升而減小。 (2)與林內對照(無林窗)相比,人工開林窗處理后邊界木細根在同一根序上的平均直徑變化不顯著,根長和比根長增加,根長密度和生物量則呈降低的趨勢。(3)開窗 150 m2面積林窗撫育后的柏木邊界木更大限度地提高了低級根序的物質分配,提高比根長并降低組織密度,更快速地利用地下林窗空間資源。研究表明,柏木低效林林窗式疏伐得到的土壤資源由于更新草本、灌木的生長,增加了林木根系競爭土壤空間的壓力,使得根系進行擴展性生長,細根形態和生物量表現出異質性變化。
關鍵詞:低效林;林窗;邊界木;根序;形態;生物量
收稿日期:2014-10-07;修改稿收到日期:2015-01-09
基金項目:國家“十二五”科技支撐計劃項目(2011BAC09B05);四川省科技支撐計劃項目(2010NZ0049);國家自然科學基金面上項目(30771717)
作者簡介:蘇宇(1991-),女,在讀碩士研究生,主要從事森林生態研究。E-mail:yusu110@163.com
中圖分類號:Q948.12 文獻標志碼:A
Fine Root Morphology and Biomass Characteristics at
Preliminary Stage of Gap Border Trees of Reformed
Low BeneficialCupressusfunebrisForests
SU Yu1,LI Xianwei1,2*,LIU Yunke1,LI Jinfeng1,YIN Haifeng1,WANG Zhinan1
(1 College of Forestry,Sichuan Agricultural University,Ya’an,Sichuan 625014,China;2 Sichuan Province Key Laboratory of Forestry Ecological Engineering in Upper Reaches of Yangtze River,Ya’an,Sichuan 625014,China)
Abstract:In order to adapt the ecological adaptation of fine root of gap border trees at preliminary stage,we studied the 34-year old Cupressus funebris plantation in central Sichuan hilly region,researched the first five orders of fine rootdynamic rules of fine root morphology and biomass ofdifferent size of gaps (50 m2,100 m2 and 150 m2) and C.funebris plantation (Ⅳ) at the first year.The main results were found as follows:(1)The fine root morphology and biomass ofdifferent root orders of C.funebris plantation have obvious change rule.The specific root length(SRL) and root lengthdensity(RLD) of the first order root were the largest,but the averagediameter,average root length and biomass of the first order root were the smallest.The SRL and RLD of the fifth order root were the smallest,but the averagediameter and biomass of the fifth order root were the largest.Fine root biomass and averagediameter of C.funebris significantly increased with increase in order(P<0.05),while SRL and RLDdecreased.