基于油酸誘導的高產三萜靈芝菌絲體發酵條件優化
2016-02-17 01:57:25孫冰沁秦可欣王桂榮何國慶
食品工業科技 2016年24期
關鍵詞:產量
孫冰沁,秦可欣,王桂榮,何國慶,*
(1.浙江大學生物系統工程與食品科學學院,浙江杭州 310058;2.哈爾濱商業大學食品工程學院,黑龍江哈爾濱 150076;3.杭州市原種場,浙江杭州 311115)
基于油酸誘導的高產三萜靈芝菌絲體發酵條件優化
孫冰沁1,秦可欣2,王桂榮3,何國慶1,*
(1.浙江大學生物系統工程與食品科學學院,浙江杭州 310058;2.哈爾濱商業大學食品工程學院,黑龍江哈爾濱 150076;3.杭州市原種場,浙江杭州 311115)
本文對油酸促進靈芝菌絲體高產靈芝三萜發酵條件進行了優化,旨在提高三萜這一具有特殊的生物活性且對人體有益物質的產量。實驗進行了碳源、氮源等單因素的篩選,利用CCD實驗設計對培養基進行優化,以及對發酵條件單因素進行了優化。實驗結果表明,促進靈芝菌絲體高產靈芝三萜的最優結果為:培養基成分為可溶性淀粉3.4%、魚蛋白胨0.72%、KH2PO40.3%、MgSO40.15%、VB10.005%(m/v),pH5.5,在發酵培養第0 d加油酸4.38%(v/v);裝液量為100 mL,接種量為10%(v/v),發酵天數為9 d,胞內三萜產量達到(282.75±3.76)mg/100 mL。該方法不僅大幅度提高了三萜產量,對于靈芝的工業化生產也有重要意義。
靈芝菌絲體,三萜,發酵條件,響應面
靈芝(Ganodermalucidum)是一種大型藥用真菌,具有重要的保健和藥用價值,在我國有著悠久的應用歷史[1]。現代研究表明,靈芝中含有較多的功效成分,包括多糖類、三萜類、核苷類、幽醇類、生物堿類、吠喃衍生物、氨基酸多肽類、微量元素、脂肪酸等物質[2-6]。而靈芝三萜則是靈芝中主要的生物活性成分[7]。靈芝三萜具有保肝、抗腫瘤、抗HIV-Ⅰ、HIV-Ⅱ病毒酶活性、抑制組胺釋放、抑制膽固醇合成[8-12]等生物活性,因此靈芝三萜的生產研究具有重要的意義[13]。
目前研究表明,茉莉酸甲酯、苯巴比妥、阿司匹林、銅離子和鈣離子都能夠誘導靈芝三萜合成[14]。但是靈芝產品作為一種食品,必須考慮其安全性和經濟性。而油酸作為一種單不飽和脂肪酸,存在于動植物體內,具有食用安全性[15],不僅可以誘導靈芝三萜合成,而且價格低廉,是一個良好的選擇[16]。本研究在添加油酸誘導靈芝三萜產生的前提下,優化發酵條件,大幅提高靈芝三萜產量,有利于高產三萜靈芝菌絲的工業化生產。
本文對油酸促進靈芝菌絲體高產靈芝三萜發酵條件進行了優化,實驗進行了碳源、氮源等單因素的篩選,利用central composite design(CCD)實驗設計對培養基進行優化,以及對發酵條件單因素進行了優化。其中CCD實驗設計的優點有:實驗所得的信息量大且直觀,適合于探索性的實驗研究;所需要的實驗次數與費用并不大,有較高的實驗精度,不僅具有篩選作用還具有預測指導作用,在實驗設計中更具優勢。
1 材料與方法
1.1 材料與儀器
