水稻短根突變體Osksr5的遺傳分析和基因定位
2016-02-06 03:25:20鄭文娟陳紅梅朱世華丁沃娜
西北植物學報 2016年12期
關鍵詞:水稻
鄭文娟,陳紅梅,葉 金,朱世華,丁沃娜
(寧波大學 科學技術學院,浙江寧波 315212)
?
水稻短根突變體Osksr5的遺傳分析和基因定位
鄭文娟,陳紅梅,葉 金,朱世華,丁沃娜*
(寧波大學 科學技術學院,浙江寧波 315212)
水稻根系對其生長、發育及產量等起著至關重要的作用。該研究從甲基磺酸乙酯(ethyl methane sulfonate,EMS)誘變的秈稻Kasalath突變體庫中篩選到1個根系變短的突變體,命名為Osksr5(Oryzasativakasalathshortroot5),該突變體植株具體表現為主根、不定根和側根都明顯變短,不定根的數目相對減少,株高與野生型相比也明顯矮小。遺傳分析結果表明,該突變性狀由1對隱性核基因控制。利用圖位克隆技術將OsKSR5基因定位在第1染色體的STS(sequence tagged site)分子標記33027k和33471k,物理距離約為444 kb。對OsKSR5基因的定位為進一步克隆該基因和闡明水稻根系發育的分子機理奠定了基礎。
水稻;短根;遺傳分析;基因定位
Genetic Analysis and Gene Mapping of a Short Root Mutant
水稻(OryzasativaL.)是中國最主要的糧食作物之一。根據中國國家統計局統計,2014年全國稻谷產量為20 642.7萬t,占糧食總產量的34.00%;2015年全國稻谷產量為20 824.5萬噸,占糧食總產量的33.51%。但中國糧食安全受到耕地面積持續縮減和自然災害的危險。因此,提高水稻的單產,對于中國糧食安全和國民經濟穩定發展具有特別重要的意義。中國水稻產量經歷過矮化育種和雜交水稻兩次飛躍后,還未再次實現大幅度的突破。秈稻與粳稻2個亞種的基因組序列測序的完成,為尋找農藝性狀相關基因提供了基礎,也提高了通過基因改造培育高產水稻的可能性。
根系能從土壤中吸收水分、養分及合成和分泌生理活性物質,為水稻提供生長的物質基礎,調節地上部生長狀況。水稻根系為須根系,一般由胚根、不定根和側根組成。根型由長度、數量和空間范圍等幾個參數決定,研究發現根長決定了其他根系性狀的遺傳表現,是所有根系性狀中最重要性狀。水稻單株產量和品質等7個地上部農藝性狀與根長呈極顯著正相關[1-6]。2005年吳偉明等[7]提出了水稻根系育種的理論和概念,以達到水稻高產、穩產和高效等目的。根系育種需要傳統育種和相應的根系分子遺傳學機制相結合。至今已報道了40個左右水稻根系基因被鑒定并克隆,通過這些基因的功能鑒定,對水稻根系生長發育的分子機理研究有了一定程度的認識,但相對于水稻地上部位器官的研究程度而言,還是遠遠不夠的,需要挖掘更多的根系突變體,克隆更多的功能基因,為水稻的根系育種打下堅實的分子遺傳學基礎。
本研究的短根突變體Osksr5是從EMS誘變的秈稻Kasalath突變體庫中篩選而來,該突變體苗期的植株表現為株高與野生型相比明顯矮小,主根、不定根和側根都明顯變短,不定根的數目也相對減少。本研究對Osksr5進行了初步的表型鑒定、遺傳分析和基因定位,為進一步克隆該基因和研究其功能奠定了基礎。