(2)Compared with C.funebris plantation (Ⅳ),fine root of gap border trees(GBT) have insignificant averagediameter,increase average root length and SRL,reduce RLD and biomass at the same root order.(3)GBT at the gap of 150 m2 highly raised Matterdistribution of low root orders,with increasing SRL anddecline RLD,to use underground forest gap in a more quality way.In conclusion,with the renew of herbs and shrubs the forest gaps have increased pressure of root to competition soil spaces,expansionary growth and heterogeneous exchange of fine root morphology and biomass.
Key words:low beneficial forests;forest gap;gap border trees;root order;morphology;biomass
森林是陸地生態系統的主體,維持著整個生態系統的物質循環和能量流動[1]。細根作為森林生態系統中的重要組成部分,是碳庫平衡和土壤養分循環進入陸地生態系統的基礎[2-3],細根在執行土壤資源有效分配的功能時,對其變化能做出快速的反應,并在生物量分布與形態特征上都表現出異質性[4-5],雖然細根(D<2 mm)生物量僅占根系總生物量的3%~30%,但在總碳循環和年初級生產力(NPP)上都占有較大的比重[6-7]。細根形態在根系功能的維持中作用重大,是衡量植物生產力的重要因素[8-10]。細根比根長(specific root length,單位重量的根長)和根長密度(root lengthdensity,單位面積的根長)決定根系吸收養分和水分的能力[11],但根系構型的改變可能并不引起根系總生物量的變化[12]。同樣,單一的生物量指標不能作為根系系統中吸收器官潛在功能的指示指標,采用綜合的根系形態參數可反映林地養分潛能并有效預測林分生產力[13]。
中國作為世界上人工林面積最大的國家,人工林地力衰退問題一直阻礙著林業可持續發展,地力衰退是一個復雜的生態學過程,而本質上就是林地林木生長養分吸收與土壤養分供給之間的不平衡。在人工林的撫育過程中,林窗干擾拓展了林隙空間、釋放了有效資源,使得生物多樣性增加、林下次生生長恢復,已有的研究多集中于林窗中光照、微地形、小氣候、生物多樣性和資源利用率等方面[14-15],而針對林窗邊緣邊界木的研究則缺少相應的關注[16-17],在小面積林窗條件下,大部分的林窗面積被邊界效應影響,邊界木對林窗環境因子的響應極為顯著[18],林冠林窗出現后伴隨著上層優勢苗木的死亡和倒伏,根系的生物量也隨之減少,形成不同于林內地下生物量的地下林窗微環境。地下林窗也同林冠林窗一樣是森林資源有效改善的重要部分,現有很少從林下土壤及根系生長的角度進行研究。