美國靈芝(赤芝) 由浙江大學農業與生物技術學院陳再鳴教授饋贈;葡萄糖、磷酸二氫鉀、無水硫酸鎂、維生素B1、瓊脂、蔗糖、乳糖、可溶性淀粉、牛肉浸膏、酵母浸出粉、冰醋酸、高氯酸、氫氧化鈉 均為分析純,國藥集團化學試劑有限公司;香草醛 Sigma-Aldrich,USA;魚蛋白胨 江蘇南通東海龍生生物制品有限公司;牛骨蛋白胨 生工生物工程(上海)股份有限公司;齊墩果酸(purity≥98%) 阿拉丁試劑(上海)有限公司;斜面培養基 PDA 4.5%,瓊脂1.7%,KH2PO40.3%,MgSO40.15%,VB10.005%(m/v);種子培養基:PDB 3.5%,蛋白胨0.5%,酵母浸出粉0.3%,KH2PO40.3%,MgSO40.15%,VB10.005%(m/v),pH5.5。
MULTISKAN GO新一代全波長酶標儀 Thermo,USA;JY96-IIN超聲波細胞粉碎機 寧波新芝生物科技股份有限公司;離心機DD-5M 湘儀離心機儀器有限公司;超聲波清洗器JP-040S 深圳市潔盟清洗設備有限公司;全溫度恒溫搖床ZHWY-2102 上海智城分析儀器制造有限公司;超凈工作臺SW-CJ-1D 蘇州蘇潔凈化設備公司。
1.2 實驗方法
1.2.1 菌種培養 取經試管斜面培養的菌株,挑取4~5塊1cm2菌絲塊接種到種子培養基中,種子培養基裝在容量為250 mL的錐形瓶中,每瓶內裝100 mL,置28 ℃搖床,180 r/min振蕩培養8 d[17],供實驗作種子用[18]。然后按10%(v/v)接種量將種子液接種到發酵培養基中(裝液量同種子培養基),并在發酵初期添加一定體積分數的油酸,于28 ℃,180 r/min培養6 d。
1.2.2 菌絲生物量(Mycelia Biomass)的測定 將菌株按照上述培養方法進行發酵培養,發酵6 d后,取適量菌絲加水混勻后進行抽濾,分離菌絲體和發酵液,再將獲得的菌絲體于60 ℃干燥至恒重,計算各菌株的生物量(以菌絲體干重g/100 mL發酵液計)[19]。
1.2.3 靈芝菌株液體發酵過程中胞內三萜(Intracellular Triterpenoids IT)產量的測定 采用超聲波提取方法:將靈芝菌絲用95%(v/v)乙醇清洗,去除上面附著的油酸,離心,收集菌絲,取適量菌絲按1 g/50 mL加入95%(v/v)乙醇并用細胞破碎儀破碎4 min,再置于超聲波清洗機中,60 ℃,400 W下提取1 h,并重復一次,合并上清液,定容后待測[20]。其三萜含量采用香草醛-高氯酸方法測定。
三萜產量(mg/100 mL)=每克菌絲胞內三萜含量(mg/g)×菌絲生物量(g/100 mL)
(1)
香草醛-高氯酸測定方法:取待測樣品溶液或樣品稀釋液0.1 mL于試管中,70℃加熱揮干溶劑,再加入5%(m/v)香草醛冰醋酸溶液0.2 mL,高氯酸0.5 mL,混勻后于60℃水浴中保溫20 min,取出后置于冷水中10 min,最后加入冰醋酸5 mL,于550 nm下測定吸光值[21]。
1.2.4 靈芝液體發酵培養基的優化 以三萜產量為主要指標,菌絲生物量為次要指標,對靈芝液體發酵培養基進行優化。