1 材料和方法
1.1 水稻材料及培養條件
水稻短根突變體Osksr5是由秈稻Kasalath經EMS誘變后篩選得到。培養條件:光照12 h,濕度70%,白天30 ℃,夜晚22 ℃。水稻培養液的配方參照Yoshida 等[8]。
1.2 表型分析
培養7 d后,觀察突變體Osksr5和野生型Kasalath表型。用相機(Nikon D5000)進行整個植株及根部的拍攝,用Leica MZ95對水稻根部進行觀察。用標尺分別測量株高、主根根長和不定根長以及統計不定根的數目,樣本統計數為20株。
土池栽培實驗于寧波大學科學技術學院人工土池中進行,野生型和突變體以同樣的密度栽培于同一地塊中,成熟期時分別統計野生型和突變體的抽穗期,測定株高、分蘗數、千粒重和結實率等指標。樣本統計數為20株。
1.3 遺傳分析和定位群體的構建
將突變體Osksr5與野生型Kasalath雜交獲得F1,觀察F1表型并判斷顯隱性。F1自交獲得F2,培養7 d后觀察并統計F2植株長短根的分離比情況并進行卡方檢測。突變體Osksr5與粳稻日本晴Nipponbare雜交,F1自交獲得的F2群體里分離出來的短根植株用于基因定位。
1.4 分子標記分析
1.4.1 分子標記的選擇和設計 用于基因定位的分子標記來自已公布的SSR(simple sequence repeat)序列(http://www.gramene.org/),每條染色體按照20 cM的遺傳距離較均勻合理地選擇約10個SSR標記進行粗定位。通過比較已公布的‘日本晴’和9311的核酸序列,對差異部位設計STS標記對OsKSR5進行進一步定位。SSR和STS標記引物均由上海生工生物工程有限公司合成。
1.4.2 PCR擴增 采用簡易TPS法[9],從水稻葉片中提取親本、F1和F2群體分離的突變株DNA。10 L PCR擴增體系:1 L 模板DNA、1 L 10×PCR buffer、1.2 L 25 mmol/L MgCl2、0.3 L 2.5 mmol/L dNTP、0.3 L 10 mol/L上下游引物、0.1 L 5U/LTaqDNA聚合酶、5.8 L超純水。PCR條件:94 ℃預變性4 min;94 ℃ 變性30 s,55 ℃ 退火30 s,72 ℃延伸 30 s,34個循環;72 ℃延伸 5 min。PCR產物用于6%非變性聚丙烯酰胺凝膠190 V電泳2.5 h,銀染后顯色分析并拍照。
2 結果與分析
2.1 突變體的表型鑒定
選取培養了7 d的野生型Kasalath和純合突變體Osksr5進行表型觀察,發現該突變體Osksr5地上部分的株高比野生型植株矮小,株高是野生型的58.87%;根部性狀比較發現,突變體主根和不定根根長明顯變短,長度分別只有野生型的44.44%和53.44%(表1)。體視鏡下觀察發現,該突變體的側根數目比野生型少,側根長度與野生型比較也變短(圖1)。
成熟期的突變體相比野生型植株整體矮小,根系也欠發達(圖2),株高是野生型的59.90%(圖2),抽穗期比野生型晚1星期左右,分蘗數僅為野生型的25.17%,千粒重和結實率分別是野生型的92.58% 和88.82%(表2)。
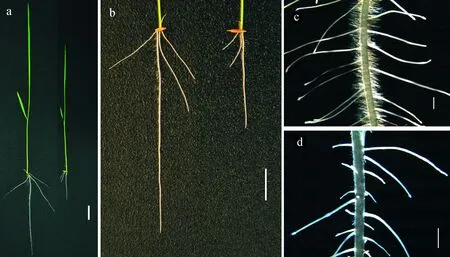