本研究以川中低山丘陵區柏木低效林為研究對象,采用小面積林窗采伐的人為干擾方式進行撫育管理,在控制水土流失、縮短植被恢復期的情況下,有效改善林分結構、促進林木生長、維持系統的生物多樣性。在人工開窗一年后針對不同面積林窗邊界木的地下根系進行研究,探討地下空窗對于邊界木細根生長與生物量的影響,充分認識林窗的基本特征,了解林窗邊界木地下競爭機制,加強對林木地上以及地下生長趨向性發育的探索,將有助于進一步揭示林窗對森林更新群落組成、結構和功能的動態影響及其維持機制。
1研究地區與研究方法
1.1 研究區概況
研究區位于德陽市旌陽區和新鎮(104°15′~104°33′E,31°5′~31°20′N),地處川中丘陵區北部邊緣,屬于典型的低山丘陵地貌。該區屬四川盆地中亞熱帶濕潤氣候區,氣候特征四季分明,氣候溫和,雨量充沛,日照較少,年平均氣溫16~17 ℃,年總降水量880~940 mm,年平均無霜期270~290d。該區域森林土壤主要為石灰型紫色土,土層淺薄,通常不超過40 cm。樣地區域土壤基本養分中全碳為39.11 g/kg,全氮為1.71 g/kg,全磷為0.27 g/kg,水解氮76.34 mg/kg,有效磷7.58 mg/kg,速效鉀105.34 mg/kg。
1.2 樣地設置與研究方法
該研究區域柏木人工林為34年生林分,區內林木尚處于中幼齡林階段,其平均樹高6.5 m,平均胸徑8.0 cm,郁閉度大于0.8,單位蓄積量約為44.7 m3/hm2,遠低于全國平均水平83.63 m3/hm2[19],屬于典型的低效林。針對實驗區林分的生境條件與生長情況,2012年3月,選擇在山體南坡向,中坡位,坡度20°~30°,生長狀況及經營管理水平相似的地段進行小面積林窗干擾采伐,面積分別為50 m2、100 m2和150 m2,每種采伐面積3個重復,各處理樣地的基本情況見表1。人造林窗均為南北向長、東西向短的近似橢圓形,且各林窗之間間隔15 m以上的過渡帶。清除枯枝和雜灌,建立鐵絲圍欄,防治人畜干擾,使其自然恢復。林窗面積的計算采用橢圓面積公式:A=πLW/4,式中,L為林窗中最長的直徑,W為與L垂直的最大直徑,其相交點作為林窗的中心點[20]。
2013年5月,在每個林窗周邊選取8株柏木邊界木,同時在未進行林窗改造的對照林分隨機選取8株樣木作為研究對象。在面向林窗方向距林窗邊界木樹基50 cm內用土塊法采集細根樣品,土塊大小為15 cm×15 cm×15 cm。土塊放入低溫保鮮盒中帶回實驗室。在實驗室,把未分離土塊放入盛有低溫(2~4 ℃)去離子水中,小心的將根系取出。在10倍放大鏡下,將根系上剩余的土、有機物質和已死的細根片段洗凈。土塊得到的細根樣使用用Fitter的根系序列位置命名方法對細根進行分級[21],位于根軸最遠端沒有分枝的根稱為1級根,1級根著生在2級根上,依次分到5級根,著生于高級根上沒有分枝的根也算作1級根。
應用Epson數字化掃描儀(Expression 10000XL 1.0)和Win RHIZO 2012根系圖像分析系統軟件對不同處理的1~5級柏木細根形態特征指標(直徑、根長、表面積等)進行定量分析。然后在65 ℃烘干至恒重,用電子天平稱重(精確到0.000 1 g)測定各級細根質量。得到各處理細根比根長(SRL),根長密度(RLD)以及生物量數據,計算公式為:
SRL=L/M;RLD=L/V;B=M/d2×104
式中,SRL為比根長(m·g-1);L為根系長度(m);M為根系干質量(g);RLD為根長密度(m·m-3);V為土塊體積(m3);B為細根生物量(g·m-2);d為土塊邊長(cm)。
1.3 數據處理與統計分析