1.2.4.1 碳源及碳源濃度篩選 首先進行單因素篩選。本文選取了葡萄糖,蔗糖,麥芽汁,玉米粉,乳糖,可溶性淀粉6個碳源進行篩選,篩選時保持其余條件不變,分別為魚蛋白胨0.5%、KH2PO40.3%、MgSO40.15%、VB1 0.005%(m/v),pH5.5,在發酵培養第0 d加油酸3%(v/v);裝液量為100 mL/250 mL,接種量為10%(v/v),發酵天數為6 d。選出最優碳源后進行濃度篩選,分別選取碳源濃度為1%、2%、3%、4%、5%、6%、7%進行篩選,得出最優碳源濃度。
1.2.4.2 氮源及氮源濃度篩選 本文選取了豆粕汁,牛肉膏,酵母粉,麩皮汁,魚蛋白胨,牛骨蛋白胨6個氮源進行篩選,篩選時保持其余條件不變,分別為葡萄糖2%、KH2PO40.3%、MgSO4 0.15%、VB1 0.005%(m/v),pH5.5,在發酵培養第0 d加油酸3%(v/v);裝液量為100 mL/250 mL,接種量為10%(v/v),發酵天數為6d。選出最優氮源后進行氮源濃度后進行濃度篩選,分別選取氮源濃度為0.5%、1%、1.5%、2%、2.5%、3%、3.5%進行篩選,得出最優氮源濃度。
1.2.4.3 最適pH篩選 再進行最適pH的篩選。本文選取了4.5、5、5.5、6、6.5、7、7.5這7個不同的pH進行篩選,篩選時碳源、氮源為之前篩選出的最優值,其余條件不變。
1.2.4.4 油酸添加量與添加時間篩選 為了確定靈芝液體發酵過程中油酸最佳的添加時間與添加量,選取3個不同的添加時間:接種當天(即發酵第0 d),發酵第1 d,發酵第2 d,選取5種不同體積分數(1%、2%、3%、4%、5%),同時進行不添加油酸的對照實驗,來篩選油酸最佳的添加時間與添加量。篩選時碳源、氮源、pH為之前篩選出的最優值,其余條件不變。
1.2.4.5 響應面法優化靈芝液體發酵培養基 再采用CCD實驗設計優化靈芝液體發酵培養基,根據單因素實驗結果確定各變量的最低、最高水平,CCD實驗設計因素水平表如表1所示。
1.2.5 靈芝液體發酵條件的篩選
1.2.5.1 裝液量(通氣量)的篩選 本文選取了50、75、100、125、150 mL 5個不同的裝液量(通氣量)進行篩選,篩選時培養基配方為1.2.4得出的最優配方,接種量為10%(v/v),發酵天數為6 d,各條件保持不變。
1.2.5.2 接種量的篩選 本文選取了5%、7.5%、10%、12.5%、15% 5個不同的接種量進行篩選,篩選時培養基配方同上,裝液量(通氣量)為之前選出的最優值,發酵天數為6 d,各條件均保持不變。
1.2.5.3 發酵天數的篩選 本文對靈芝接種后發酵12 d過程中三萜產量與生物量變化進行檢測,得出其變化曲線,從而篩選出靈芝發酵最優時間。
1.3 數據處理
使用Origin和Design Expert軟件處理數據并進行誤差分析。
2 結果與討論
2.1 靈芝液體發酵培養基的優化