a. 野生型WT(左)和突變體Osksr5 (右)的全株表型(標尺=2 cm);b. 野生型(左)和突變體(右)的根部表型(標尺=2 cm);c和d:野生型和突變體主根的體視鏡照(標尺=2 mm)圖1 水稻野生型Kasalath和突變體Osksr5培養7 d的表型a. Seedlings of the wild type (left) and Osksr5 mutant (right), bar=2 cm; b. The root of WT (left) and Osksr5 mutant (right), bars=2 cm; c,d. The primary root of the wild type (c) and Osksr5 mutant (d) under stereoscope, bar=2 mmFig.1 Phenotypic characterization of 7-day old seedlings of the wild type and Osksr5 mutant

性狀Trait野生型WT突變體Osksr5主根長Primaryrootlength/cm12.61±0.045.60±0.61不定根長Adventitiousrootlengtha/cm3.63±0.461.94±0.31不定根數Adventitiousrootnumber4.6±0.593.1±0.22株高Height/cm14.12±0.558.33±0.86
注:a最長3根不定根長度的平均值
Note:aAverage of three longest adventitious roots on each plant
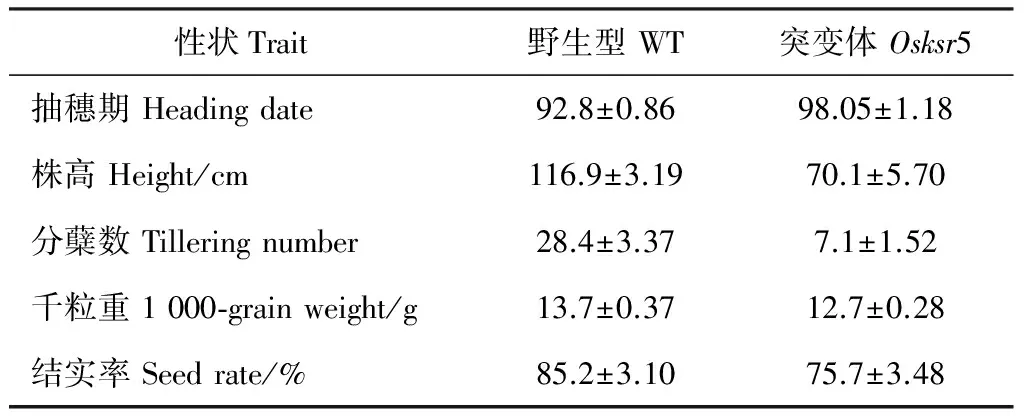
表2 野生型和突變體Osksr5的農藝性狀參數

圖2 野生型Kasalath(左)和突變體Osksr5(右)成熟期全株表型(標尺=10 cm)Fig.2 Mature phenotypes of the wild type(left) and Osksr5 mutant (right)
2.2 遺傳分析
2.3 OsKSR5基因定位
選用純合突變體Osksr5和日本晴Nipponbare作為親本雜交,獲得F2群體中表現為短根的群體作為定位群體。在F2分離群體中隨機選取30株短根表型,將這30株進行單株提取DNA,然后取等量DNA混合均勻構建一個突變池。將突變池作為模板,Kasalath、Nipponbare和F1作為對照,從均勻分布在水稻12條染色體的SSR標記的引物進行擴增后,發現OsKSR5與第1號染色體上的RM3240引物連鎖(圖3)。根據Nipponbare和9311的基因組序列存在的差異性,在RM3240附近的位置處,設計發展出新的8對STS標記(表3),在799株F2代分離的短根植株182株為模板,PCR擴增RM3240和新設計的STS引物,最終把OsKSR5基因定位在33027k和33471k之間,物理距離大約為444 kb(圖4和圖5)。
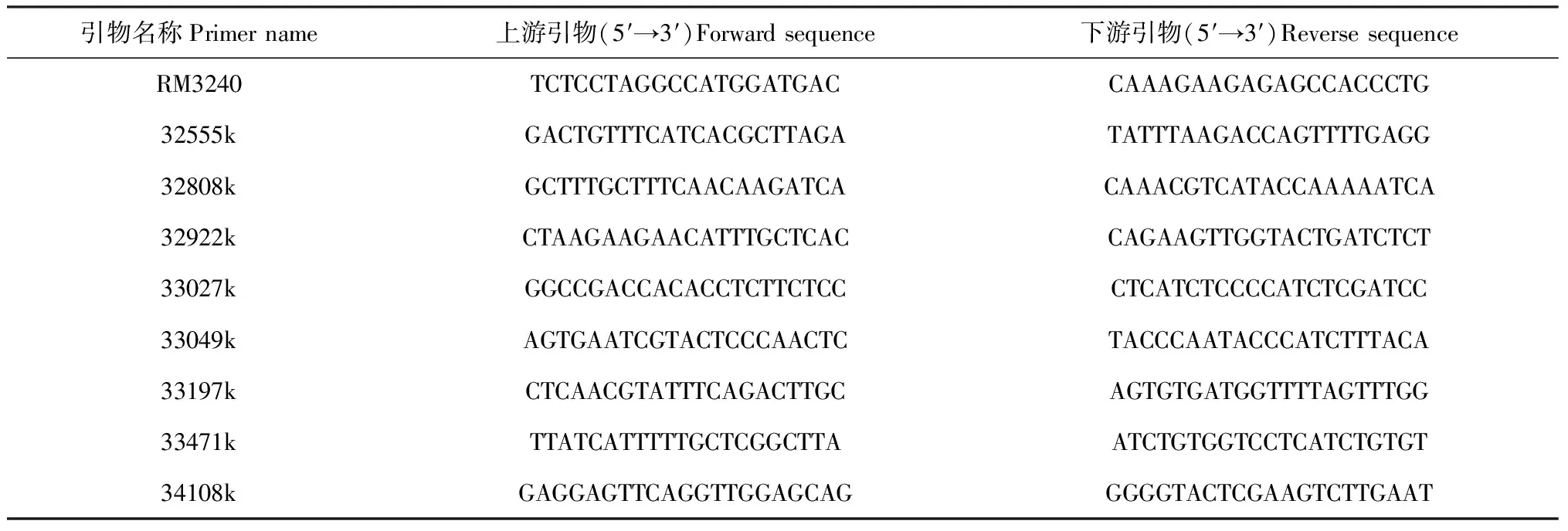
表3 具有多態性的SSR及STS標記序列
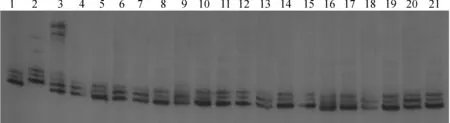