數據采用Excel 2010和SPSS 20.0軟件做統計處理和分析,采用單因素方差分析(one-way ANOVA)結合Duncan檢驗分析比較不同開窗面積處理下邊界木不同根序間細根直徑、根長、比根長、根長密度的差異(P<0.5)。
2結果與分析
2.1 不同林窗面積下邊界木各級根序形態特征
由圖1,A可以看出,各處理樣地柏木細根平均
直徑隨著根序的上升而增加,前5級細根的直徑在0.36~1.50 mm之間,其中1、2級根之間差異不顯著(P>0.05),2~5級根之間則呈現顯著性差異(P<0.05),且各面積開窗邊界木1~5級細根平均直徑大小均無顯著影響;此外,各處理樣地柏木1~5級細根平均直徑呈現不均勻增長狀態,并在3級根出現快速增長的拐點,且3~5級根的平均直徑分別是1級根的1.3倍、2.1倍和3.6倍。
各處理柏木細根根長方差分析顯示(圖1,B),不同面積開窗處理對邊界木各級根序根長的影響不同。50、100 m2開窗處理后,林窗邊界木細根平均根長在1~5級根之間均表現出顯著的差異(P<0.05);150 m2開窗后,林窗邊界木1~5級細根平均根長則同對照處理的柏木細根變化規律一致,其中,在1、2級,2、3級根序之間均未表現出顯著差異性,在3~5級根序間差異顯著(P<0.05);50、100 m2開窗處理后,邊界木2~4級根序細根平均根長大于150 m2林窗細根;此外,對照組的5級根根長顯著小于各面積林窗處理(P<0.05)。
不同開窗面積對柏木細根不同根序比根長的影響見圖1,C。各處理細根SRL均表現為1~5級根依次降低,然而,同一級根序內SRL隨著開窗處理面積的增加而增加,對照處理則最低;各處理的同級根序SRL規律表現為:對照<50 m2<100 m2<150 m2;此外,150 m2開窗處理后細根SRL在1~5級根均較對照處理有顯著增加(P<0.05),100 m2處理細根SRL僅在3~5級根顯著大于對照(P<0.05),而50 m2處理細根SRL與對照比較并沒有顯著差異(P>0.05)。
由圖1,D可發現,各面積開窗處理后,邊界木1~3級細根RLD均顯著低于對照組(P<0.05),而4、5級根RLD僅在150 m2開窗處理后,其邊界木較對照組出現顯著降低(P<0.05)趨勢。總之,不同面積開窗處理對細根不同根序根長密度影響不一致,1、2級根RLD表現為:50 m2<100 m2<150 m2,而3~5級根RLD表現為:50 m2、100 m2處理均大于150 m2處理。
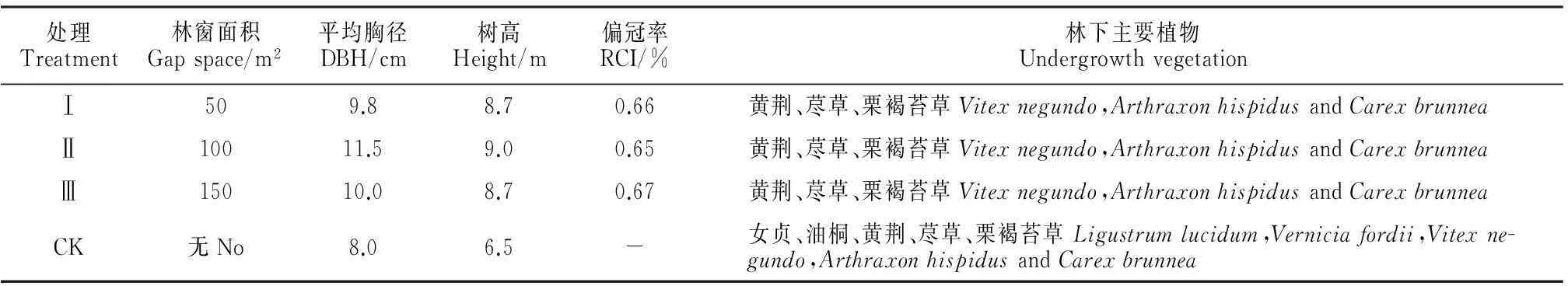
表1 柏木人工林人造林窗處理樣地基本概況
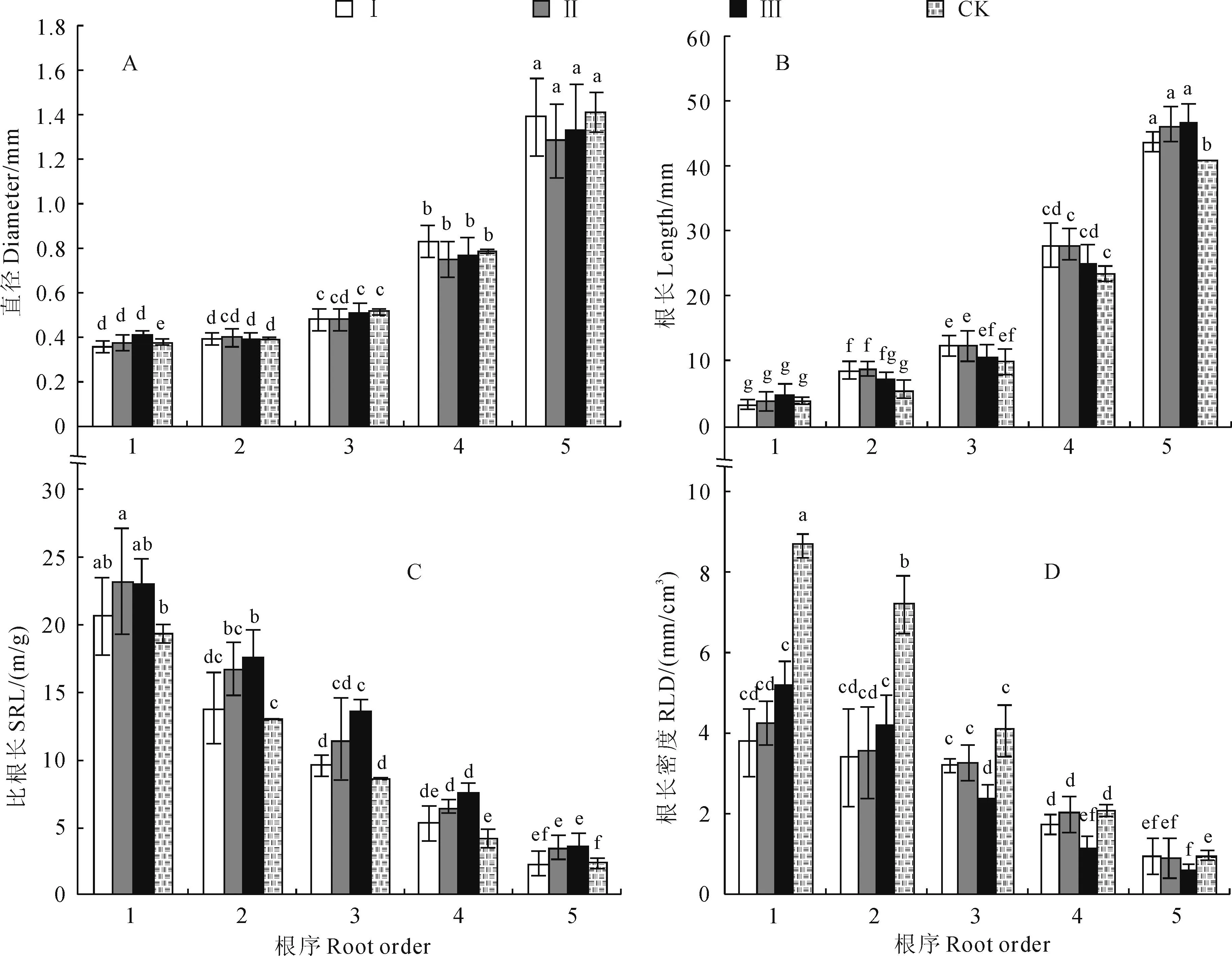
圖1 不同林窗面積下邊界木各級根序細根直徑、根長、比根長、根長密度
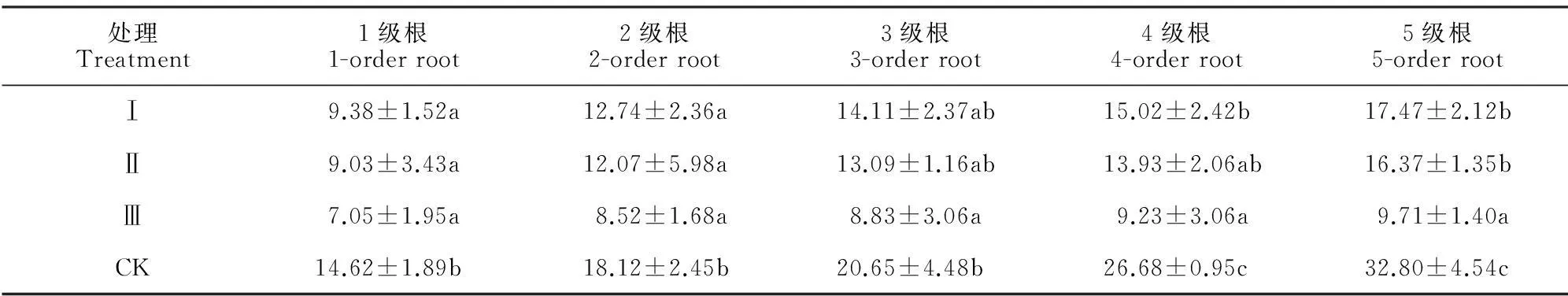
處理Treatment1級根1-orderroot2級根2-orderroot3級根3-orderroot4級根4-orderroot5級根5-orderrootⅠ9.38±1.52a12.74±2.36a14.11±2.37ab15.02±2.42b17.47±2.12bⅡ9.03±3.43a12.07±5.98a13.09±1.16ab13.93±2.06ab16.37±1.35bⅢ7.05±1.95a8.52±1.68a8.83±3.06a9.23±3.06a9.71±1.40aCK14.62±1.89b18.12±2.45b20.65±4.48b26.68±0.95c32.80±4.54c
注:不同小寫字母表示同列不同處理柏木細根生物量差異顯著(P<0.05)。
Note:Different normal letters within the same column meant significantdifference at 0.05 level.