2.1.1 碳源及碳源濃度的篩選 本文選取了葡萄糖、蔗糖、麥芽汁、玉米粉、乳糖、可溶性淀粉作為碳源。由圖1可知,當碳源為可溶性淀粉時,靈芝液體發酵所得胞內三萜產量最高,而當麥芽汁作為碳源時,靈芝發酵所得生物量最高,但與可溶性淀粉相比,無明顯差異。由于胞內三萜產量為所需要的主要活性物質,其產量的高低直接影響靈芝菌絲的質量,因此,選取可溶性淀粉為靈芝液體發酵的碳源,胞內三萜產量可達到(141.45±3.02) mg/100 mL。
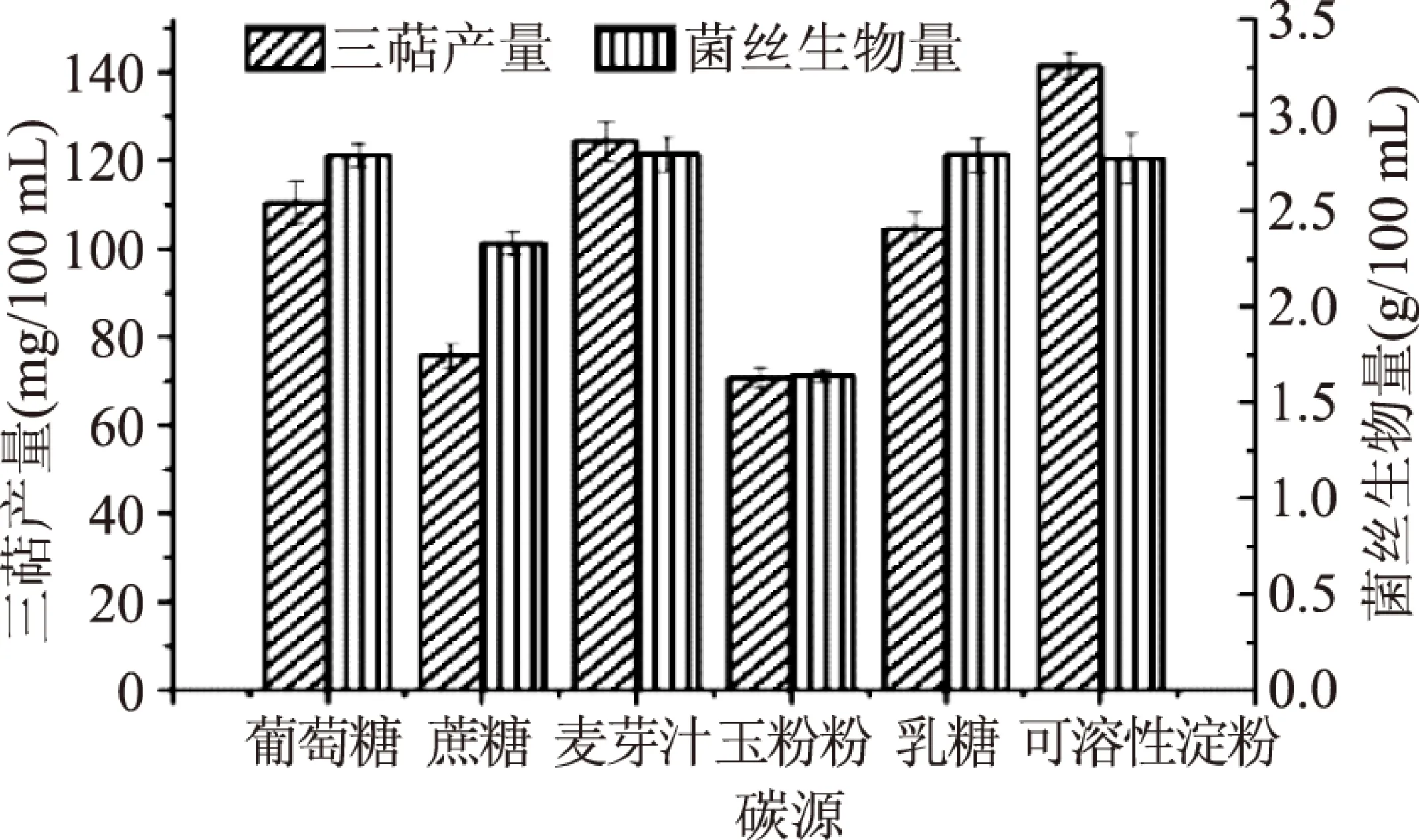
圖1 最適碳源篩選Fig.1 Optimum carbon sourc screening
由圖2可知,隨著可溶性淀粉質量濃度的增加,胞內三萜產量有所提高,當可溶性淀粉濃度為4%(m/v)時,胞內三萜產量達到最高,但當可溶性淀粉濃度為5%(m/v)時,胞內三萜產量下降。所以選取4%(m/v)可溶性淀粉作為碳源濃度,胞內三萜產量可達到(173.16±3.92) mg/100 mL。

圖2 碳源濃度篩選Fig.2 Carbon source concentration screening
2.1.2 氮源及氮源濃度篩選 本文選取了豆粕汁、牛肉膏、酵母粉、麩皮汁、魚蛋白胨、牛骨蛋白胨(作為氮源。由圖3可知,當氮源為魚蛋白胨時,靈芝液體發酵所得胞內三萜產量最高,而當麩皮汁作為氮源時,靈芝發酵所得生物量最高。因所需的主要活性物質為胞內三萜,所以選取魚蛋白胨為靈芝液體發酵的氮源,胞內三萜產量可達到(126.92±2.42) mg/100 mL。
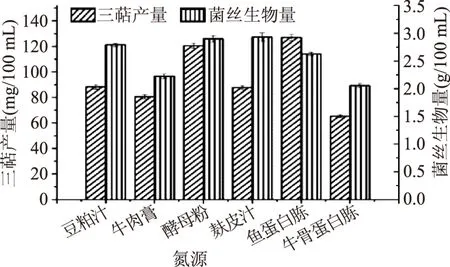
圖3 最適氮源篩選Fig.3 Optimum nitrogen source screening
由圖4可得,隨著魚蛋白胨質量濃度的增加,胞內三萜產量有所提高,當魚蛋白胨濃度為1%(m/v)時,胞內三萜產量達到最高,可達(195.88±2.17) mg/100 mL,但當魚蛋白胨濃度為1.5%(m/v)時,胞內三萜產量下降。當魚蛋白胨濃度為1.5%(m/v)時,菌絲生物量達到最高。因所需的主要活性物質為胞內三萜,所以選取1%(m/v)魚蛋白胨作為氮源濃度,胞內三萜產量可達到(195.88±2.17) mg/100 mL。

圖4 氮源濃度篩選Fig.4 Nitrogen source concentration screening
2.1.3 最適pH的篩選 由圖5可知,當發酵培養基的pH為5.5時,胞內三萜產量達到最高,而當pH增大或是減小時,胞內三萜產量降低。當pH為5時,菌絲生物量達到最高。因所需的主要活性物質為胞內三萜,所以選取5.5為靈芝液體發酵培養基的pH,胞內三萜產量可達到(206.35±2.85) mg/100 mL。
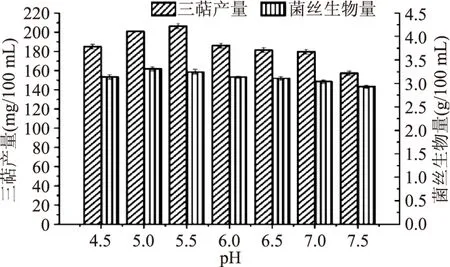
圖5 最適pH篩選Fig.5 Optimum pH screening
