miR-21調節缺血缺氧誘導的大鼠肝卵圓細胞自噬的研究
2016-03-30 10:53:26高勝強陳冬冬黃立棟戴瑞杰胡偉建余華軍張啟瑜單云峰溫州醫科大學附屬第一醫院肝膽外科浙江溫州325000
肝膽胰外科雜志 2016年1期
高勝強,陳冬冬,黃立棟,戴瑞杰,胡偉建,余華軍,張啟瑜,單云峰(溫州醫科大學附屬第一醫院肝膽外科,浙江溫州325000)
?
miR-21調節缺血缺氧誘導的大鼠肝卵圓細胞自噬的研究
高勝強,陳冬冬,黃立棟,戴瑞杰,胡偉建,余華軍,張啟瑜,單云峰
(溫州醫科大學附屬第一醫院肝膽外科,浙江溫州325000)
[摘 要]目的 探討miR-21在缺血缺氧誘導的大鼠肝卵圓細胞自噬中的作用。方法 體外培養肝卵圓細胞,慢病毒轉染肝卵圓細胞構建穩定的細胞株,分別提取各組細胞RNA逆轉錄后行SYBR Green實時熒光定量PCR,檢測各組miR-21的表達,以轉染好的穩定的細胞建立缺血缺氧模型,共分為3組:miR-21空載體慢病毒轉染HOC自噬組,miR-21增強慢病毒載體轉染HOC自噬組,miR-21-inhibition慢病毒載體轉染HOC自噬組。Hoechst 33258染色觀察細胞凋亡;應用丹(磺)酰戊二胺(Monodansylcadaverine,MDC)染色熒光定位法、觀察各組細胞的自噬;免疫印跡法(Western blotting)檢測各組細胞LC3-II/I蛋白表達情況。結果與空載體組比,增強組miR-21基因水平表達增強,而MDC染色減弱(自噬減少),蛋白LC3-II/LC3-I的比值減少;而抑制組miR-21基因水平表達減少,MDC染色增強(自噬增強),蛋白LC3-II/LC3-I的比值增多。結論 抑制miR-21過表達可以增強缺血缺氧引起的肝卵圓細胞的自噬,有利于肝卵圓細胞在缺血缺氧微環境中穩定細胞內環境,維持細胞的存活。
[關鍵詞]microRNA-21;自噬;肝卵圓細胞
Research of miR-21 on ischemia and hypoxia-induced autophagy modulation in rat hepatic oval cell GAO Sheng-qiang, CHEN Dong-dong, HUANG Li-dong, DAI Rui-jie, HU Wei-jian, YU Hua-jun, ZHANG Qi-yu, SHAN Yun-feng.Department of Hepatobiliary Surgery, the First Af■liated Hospital of Wenzhou Medical University, Wenzhou, Zhejiang 325000, China
Abstract ObjectiveTo investigate the role of miR-21 in ischemia-thypoxiainduced autophagic model of rat hepatic oval cells.MethodsHepatic oval cells were cultured in vitro and a stable hepatic oval cells line was constructed by lentivirus (miR-21 enhanced lentivirus; miR-21 inhibited lentivirus; vector lentivirus).The RNA was extracted from each group and reverstranscripted, then SYBR real-time PCR was conducted to detect the miR-21 expression.Simulated ischemia and hypoxia of transfected HOCs (hepatic oval cells) as an in vitro model of ischemic-hypoxic microenvironment were employed in this study.Hoechst 33258 staining was used to determine cell apoptosis.MDC (Monodansylcadaverine)-labeled autophagic vacuoles were observed under ■uorescent inverted phase contrast microscopy in HOC.The expression of LC3 and the result of LC3-I converted to LC3-II was detected with Western blotting.ResultsCompared with the empty vector group, the expression of miR-21 was increased in enhanced group and decreased in inhibited group; Compared with the empty vector group, enhanced group MDC staining was decreased, and protein LC3-II/LC3-I was reduced.Compared with the empty vector, inhibited group MDC staining was enhanced, and the ratio of protein LC3-II/LC3-I was increased.Conclusion Inhibition of miR-21 expression can enhance autophagy in ischemia-hypoxiainduced model.Autophagy of HOC can be induced by the ischemic and hypoxic microenvironment and maintain survival and the stability of cells.