2.2 不同林窗面積下邊界木各級根序生物量
不同林窗面積下邊界木1~5級根序生物量以及分配比例(表2;圖2)。各處理柏木細根生物量分配均表現為1級根最小,5級根最大;150 m2開窗處理邊界木各級根序生物量均顯著小于對照,50 m2、100 m2處理除3級根外生物量也顯著小于對照(P<0.05);此外,隨著開窗面積的增加,50 m2、100 m2、150 m2開窗處理的1~3級根生物量占前5級根總生物量的比值均有所增加,1級所占比例分別比對照組增加0.70 %、1.05 %、3.31%。
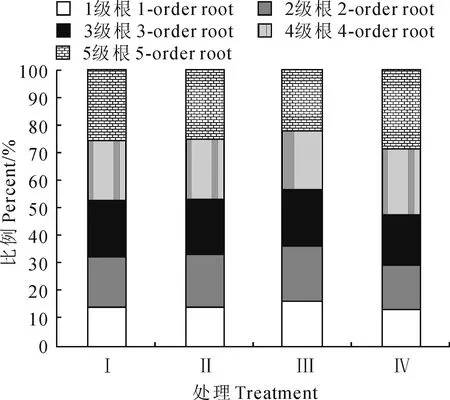
圖2 各級根序占總生物量的百分比
3結論與討論
3.1 不同林窗面積下邊界木細根形態特征
根系系統是植物從土壤中吸收水分與養分的主要器官,在地下生態系統的循環中有重要意義[22],細根直徑、根長、比根長和根長密度都是反映細根形態和功能的重要指標。本文以人工撫育開窗處理初期反應最快的林窗邊界木細根為研究對象,研究柏木細根特征和各形態指標對不同面積開窗處理的響應。本實驗對柏木1-5級細根形態的研究表明,1級根直徑、根長、比根長最小,根長密度最大,而5級根直徑、根長、比根長最大,根長密度最小,這種根序間的變化趨勢與差異性說明了細根存在普遍異質性[23-25],細根形態異質性與其生理功能相適應,這些特征同時又受到植物自身遺傳特性及立地條件的影響,并對土壤條件及環境變化的反應較為敏感[26-27],故而細根中行使不同生理功能的根序會做出不同的應對策略。
開窗后,邊界木細根在同一根序上的平均直徑變化不顯著,而根長和比根長則有所增加,根長密度呈現降低的趨勢。不同面積撫育開窗后的柏木邊界木以及對照樣地內柏木的細根直徑均呈現隨著根序增加直徑增加的規律,且各處理之間并沒有顯著的變化,這與“擴展的土壤資源并不能通過顯著增加柏木細根的直徑來拓展養分利用范圍”的結論相吻合[28]。本研究中柏木前4級根序的直徑都在1 mm以下,直徑較細,具有較強的吸收功能,有利于從土壤中吸收養分和水分[29],同時,開窗處理并沒有對周轉快速的1級細根根長產生影響,但隨著根序的增加細根對開窗響應的敏感度呈現上升趨勢,相較于150 m2林窗中較為疏松的邊界木分布情況,位于50 m2和100 m2林窗的邊界木根長出現了更顯著的增加,這使得根系增殖速度快,活力提高,土壤空間的利用效率增加[30-31]。
林窗邊界木各級根序細根比根長較對照都有增幅,這種變化隨開窗面積的增加而增加,細根中出現較大的比根長表明根系在該根序生物量的投入方面提高了效率;根長密度作為單位面積上根的長度,直觀反映樹木根系吸收水肥的能力,是樹木地下競爭力的體現[32],但是開窗處理減小了邊界木細根根長密度,1~3級根和4、5級根序表現出不同的響應機制,低級根序的細根根長密度降低程度尤為突出。此現象可能是由于開窗處理帶來的林冠林窗與地下林窗的雙向形成和光照增加導致的土壤水分養分變化和林下生物多樣性的改變,細根根長密度與土壤有效N呈現顯著相關[33],在開窗初期地上部分的伐除導致邊界木地下有效養分競爭的增加,邊界木細根生長的相對應策略變為增加單位生物量的根系長度,進而通過比根長的增加和根長密度的減少來快速利用土壤空間。
3.2 不同林窗面積下邊界木細根生物量
細根是林木從土壤中吸收養分和水分并維持自身正常生理活性的重要器官,其生物量與立地條件、土壤類型、群落結構、氣候、樹種等因素有關[34]。川中丘陵區森林分布范圍內土壤類型主要為新沖積物的灰棕和紫色沖積土,土層淺薄,低于40 cm,根據實地土壤剖面確定柏木根系中的細根分布在15 cm以內的淺表層土壤中(占總數生物量的85%以上),Makita等[35]的研究同樣指出表層土壤中根系的65%為細根。本研究對柏木細根生物量的研究就集中在15 cm以內,結果表明柏木細根生物量隨根序上升增加,然而前人的研究表明不同樹種的細根生物量隨著根序變化的規律并不一致,這說明細根生物量在各根序的分布會由于樹種和立地條件的不同而不同[24,36-37]。