2.1.4 油酸添加量與添加時間的篩選 由圖6可得,添加油酸后靈芝三萜產量大幅度提高,其中發酵培養第0 d添加油酸最具優勢,胞內三萜產量與菌絲生物量均高于第1、2 d,因此,選取發酵培養第0 d為油酸添加時間。而隨著油酸添加量的增加,胞內三萜產量有所提高,當第0 d油酸添加量為4%(v/v)時,胞內三萜產量達到最高,但當油酸添加量繼續增加時,胞內三萜產量下降。菌絲生物量隨油酸添加量的增加而增加。因所需的主要活性物質為胞內三萜,選取4%(v/v)作為油酸添加量,胞內三萜產量達到(210.52±2.62) mg/100 mL。
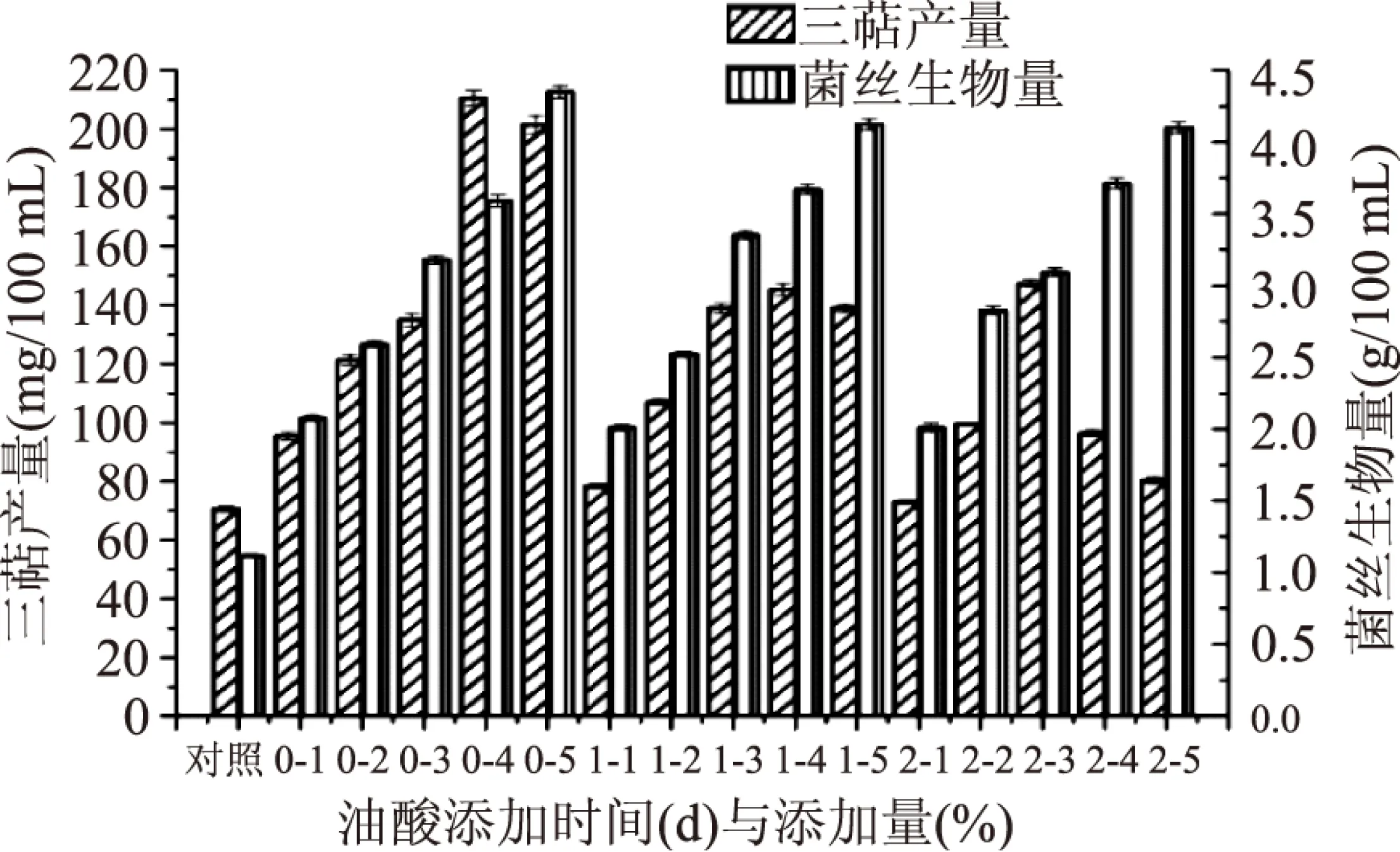
圖6 最適油酸添加量與添加時間的篩選Fig.6 Oleic acid concentration and adding time screening
2.1.5 響應面法優化靈芝液體發酵培養基 采用CCD實驗設計優化靈芝液體發酵培養基,CCD實驗設計及結果見表2,方差分析結果見表3。

表2 CCD實驗設計及結果
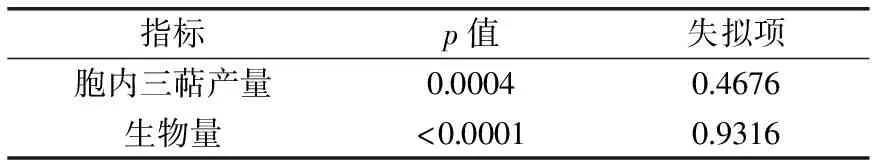
表3 方差分析表
由表3可知,對于胞內三萜產量,模型項p值為0.0004≤0.01,顯著,失擬項p值為0.4676≥0.05,不顯著;對于菌絲生物量,模型項p值<0.0001,顯著,失擬項值為0.9316≥0.05,不顯著;兩者均表明回歸方程關系顯著,且方程與實際擬合中非正常誤差所占比例小,實驗設計能夠很好的反映實驗優化。胞內三萜產量(Y1)與菌絲生物量(Y2)的多項式回歸方程如下:
Y1=211.30-11.60A-15.67B+6.82C+3.73AB-19.18AC-3.45BC-17.39A2-17.00B2-26.95C2
Y2=4.43+0.099A-0.042B+0.63C-0.058AB-4.500E-0.03AC-0.052BC+2.238E-0.04A2-0.13B2-0.045C2
固定其中一個變量,研究其他2個變量的交互作用對各響應值的影響。可溶性淀粉與魚蛋白胨、魚蛋白胨與油酸添加量的交互作用對胞內三萜的產量影響并不顯著。可溶性淀粉與油酸添加量的交互作用影響較為顯著,其對胞內三萜產量影響的曲面圖如圖7所示。由圖7可知,隨著可溶性淀粉濃度和油酸添加量的增加,胞內三萜產量會先隨之增加,再隨之降低。3個變量兩兩之間的交互作用對菌絲生物量的影響并不顯著。
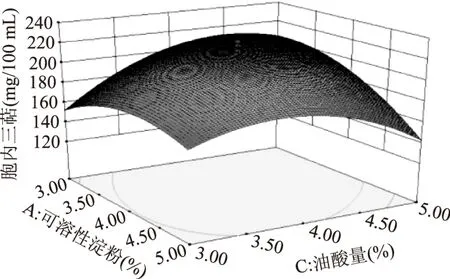
圖7 可溶性淀粉與油酸交互作用對IT產量的影響Fig.7 The contour(a)and surface(b)plots of the combined effects of soluble starch and oleic acid on the IT production
實驗模型預測:當以重量百分數計,可溶性淀粉濃度為3.40%(m/v),魚蛋白胨濃度為0.72%(m/v),油酸添加量為4.38%(v/v)時,胞內三萜產量可達到220.502 mg/100 mL。將預測結果進行驗證實驗(3次重復),所得胞內三萜產量為(223.73±2.17) mg/100 mL,與預測值接近,表明該模型對于優化靈芝發酵培養基具有可行性。因此,最優培養基條件為可溶性淀粉3.4%、魚蛋白胨0.72%、KH2PO40.3%、MgSO40.15%、VB10.005%(m/v),pH5.5,并在發酵培養第0 d添加4.38%(v/v)的油酸。