Key words microRNA-21; autophagy; hepatic oval cell
肝卵圓細胞被認為是原始的兼性多能干細胞,目前普遍認為卵圓細胞位于小葉間膽管或Hering管,即終末膽管,也是門靜脈周圍肝細胞和膽管細胞的過渡區域。自噬(autophagy)是將細胞內受損、變性或者衰老的蛋白質以及受損細胞運輸到溶酶體進行消化降解的過程,是細胞應對營養缺乏、細胞密度負荷、低氧、氧化應激、感染等惡劣環境的生存方式之一。microRNAs(miRNAs)是一類新發現的在進化上高度保守的非編碼小RNAs,長約19-22nt,在轉錄后水平調控基因的表達,廣泛參與發育、器官形成、凋亡、增殖,甚至是腫瘤發生等重要生命過程。miR-21目前研究最多的miRNAs之一,它定位于人類第17q染色體區域,通過誘導S期細胞周期調節,在肝再生的增殖期上調,顯著促進細胞增殖。miRNAs通過調控自噬相關基因的表達,從而發揮對自噬的調節作用,miRNAs在調控自噬的發生過程中,大多為抑制自噬發生,也有少部分miRNAs能激活自噬。LC3是檢測自噬現象的特異性標記分子,LC3-II/LC3-I反映自噬程度。
1 材料和方法
1.1材料
大鼠肝卵圓細胞(HOC)購自中科院上海細胞庫;丹(磺)酰戊二胺(Monodansylcadaverine,MDC)、微管相關蛋白1輕鏈3β(LC3B)蛋白多克隆抗體、兔抗鼠GAPDH單克隆抗體購自美國Sigma公司;DMSO(北京索萊寶公司);CCK-8(日本同仁化學公司);胎牛血清、1640培養液、PBS(GIBCO公司);Hoechst33258染色液、BCA法蛋白定量試劑盒(上海碧云天生物技術研究所);密封培養罐、AnaeroPack(安寧包)產氣袋和氧氣指示劑(日本三菱瓦斯化學株式會社);RNA提取試劑Trizol購自美國invitrogen公司;逆轉錄試劑盒、SYBR Green Realtime PCR Master Mix-Plus-購自日本Toyobo公司;GAPDH購自中國杭州賢至生物科技有限公司;miR-21增強慢病毒載體、miR-21-inhibition慢病毒載體、miR-21慢病毒空載體由上海吉凱基因化學技術有限公司構建;Marker、ECL發光液購自美國Thermo公司;7500型實時熒光定量PCR儀購自美國ABI公司;ChemiDoc XRS儀器購自BIO-RAD公司。