開窗處理伴隨著邊界木細根生物量的減少,在50 m2開窗的邊界木細根上縮減39.12%,開窗150 m2后更是縮減61.60%,但由于在開窗的初期,撫育采伐強度的增大,使得單位面積上各級根序的生物量隨著開窗面積的增加而減少,且早期樹木生長的擴展性更多的表現在細根不同根序生物量的分配上。本研究發現,開窗后柏木低級根序所占總生物量的比值有不同程度的提高,特別是在150 m2開窗處理下,不同根序細根生物量所占比值的變化反應出根系功能分配的變化,造成此結果的原因可能是植物細根可塑性對潛在的養分、水分吸收和物種本身指定的覓食策略的響應使得低級根序在生物量總量中所占的比值增加,優先擴展活性強的器官,同時伴隨降低根系組織密度拓展利用地下林窗空間,高效平衡其生長所需的資源[38]。同時,低級根的高比根長可以保持細根的吸收能力,且由于低級根生物量較小,細根構建和維持單位長度細根消耗的碳水化合物較少,由這種比根長與生物量不對稱的比例關系可知,異質性的物質分配對細根的吸收功能有利[39]。
本研究使用的小面積林窗撫育間伐方式,快速干擾了已處于退化狀態的柏木純林,增加了林內光照,改變了土壤條件,影響了植物地上部分的生長,而這些因素均與細根的生長有密切的聯系,林下植被物種數和多樣性隨開窗面積的增加而增加,更新的林下灌木和草本都是淺根系植物,這勢必增加了喬木樹種根系競爭土壤資源的壓力[40],使得喬木樹種的根系需要更加快速的進行擴展性、養分誘導性生長[41]。在開窗撫育的初期,林窗邊界木作為首先受到林窗影響的生態邊界,其根系在形態和生物量分布上都做出了快速的競爭策略,在直徑基本不變的基礎上,通過增加根長,降低組織密度,迅速占領營養空間。研究根系分布在環境改變響應上的敏感性,有利于完整掌握在人為干擾下的森林生態系統養分庫養分循環和碳分配流動。林窗改造恢復過程是一個動態的過程,所形成的界面是非穩定的界面,針對整體林分變化的持續監測是人工林分撫育的必要手段,更是今后研究的方向。
參考文獻:
[1]FINER L,OHASHI M,NOGUCHI K,etal.Fine root production and turnover in forest ecosystems in relation to stand and environmental characteristics[J].ForestEcologyandManagement,2011,262(11):2 008-2 023.
[2]FAHEY T J,HUGHES J W.Fine rootdynamics in a northern hardwood forest ecosystem,Hubbard Brook Experimental Forest,NH[J].JournalofEcology,1994:533-548.
[3]KING J S,ALBAUGH T J,Allen H L,etal.Below-ground carbon input to soil is controlled by nutrient availability and fine rootdynamics in loblolly pine[J].NewPhytologist,2002,154(2):389-398.
[4]HISHI T.Heterogeneity of individual roots within the fine root architecture:Causal links between physiological and ecosystem functions[J].JournalofForestResearch,2007,12(2):126-133.