2.2 靈芝液體發酵條件的篩選
2.2.1 裝液量(通氣量)的篩選 由圖8可得,隨著裝液量的增加,胞內三萜產量隨之增加,當裝液量為100 mL時,胞內三萜產量達到最高,但當裝液量繼續增加時,胞內三萜產量下降。菌絲生物量隨裝液量的增加而降低。因所需的主要活性物質為胞內三萜,所以選取裝液量為100 mL,胞內三萜產量可達(225.21±2.23) mg/100 mL。
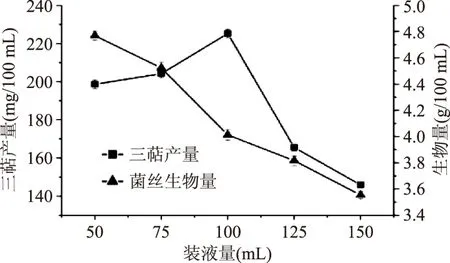
圖8 裝液量(通氣量)篩選Fig.8 Liquid loading screening
2.2.2 接種量的篩選 由圖9可知,當接種量從5%(v/v)增加到10%(v/v)時,胞內三萜產量隨接種的增加而增加,而當接種量從10%(v/v)增加到15%(v/v)時,胞內三萜產量趨于平穩并略有下降。菌絲生物量在接種量為7.5%(v/v)時達到最大。綜合考慮,選取得接種量為10%(v/v),胞內三萜產量可達(241.01±2.42) mg/100 mL。

圖9 接種量的篩選Fig.9 Inoculum concentration screening
2.2.3 發酵天數的篩選 由圖10可知,隨著發酵天數的增加,各指標均隨之增加。胞內三萜產量在發酵9 d時達到最高,可達(287.67±3.77) mg/100 mL,此時菌絲生物量達到(4.755±0.041) g/100 mL;而菌絲生物量在發酵10 d時達到最大,為(4.93±0.043) g/100 mL,其胞內三萜產量達到(282.75±3.76) mg/100 mL。綜合考慮,選擇發酵時間為9 d。
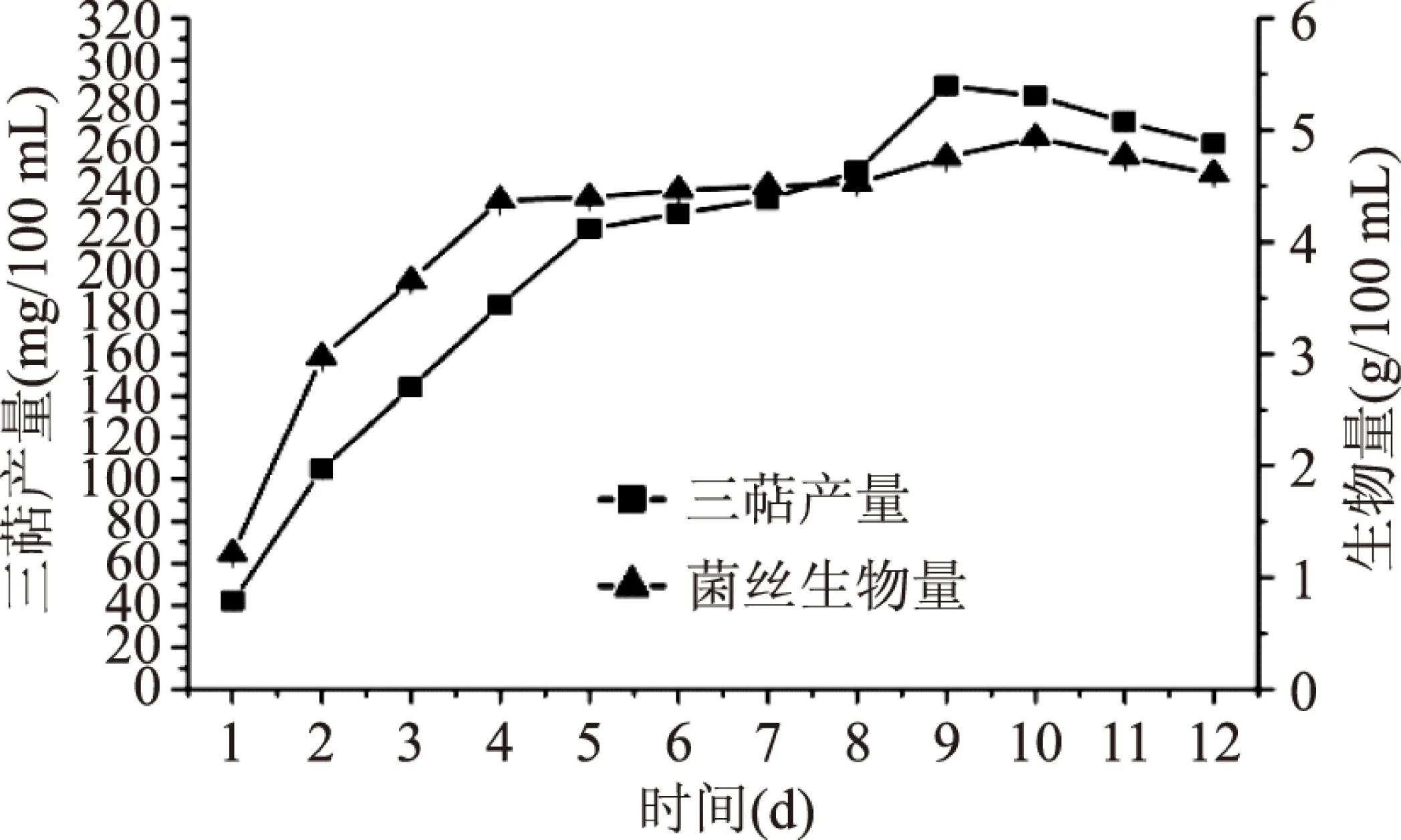
圖10 發酵天數的篩選Fig.10 Fermentation time screening
3 結論
通過以上的篩選優化,得到油酸促進靈芝菌絲體高產三萜的最優發酵條件:培養基成分為可溶性淀粉3.4%、魚蛋白胨0.72%、KH2PO40.3%、MgSO40.15%、VB10.005%(m/v),pH5.5,并在發酵培養第0 d添加4.38%(v/v)的油酸;裝液量為100 mL/250 mL,接種量為10%(v/v),發酵天數為9 d。在此條件下,靈芝菌絲體胞內三萜產量可達282.75 mg/100 mL,而不添加油酸時胞內三萜產量一般只能達到90~100 mg/100 mL[17]。該發酵條件可大幅度提高靈芝菌絲中靈芝三萜的產量,提高生產效率,有利于高產三萜靈芝菌絲的工業化生產。由于現今對靈芝三萜的生物合成途徑研究較少,油酸是如何誘導靈芝三萜合成這一關鍵點無從得知。所以,下一步將研究油酸是如何影響靈芝三萜的代謝途徑從而達到促進其產量的提高的效果,根據探索出的作用機制改進靈芝三萜合成的代謝途徑,從而獲得靈芝三萜的高產。