1. Kasalath;2.Nipponbare;3.F1(Osksr5×Nipponbare);4~21. F2短根單株圖3 與OsKSR5基因連鎖的SSR標記RM3240的擴增結果1. Kasalath;2.Nipponbare;3.F1(Osksr5×Nipponbare);4~21. Short root individuals in F2 populationFig.3 Electrophoresis of the SSR marker RM3240 linked with the OsKSR5 gene
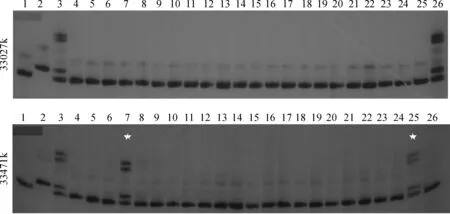
1. Kasalath;2.Nipponbare;3. F1(Osksr5×Nipponbare);4-26. F2短根單株;*. 交換單株圖4 標記 33027k 和 33471k 在日本晴/Osksr5 F2 群體中分離的部分擴增結果1. Kasalath;2.Nipponbare; 3. F1(Osksr5×Nipponbare); 4-26. Short root individuals in F2 population; *. Exchange plantFig.4 Marker 33027k and 33471k in the partial F2 population of Nipponbare/Osksr5
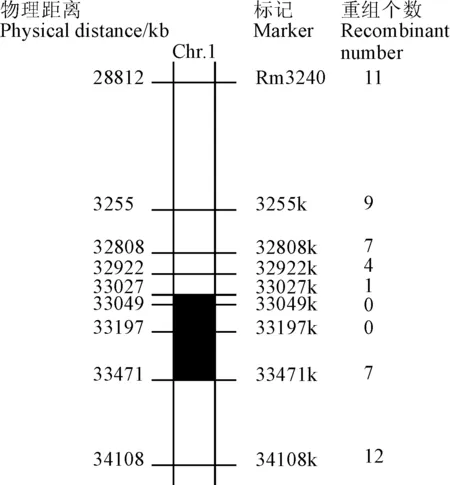
圖5 OsKSR5基因在水稻第1號染色體上的分子定位Fig.5 The gene mapping of OsKSR5 on rice chromosome 1
3 討 論
根系是水稻的三大器官之一,是水稻與土壤進行物質交換和信息交流的紐帶,將吸收的水分、養分等供給地上部分生長和發育,因此良好的根系是水稻高產和穩產的基礎。
目前已報道的被鑒定并克隆水稻根系相關基因,主要涉及主根,不定根,側根以及根毛的起始和發育,而這些基因大多是與生長素、細胞分裂素、茉莉酸等幾大主要激素的信號傳遞通道相關,另外,還與細胞壁多糖和纖維素合成、蔗糖代謝,金屬離子等相關[10-11]。各個相關基因所控制的表型包括根的長度、數量和角度等。本研究鑒定的突變體Osksr5植株表現為主根、側根和不定根都明顯變短,不定根的數目相對減少,與已報道的水稻根系發育相關基因所控制表型類似的有:OsWOX11(第7號染色體)[12]、GLR3.1(第4號染色體)[13]、OsGNA1(第9號染色體)[14]、Oscyt-inv1(第2號染色體)[15]、OsGLU3(第4號染色體)[16]、OsDGL1(第7號染色體)[17]和OsSPR1(第1號染色體)[18]等。
OsWOX11基因可能是生長素和細胞分裂素信號傳導的整合器,并通過細胞分裂素應答基因RR2調節不定根發育過程中的細胞增殖,該基因發生突變后表現主根變短,不定根稀少甚至缺失[12]。為進一步解析OsWOX11在調控水稻不定根發育中的作用機理,2015年該研究組分離并鑒定了1個與WOX11相互作用的AP2/ERF類轉錄因子ERF3。研究表明ERF3和WOX11調控著一個共同基因RR2。在不定根起始發育時期,ERF3通過促進RR2的表達控制不定根的起始,而在不定根伸長生長過程中,ERF3與WOX11相互作用,促進WOX11對RR2的抑制來促進不定根的伸長生長。