[5]MAJDI H,TRUUS L,JOHANSSON U,etal.Effects of slash retention and wood ash addition on fine root biomass and production and fungal mycelium in a Norway spruce stand in SW Sweden[J].ForestEcologyandManagement,2008,255(7):2 109-2 117.
[6]HELMISAARI HS,MAKKONEN K,KELLOMAKI S,etal.Below- and above-ground biomass,production and nitrogen use in Scots pine stands in eastern Finland[J].ForestEcologyandManagement,2002,165(1):317-326.
[7]TATENO R,HISHI T,TAKEDA H.Above- and belowground biomass and net primary production in a cool-temperatedeciduous forest in relation to topographical changes in soil nitrogen[J].ForestEcologyandManagement,2004,193:297-306.
[8]HARPER J L,JONES M,HAMILTON N R.The evolution of roots and the problems of analyzing their behavior[M].Oxford:Blackwell Scientific Publications,1991:3-24.
[9]HENDRICK RL,PREGITZER KS.Thedemography of fine roots in a northern hardwood forest[J].Ecology,1992,73:1 094-1 104.
[10]NADELHOFFER KJ,RAICH JW.Fine root production estimates and belowground carbon allocation in forest ecosystems[J].Ecology,1992,73:1 139-1 147.
[11]FRANSEN B,KROON HD,BERENDSE F.Root morphological plasticity and nutrient acquisition of perennial grass species from habitats ofdifferent nutrient availability[J].Oecologia,1998,115(3):351-358.
[12]HODGE A.The plastic plant:root responses to heterogeneous supplies of nutrients[J].TheNewPhytologist,2004,162(1):9-24.
[13]CRAINE J M.Competition for nutrients and optimal root allocation[J].PlantandSoil,2006,285(1-2),171-185.
[14]MCGUIRE J P,MITCHELL R J,MOSER E B,etal.Gaps in a gappy forest:plant resources,longleaf pine regeneration,and understory response to tree removal in longleaf pine savannas[J].CanadianJournalofForestResearch,2001,31(5):765-778.
[15]DECHANTAL M,LEINONEN K,KUULUVAINEN T,etal.Early response ofPinussylvestrisandPiceaabiesseedlings to a experimental canopy gap in a boreal spruce forest[J].ForestEcologyandManagement,2003,176(1):321-336.
[16]XIAN J R(鮮駿仁),HU T X(胡庭興),ZHANG Y B(張遠彬),etal.Effects of forest canopy gap onAbiesfaxonianaseedling's biomass and its allocation in subalpine coniferous forest of West Sichuan[J].ChineseJournalofAppliedEcology(應用生態學報),2007,18(4):721-727(in Chinese).
[17]WU Q G(吳慶貴),WU F ZH(吳福忠),YANG W Q(楊萬勤),etal.Characteristics of gaps anddisturbance regimes of the alpine fir forest in Western Sichuan[J].J.Appl.EnvironBiol.,2013,19(06):922-928.
[18]HUGHES M K,FUNKHOUSER G.Frequency-dependent Climate Signal in Upper and Lower Forest Border Tree Rings in the Mountains of the Great Basin[M]//Climate Variability and Change in High Elevation Regions:Past,Present & Future.Springer Netherlands,2003:233-244.
[19]XU P F(徐培富),CHEN J J(陳進軍).The effects ofdifferent cultural practice on speciesdiversity of low-beneficial cypress forests in low hill of Mianyany City[J].JournalofSichuanForestryScienceandTechnology(四川林業科技),2009,30(5):88-94(in Chinese).
[20]XIAN J R(鮮駿仁),HU T X(胡庭興),WANG K Y(王開運),etal.Studies on the characteristics of gap border trees in the subalpine coniferous forest in Western Sichuan [J].ForestResearch(林業科學研究),2004,5:636-640(in Chinese).
[21]FITTER A H,STICKLAND T R.Architectural analysis of plant root systems.Ⅲ.Studies on plans under field conditions[J].NewPhytologist,1992,121(2):243-248.
[22]HODGE A.The plastic plant:root responses to heterogeneous supplies of nutrients[J].NewPhytologist,2004,162(1):9-24.
[23]PREGITZIER K S,DEFOREST J L,BURTON A J,etal.Fine root architecture of nine north American trees[J].EcologicalMonographs,2002,72(2):293-309.
[24]SHI W(師偉),WANG ZH Q(王政權),LIU J Q(劉金梁),etal.Fine root morphology of twenty hardwood species in Maoer shan natural secondary forest in northeastern China[J].JournalofPlantEcology(植物生態學報),2008,32(6):1 217-1 226(in Chinese).
[25]LIU J(劉佳),XIANG W H(項文化),XU X(徐曉),etal.Analysis of architecture and functions of fine roots of five subtropical tree species in Huitong,Hunan Province,China[J].JournalofPlantEcology(植物生態學報),2010,34(8):938-945(in Chinese).