1.2方法
1.2.1 細胞培養:大鼠肝卵圓細胞接種于含1%青鏈霉素、10%胎牛血清的1640培養液中,置于CO2培養箱中培養,細胞貼壁生長至對數生長期后,細胞計數后置入缺血缺氧微環境中進行試驗。
1.2.2 慢病毒轉染:以4×104個/mL HOC接種到96孔板中,等到細胞融合度達到30%進行慢病毒載體轉染,選用復感染指數(MOI值)為100,每個孔100 μL:慢病毒載體10 μL,慢病毒感染增強液80 μL,Polybrene液(50 μg/mL)10 μL。混勻后繼續培養,12 h后觀察細胞狀態,更換新鮮培養基,繼續培養。
1.2.3 慢病毒轉染HOC后穩定細胞株的篩選:將待篩選細胞接種于48孔板中至70%~80%融合度,待細胞貼壁后,加入嘌呤霉素(0,1,2.5,5,10 μg/mL)進行篩選。得出藥物處理48 h后細胞全部死亡的最低藥物濃度是5 μg/mL,將細胞種植在24孔板中,貼壁后進行慢病毒感染,感染第2天,加入嘌呤霉素藥物篩選,篩選后的細胞即時觀察熒光以確定獲得感染效率100%的細胞。
1.2.4 缺血缺氧模型:將慢病毒轉染好的肝卵圓細胞接種于培養皿,當細胞密度達到80%~90%時,換成無血清1640培養基,放入密封培養罐后迅速放入安寧包產氣袋一袋和氧氣指示劑一枚,密封,45 min后開始計時。
1.2.5 RNA的提取和逆轉錄:取三組長滿細胞的六厘米培養皿,同時每組加入1 mL Trizol,在冰上中靜置5 min后,用槍頭吹打細胞后移至離心管中,靜置5 min,按照invetrogen公司Trizol試劑說明書提取RNA。以所提總RNA為模版,分別進行RT反應,反應體系如下:總RNA 1 μL加入Enzyme Mix 0.5 μL、Buffer 2 μL、逆轉錄U6引物1 μL、miR-21莖環引物1 μL,補充Nuclease-free Water至10 μL。反應條件:37 ℃ 15 min,98 ℃ 5 min。