[1]方慶華. 靈芝真菌發酵生產靈芝酸和靈芝多糖的研究[D].上海:華東理工大學,2000.
[2]TANG Y J,J J ZHONG. Modeling the kinetics of cell growth and ganoderic acid production in liquid static cultures of the medicinal mushroom Ganoderma lucidum[J]. Biochemical Engineering Journal,2004,21(3):259-264.
[3]WANG X M,M YANG,S H GUAN,et al. Quantitative determination of six major triterpenoids in Ganoderma lucidum and related species by high performance liquid chromatography[J]. J Pharm Biomed Anal,2006,41(3):838-844.
[4]QIAO Y,X M ZHANG,M H QIU. Two novel lanostane triterpenoids from ganoderma sinense[J]. Molecules,2007,12(8):2038-2046.
[5]KEYPOUR S,H RAFATI,H RIAHI,et al. Qualitative analysis of ganoderic acids in Ganoderma lucidum from Iran and China by RP-HPLC and electrospray ionisation-mass spectrometry(ESI-MS)[J]. Food Chemistry,2010,119(4):1704-1708.
[6]林志彬. 靈芝的現代研究[M]. 北京:北京大學醫學出版社,2015:14-29.
[7]余素萍. 靈芝深層發酵生產生物活性物質的研究[D]. 南京:南京農業大學,2004.
[8]Kim H M,S Y Paik,K S Ra,et al. Enhanced production of exopolysaccharides by fed-batch culture of Ganoderma resinaceum DG-6556[J]. Journal of Microbiology,2006,44(2):233-242.
[9]ZHAO W,J W XU,J J ZHONG. Enhanced production of ganoderic acids in static liquid culture of Ganoderma lucidum under nitrogen-limiting conditions[J]. Bioresour Technol,2011,102(17):8185-8190.