揭示了水稻不定根從起始到伸長過程中存在著一套對細胞分裂素信號途徑的精細調控模式[19]。GLR3.1編碼一個類谷氨酸受體蛋白,對水稻幼苗早期根尖分生組織維持細胞分裂和單個細胞存活是不可或缺的。該基因突變體的主根、不定根和側根都變短,組織學和DNA合成分析表明突變體根的分生組織活性喪失,伴隨著細胞程序性死亡提高[13]。OsGNA1基因編碼葡糖胺-6-磷酸乙酰轉移酶,是維持水稻根細胞形狀所必需的。gna1突變體表現出在根伸長過程中對溫度敏感的缺陷:主根、不定根和根毛都變短,在32 ℃為野生型的70%;而23 ℃時根長變得很短,根中微管破裂,根伸長區細胞皺縮[14]。Oscyt-inv1基因與擬南芥AtCyt-inv1同源,編碼一個堿性/中性轉化酶,Oscyt-inv1突變體中蔗糖含量增加而己糖含量降低,突變體植株表現為主根、不定根和側根變短成短根表型。切片分析發現突變體沿縱軸方向細胞的長度減小,根的伸長區細胞皺縮。外施葡萄糖能夠恢復Oscyt-inv1突變體的根生長。這些結果表明OsCyt-inv1對水稻根系細胞發育和生殖發育有著重要的作用[15]。OsGLU3基因編碼水解纖維素β-1,4-糖苷鍵的β-1,4-葡聚糖內切酶,參與根細胞壁纖維素合成,通過影響根細胞的分裂及細胞的伸長來調控根系的發育,突變體Osglu3的主根和不定根長度變短[16]。OsDGL1編碼多萜長醇二磷酸寡糖蛋白環糊精糖基轉移酶亞基。OsDGL1是形成寡糖轉移酶(OST)復合體所必需的,參與水稻N-糖基化和根的發育。該突變體中,根中N糖基化過程受損,根中細胞壁的多糖基質發生變化,根細胞長度變短,根分生組織變小,根細胞發生死亡[17]。OsSPR1編碼一種參與根的伸長和金屬離子的動態平衡的新的線粒體蛋白,具有Armadillo-like重復域。與野生型相比,突變體幼苗中對鐵、錳和鋅的吸收受到抑制,植株表現為主根、不定根和側根的伸長受到嚴重的抑制,呈短根系表型[18]。
本實驗經圖位克隆技術最終將OsKSR5定位在第1號染色體的分子標記33027k和33471k之間,物理距離大約為444 kb,目前在第1號染色體上已克隆的與根系發育相關的基因只有OsSPR1[18],OsRAA1[20]和OsSRH3[21]。OsSPR1雖然其突變體植株表型與本研究相似,但和OsKSR5不在同一區間。OsRAA1過量表達的轉基因株系中,主根生長被抑制,不定根數目增加,主根螺旋卷曲,幼苗中根對重力的響應延遲,其物理位置位于8 586 049 bp和8 586 941 bp之間[20];ossrh3突變體根毛伸長嚴重受阻,主根和不定根略變短,側根變長,該基因的位置位于為38 978 883 bp 和39 016 640 bp[21],OsRAA1和OsSRH3基因所調控的表型和區間都和OsKSR5不一樣,同時生物信息學分析表明,在定位的444 kb區間內沒有已知的與根長相關的基因,因此OsKSR5是一個新的根系發育相關基因,對OsKSR5基因的定位為進一步該基因克隆,功能研究和闡明水稻根系發育的分子遺傳機理奠定一個基礎。
[1] 凌啟鴻, 凌 勵. 水稻不同層次根系的功能及對產量形成作用的研究[J]. 中國農業科學, 1984,17(5): 3-11.