1.2.6 實時熒光定量PCR檢測miR-21表達:根據miRBase數據庫(http://www.mirbase.org/)獲得成熟miRNA序列,并用Primer5軟件設計相應的逆轉錄和PCR擴增引物,由上海生工生物股份有限公司合成。miR-21逆轉錄莖環引物5'-GTCGTATCCAGTGCA GGGTCCGAGGTATTCGCACTGGATACGACTCAACA-3',實時熒光定量PCR前向引物為5'-GCGGCGGTAGCTTATCAGAC TG-3',反向引物為莖環引物上的一段片段,為5'-ATC CAGTGCAGGGTCCGAGG-3'。內參U6逆轉錄引物為:5'-CG CTTCACGAATTTGCGTGTCA-3',實時熒光定量U6引物為Fp-5'-GCTTCGGCAGCACATATACTAAAAT-3',Rp-5'-CG CTTCACGAATTTGCGTGTCAT-3'。所得逆轉錄產物稀釋10倍后行熒光定量PCR,反應體系為:模版1 μL,SYBRqPCR Mix液5 μL,miR-21引物和負鏈引物各1 μL(濃度均為1 μmol/L),Plus solution 1 μL,補充水至10 μL,以U6為內參。反應條件為:95 ℃ 1 min預變性,95 ℃ 15 s、60 ℃ 15 s和72 ℃ 45 s,40個循環,采用2-△△CT法計算不同組織中miR-21的相對表達量。
1.2.7 MDC熒光染色:取細胞爬片,移去培養液,PBS洗2次,重新加入含有50 μmol/LMDC的培養基在37 ℃孵育1 h后,棄培養基,PBS洗三遍,抗熒光淬滅劑于載玻片上封片,置于DM4000 B LED熒光正置顯微鏡上,用380 nm波長的激發光525 nm波長的發射光濾光片進行觀察。
1.2.8 Hoechst 33258染色:取細胞爬片,棄培養液,4%多聚甲醛固定15 min,PBS洗兩次,Hoechst 33258染色5 min,PBS洗兩次,滴加抗熒光淬滅劑于載玻片上封片,熒光顯微鏡觀察,并隨機選取視野拍照。1.2.9 Western blotting:分別提取各組細胞蛋白,BCA法測定蛋白濃度,選擇10% SDS-PAGE凝膠,30 μg上樣量進行電泳,80 V恒壓至樣品進入分離膠后改120 V恒壓至電泳結束,350 mA恒流60 min轉膜后,用含5%脫脂奶粉TBST常溫封閉90 min,4 ℃搖床孵育一抗過夜(抗體稀釋濃度為LC3B為1:1 000,GAPDH 為1:500)。TBST洗膜三次后,以1:5 000濃度加入二抗至牛奶,室溫孵育1 h后,TBST洗三次,加入ECL,ChemiDoc XRS儀器曝光,并采用Image Lab對其進行分析。