[10]KANG D,M MUTAKIN,J LEVITA. Computational Study of Triterpenoids of Ganoderma lucidum with Aspartic Protease Enzymes for Discovering HIV-1 and Plasmepsin Inhibitors[J]. International Journal of Chemistry,2015,7(1):61-62.
[11]TRAN H B,A YAMAMOTO,S MATSUMOTO,et al. Hypotensive Effects and Angiotensin-Converting Enzyme Inhibitory Peptides of Reishi(Ganoderma lingzhi)Auto-Digested Extract[J]. Molecules,2014,19(9):13473-13485.
[12]Fang X,Shi L,Ren A,et al. The cloning,characterization and functional analysis of a gene encoding an acetyl-CoA acetyltransferase involved in triterpene biosynthesis in Ganoderma lucidum[J]. Mycoscience,2013,54(2):100-105.
[13]徐曉蘭. 靈芝三萜和金銀花綠原酸生物合成途徑關鍵酶基因的挖掘及分析[D]. 北京:北京協和醫學院,2013.
[14]任昂. 茉莉酸甲酯對靈芝三萜生物合成的影響及其靈芝應答基因的差異表達研究[D]. 南京:南京農業大學,2012.
[15]劉真,盧義和,宮素芝,等. 我國油酸的生產現狀及展望[J]. 河北化工,2006(9):18-20.
[16]馮杰,張勁松,楊焱,等. 一種提高靈芝液體深層發酵菌絲體中靈芝三萜含量的方法:中國,CN104017852A[P]. 2014-09-03.
[17]崔美林. 高產三萜靈芝菌株液體發酵及生物轉化大豆異黃酮的研究[D]. 杭州:浙江大學,2015.
[18]高建莉,禹志領,李紹平,等. 靈芝三萜類成分研究進展[J]. 中國食用菌,2005,4:6-11.
[19]許海燕,侯敏娜,劉劍. 靈芝總三萜提取工藝研究[J]. 陜西中醫學院學報,2007,30(3):43-44.
[20]CUI Meilin,HE Guoqing. Optimization of the Submerged Fermentation Conditions of Ganoderma lucidum with High Triterpenoids Production by Response Surface Analysis[J]. Advanced Materials Research,2012,621:259-262.
[21]AHMAD M F,B P PANDA,Z A A AZAD. Simultaneous Bioprospecting of Ganoderma lucidum OE 52 with Ganoderic Acid B and C2 by Submerged Fermentation Process[J]. Advanced Science Focus,2013,1(3):258-261.
Optimization ofGanodermalucidummycelium’s fermentation conditions with high triterpenoids production based on oleic acid induce
SUN Bing-qin1,QIN Ke-xin2,WANG Gui-rong3,HE Guo-qing1,*
(1.College of Biosystems Engineering and Food Science,Zhejiang University,Hangzhou 310058,China; 2.Food Science and Engineering College,Harbin University of Commerce,Harbin 150076,China; 3.Seed Field of Hangzhou,Hangzhou 310058)
We optimize theGanodermalucidummycelium’s fermentation conditions of high triterpenoids production with adding oleic acid,to induce the synthesis of triterpenoids and boost production,which has special biological activity and beneficial to the human body.In this research,we screened the single factors such as carbon source and nitrogen source,and the optimization of the culture medium was optimized by CCD experiment,we also screened the single factors of fermentation conditions. This research shows that the optimal results is:soluble starch 3.4%,fish peptone 0.72%,KH2PO40.3%,MgSO40.15%,VB10.005%(m/v),pH5.5,and adding 4.38%(v/v)of oleic acid at the first day of the fermentation. The liquid loading is 100 mL,the inoculation concentration is 10%(v/v),and the fermentation time is 9 d. And the production of triterpenoids was up to(282.75±3.76)mg/100 mL. This method not only induce the synthesis of triterpenoids and boost production,but also has important significance for the industrial production ofGanodermalucidum.
Ganodermalucidummycelia;triterpenoids;fermentation conditions;response surface
2016-06-15
孫冰沁(1991-),女,碩士,研究方向:食品微生物,E-mail:sbqyjyjv@sina.cn。
*通訊作者:何國慶(1957-),男,博士,教授,研究方向:食品生物技術及發酵工程,E-mail:gqhe@zju.edu.cn。
TS201.3
A
1002-0306(2016)24-0000-00
10.13386/j.issn1002-0306.2016.24.000
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
當代水產(2021年10期)2021-12-05 16:31:48
今日農業(2021年14期)2021-11-25 23:57:29
今日農業(2021年13期)2021-08-14 01:37:56
石油與天然氣地質(2021年1期)2021-02-22 14:14:44
今日農業(2020年20期)2020-11-26 06:09:10
中國果業信息(2019年10期)2019-11-13 01:21:34
中國化肥信息(2019年2期)2019-01-18 15:24:35
中國化肥信息(2019年1期)2019-01-17 21:31:12
中國化肥信息(2019年4期)2019-01-17 18:47:06