LING Q H, LING L. Studies on the functions of roots at different nodepositions and their relation to the yield formation in rice plants[J].ScientiaAgriculturaSinica, 1984, 17(5): 3-11.
[2] 川田信一郎.水稻的根系[M]. 申廷秀, 劉執鈞, 彭望瑗, 譯.北京:農業出版社, 1984: 1.
[3] MORITA S, IWABUCHI A, YAMAZAKI K. Relationships between the growth direction of primary roots and yield in rice plants[J].JapaneseJournalofCropScience, 1996, 55: 520-525.
[4] 吳偉明, 宋祥甫, 孫宗修, 等. 不同類型水稻的根系分布特征比較[J]. 中國水稻科學, 2001,15(4): 276-280.
WU W M, SONG X F, SUN Z X,etal. Comparison of root distribution between different type rice[J].ChineseJournalofRiceScience, 2001,15(4): 276-280.
[5] 蔡昆爭, 駱世明, 段舜山. 水稻根系的空間分布及其與產量的關系[J]. 華南農業大學學報(自然科學版), 2003,24(3): 1-4.
CAI K Z, LUO S M, DUAN S S. The relationship between spatial distribution of rice root system and yield[J].JournalofSouthChinaAgriculturalUniversity(Natural Science Edition), 2003,24(3): 1-4.
[6] 張 耗, 黃鉆華, 王靜超, 等. 江蘇中秈水稻品種演進過程中根系形態生理性狀的變化及其與產量的關系[J]. 作物學報, 2011,37(6): 1 020-1 030.
ZHANG H, HUANG Z H, WANG J C,etal. Changes in morphological and physiological traits of roots and their relationships with grain yield during the evolution of mid-season indica rice cultivars in Jiangsu Province[J].ActaAgronomicaSinica, 2011,37(6): 1 020-1 030.
[7] 吳偉明, 程式華. 水稻根系育種的意義與前景[J]. 中國水稻科學, 2005,19(2): 174-180.
WU W M, CHENG S H. Significance and prospects of breeding for root system in rice(Oryzasativa)[J].ScientiaAgriculturaSinica, 2005,19(2): 174-180.
[8] YOSHIDA S, FORNO D A, COCK J H. Laboratory manual for physiological studies of rice[M]. Manila, Philippines: The International Rice Research Institute, 1976: 62.
[9] 張向前, 鄒金松, 朱海濤, 等. 水稻早熟多子房突變體fon5的遺傳分析和基因定位[J]. 遺傳, 2008,30(10): 1 349-1 355.
ZHANG X Q, ZOU J S, ZHU H T,etal. Genetic analysis and gene mapping of an early flowering and multi-ovary mutant in rice(OryzasativaL.)[J].Hereditas, 2008,30(10): 1 349-1 355.
[10] WU W M, CHENG S H. Root genetic research, an opportunity and challenge to rice improvement[J].FieldCropsResearch, 2014, 165: 111-124.
[11] MAI C D, PHUNG N T, TO H T,etal. Genes controlling root development in rice[J].Rice, 2014, 7: 30.