1.3統計學分析

圖1 細胞轉染圖片(×100)
2 結果
2.1慢病毒轉染情況
由圖1可見慢病毒轉染效率達95%以上。
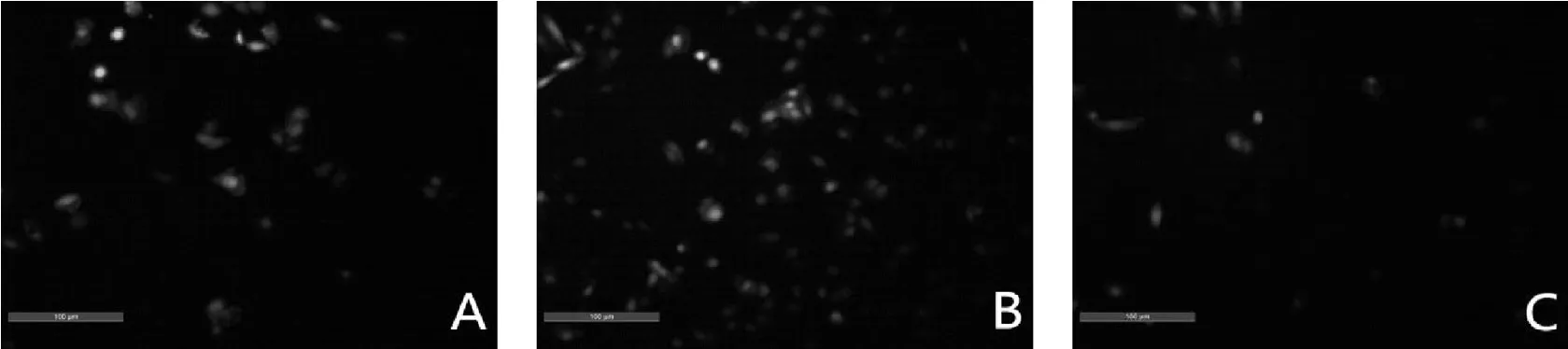
圖2 Hoechst 33258染色圖片(×100)

圖3 MDC染色圖片(×100)
2.2Hoechst 33258染色檢測細胞凋亡情況
相對于空載體對照細胞,miR-21增強慢病毒轉染的細胞染色增多(凋亡增多),miR-21-inhibition慢病毒轉染的細胞染色減少(凋亡減少),見圖2。
2.3MDC染色檢測細胞自噬
相對于空載體對照細胞,miR-21增強慢病毒轉染的細胞染色減少(自噬減少),miR-21-inhibition慢病毒轉染的細胞染色增多(自噬增多),見圖3。
2.4實時熒光定量PCR檢測miR-21
采用2-△△CT法計算出不同標本中miR-21的相對表達量,如圖4所示。
2.5Western blotting檢測LC3-II和LC3-I蛋白
Western blotting觀察到,相對于空載體對照細胞,miR-21增強慢病毒轉染的細胞LC3-II與LC3-I的比值降低,miR-21-inhibition慢病毒轉染的細胞LC3-II與LC3-I的比值增加(圖5)。
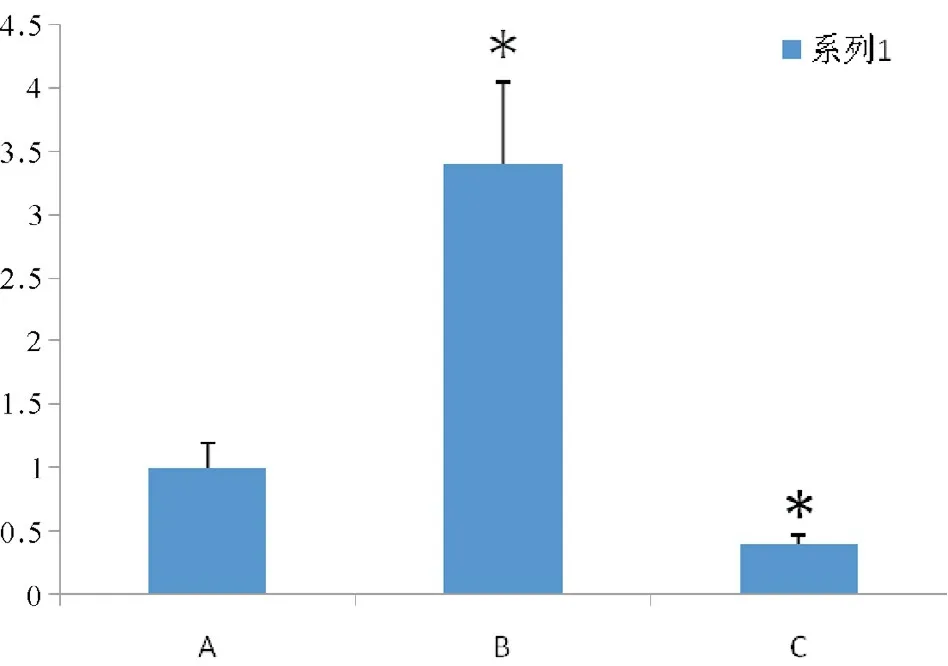
圖4 miR-21基因表達情況圖片
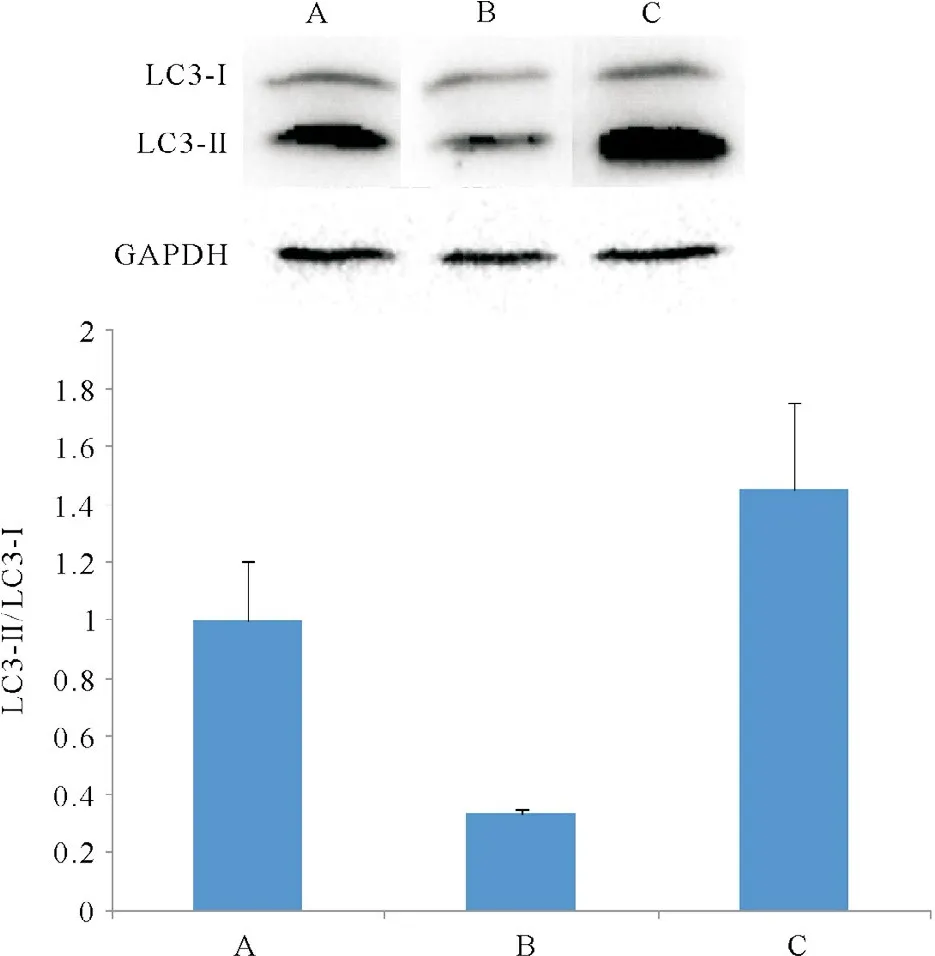
圖5 Western blotting檢測LC3蛋白圖片
3 討論
當肝臟受到嚴重損傷,肝細胞大量缺失或者肝細胞增殖受到毒素或者致癌因子的抑制時,就會激活肝卵圓細胞。肝卵圓細胞同時具有膽管細胞與肝細胞的表型,且可進一步分化為肝細胞及膽管上皮細胞,因而認為其是一種原始的兼性多功能干細胞[1]。目前普遍認為卵圓細胞位于小葉間膽管或Hering管,即終末膽管,也是門靜脈周圍肝細胞和膽管細胞的過渡區域。