[26]HENDRICK R L,PREGITZER K S.Thedynamics of fine root length,biomass,and nitrogen content in two northern hardwood ecosystems[J].CanadianJournalofForestResearch,1993,23(12):2 507-2 520.
[27]YU S Q,WANG Z Q,SHI J W,QUAN X K,etal.Estimating fine-root longevity of and using mini-rhizotrons[J].JournalofPlantEcology,2007,31(1):102-109.
[28]NOGUCHI K,HAN Q,ARAKI M G,etal.Fine-rootdynamics in a young hinoki cypress (Chamaecyparisobtusa) stand for 3 years following thinning[J].JournalofForestResearch,2011,16(4):284-291.
[29]LIU J L(劉金梁),MEI L(梅莉),GU C C(谷加存),etal.Effects of nitrogen fertilization on fine root biomass and morphology ofFraxinusmandshuricaandLarixgmelinii:A study with in-growth core approach[J].ChineseJournalofEcology(生態學雜志),2009,28(1):1-6(in Chinese).
[30]PERSSON H,MAJDI H,CLEMENSSON-LINDELL A.Effect of aciddeposition on tree roots[J].EcolBull,1995,44:158-167.
[31]PERSSON H,AHLSTROM K.Fine root response to nitrogen supply in nitrogen manipulated Norway spruce catchment areas[J].ForestEcologyandManagement,2002,168(1),29-41.
[32]BAYALA J,TEKLEHAIMANNOT Z,OUEDRAOGO S J.Fine rootdistribution of pruned trees and associated crops in a parkland system in Burkina Faso[J].AgroforestrySystems,2004,60(1):13-26.
[33]EISSENSTATd M,YANAI Rd.The ecology of root lifespan[J].AdvancesinEcologicalResearch,1997,27:1-60.
[34]CANHAM C A,FROEND R H,STOCK Wd,etal.Dynamics of phreatophyte root growth relative to a seasonally fluctuating water table in a Mediterranean-type environment[J].Oecologia,2012,170(4):909-916.
[35]MAKITA N,ISHII H,KANAZAWA Y,etal.Very fine roots respond to soildepth biomass allocation,morphology,and physiology in a broad leaved temperate forest[J].EcologicalResearch,2011,(1),26:95-104.
[36]WEI P(魏鵬),LI X W(李賢偉),FAN CH(范川),etal.Fine root biomass and carbon storage in surface soil ofCinnamomumcamphoraplantation in Rainy Area of West China[J].ChineseJournalofAppliedEcology(應用生態學報),2013,24(10):2 755-2 762(in Chinese).
[37]LI R X(李瑞霞),PENG T T(彭婷婷),LIU N(劉娜),etal.Response of carbon storage,fine root biomass and morphology to thinning inPlatycladusorientalisplantation[J].JournalofNortheastForestryUniversity(東北林業大學學報),2013,41(5):33-37(in Chinese).
[38]GUOd L,XIA M X,WEI X,etal.Anatomical traits associated with absorption and mycorrhizal colonization are linked to root branch order in twenty-three Chinese temperate tree species[J].NewPhytol.,2008,180(3):673-683.
[39]GUOd L,MITCHELL R J,HENDRICKS J J.Fine root branch orders responddifferentially to carbon source-sink manipulations in a longleaf pine forest[J].Oecologia,2004,140(3):450-457.
[40]WANG ZH Q(王政權),ZHANG Yd(張彥東).Study on the root intreactions betweenFraxinusmandshuricandLarixgmelinii[J].ActaPhytoecologicaSinica,2000,24(3):346-350(in Chinese).
[41]WANG Z H(王祖華),LI R X(李瑞霞),HAO J P(郝俊鵬),etal.Effects of thinning on fine root morphology in Chinese Fir plantations[J].JournalofNortheastForestryUniversity(東北林業大學學報),2011,39(6):13-15(in Chinese).
(編輯:潘新社)
猜你喜歡
中國綠色畫報(2016年11期)2017-02-18 15:21:32
中州學刊(2016年11期)2017-01-06 02:17:50
中國綠色畫報(2016年7期)2016-12-26 16:37:18
藝術科技(2016年10期)2016-12-14 20:02:46
文藝生活·下旬刊(2016年11期)2016-12-12 08:14:33
商情(2016年40期)2016-11-28 11:50:53
藝術科技(2016年9期)2016-11-18 18:35:07
人間(2016年27期)2016-11-11 15:56:02
中國科技博覽(2016年13期)2016-07-13 02:25:11
科技視界(2016年7期)2016-04-01 12:24:45