[12] ZHAO Y, HU Y, DAI M Q,etal. The WUSCHEL-related homeobox gene WOX11 is required to activate shoot-borne crown root development in rice[J].ThePlantCell, 2009,21(3): 736-748.
[13] LI J, ZHU S H, SONG X W,etal. A rice glutamate receptor-like gene is critical for the division and survival of individual cells in the root apical meristem[J].ThePlantCell, 2006,18(2): 340-349.
[14] JIANG H W, WANG S M, DANG L,etal. A novel short-root gene encodes a glucosamine-6-phosphate acetyltransferase required for maintaining normal root cell shape in rice[J].PlantPhysiology, 2005,138(1): 232-242.
[15] JIA L Q, ZHANG B T, MAO C Z,etal. OsCYT-INV1 for alkaline/neutral invertase is involved in root cell development and reproductivity in rice (OryzasativaL.)[J].Planta, 2008,228(1): 51-59.
[16] ZHANG J W, XU L, WU Y R,etal. OsGLU3, a putative membrane-bound endo-1,4-beta-glucanase, is required for root cell elongation and division in rice (OryzasativaL.)[J].MolecularPlant, 2012,5(1): 176-186.
[17] QIN C, LI Y Y, GAN J,etal. OsDGL1, a homolog of an oligosaccharyltransferase complex subunit, is involved in N-glycosylation and root development in rice[J].PlantandCellPhysiology, 2013,54(1): 129-137.
[18] JIA L Q, WU Z C, HAO X,etal. Identification of a novel mitochondrial protein, short postembryonic roots 1 (SPR1), involved in root development and iron homeostasis inOryzasativa[J].TheNewPhytologist, 2011,189(3): 843-855.
[19] ZHAO Y, CHENG S, SONG Y,etal. The interaction between rice ERF3 and WOX11 promotes crown root development by regulating gene expression involved in cytokinin signaling[J].PlantCell, 2015,27(9): 2 469-2 483.
[20] GE L, CHEN H, JIANG J F,etal. Overexpression of OsRAA1 causes pleiotropic phenotypes in transgenic rice plants, including altered leaf, flower, and root development and root response to gravity[J].PlantPhysiology, 2004,135(3): 1 502-1 513.
[21] 丁沃娜, 黃 煒, 寧永強, 等. 一個新的水稻短根毛突變體的遺傳分析和基因定位[J].作物學報,2012,38(3):429-435.
DING W N, HUANG W, NING Y Q,etal. Genetic analysis and gene mapping of a novel short root hair mutant in rice[J].ActaAgronomicaSinica, 2012, 38(3): 429-435.
(編輯:宋亞珍)
Osksr5 in Rice (OryzasativaL.)
ZHENG Wenjuan, CHEN Hongmei, YE Jin, ZHU Shihua, DING Wona*
(College of Science and Technology Ningbo University,Ningbo,Zhejiang 315212,China)
The root system plays vital roles in plant growth, development and yield. In this study, a rice mutant with significantly short roots was isolated from an EMS (ethyl methane sulfonate) generated rice mutant library in theKasalathbackground, designated asOsksr5(Oryzasativakasalathshortroot5).Osksr5 showed a dwarf phenotype, the elongation of primary roots, adventitious roots and lateral roots in the mutant was severely impaired, and the number of adventitious roots was reduced. Genetic analysis indicated that the mutant phenotype was controlled by a single recessive nuclear gene. Map-based cloning analysis locatedOsKSR5 to a 444 kb region between STS(sequence tagged site)markers 33027k and 33471k on chromosome 1. This result will be helpful for the cloning ofOsKSR5 and further characterization of molecular genetic mechanisms underlying root architecture in rice.
rice; short root; genetic analysis; gene mapping
1000-4025(2016)12-2405-06
10.7606/j.issn.1000-4025.2016.12.2405
2016-08-23;修改稿收到日期:2016-10-26
國家自然科學基金(31371595, 31300246);浙江省自然科學基金(LQ16C020001)
鄭文娟(1982-),女,碩士,主要從事水稻分子生物學研究。E-mail:zhengwenjuan@nbu.edu.cn
*通信作者:丁沃娜,博士,副教授,主要從事水稻分子生物學研究。E-mail:dwn@zju.edu.cn
Q343.1+7
A
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