自噬(autophagy)是一種進化保守的溶酶體依賴的自身降解途徑,是細胞在遇到應激反應等刺激時,將細胞質內多余的蛋白質或損傷的細胞器等物質包繞至自噬體中并與溶酶體結合將其降解的過程。自噬幾乎存在于所有真核細胞,不僅參與維持細胞內環境穩定及生長、分化的重要過程,而且是細胞面臨饑餓、生長因子匱乏等應激環境的應對策略[2]。根據其生理功能和自噬體與溶酶體的結合方式,將自噬分為3種類型:巨自噬(macroautophagy)、微自噬(microautophagy)和分子伴侶介導的自噬(chaperonmediated autophage),其中以巨自噬的研究最為成熟[3-4]。段文彪等[5]研究得出缺血缺氧可以抑制肝卵圓細胞增殖,并且可誘導肝卵圓細胞自噬。自噬有利于肝卵圓細胞在缺血缺氧微環境中穩定細胞內環境,維持細胞的存活。微管相關蛋白1輕鏈3(LC3)蛋白有兩種形式:LC3-I和LC3-II,細胞自噬體形成過程中,LC3-I可以轉化為LC3-II。LC3-II含量的多少與自噬體的含量有關,自噬體膜的含量由LC3-II標記[6],在自噬形成中,LC3-I與LC3-II/LC3-I比例與自噬體數量有關[7]。LC3-II/LC3-I的比值與自噬呈正相關。
microRNAs(miRNAs)是一類新發現的在進化上高度保守的非編碼小RNAs,長約19~22 nt在轉錄后水平調控基因的表達[8]。miR-21參與發育、再生及其干細胞生物學中有發揮重要作用,Marquez等[9]切除小鼠的部分肝臟,發現和肝切除術前相比,切除術后小鼠肝臟miR-21的表達在12、24 h達到高峰(超過正常2倍),提示miR-21在肝再生早期可能參與基因表達調控。陳楨坤等[10]證實了miR-21在肝卵圓細胞活化與增殖中起重要作用,Smad7作為miR-21的靶基因可能參與這一過程。
miRNA也被發現在自噬調節中扮演了關鍵的角色,在自噬的啟動(induction)、囊泡成核(vesicle nucleation)、囊泡延伸(vesicle elongation)、自噬回收(retrieval)與囊泡融合(fusion)等幾個階段中均有miRNA參與調控[11]。研究表明miRNAs能調節一些Atgs基因,而且調控子存在于自噬過程的不同階段[12]。Zhu等[13]首次揭示了miRNA與細胞自噬之間的聯系:miR-30a負調節自噬關鍵蛋白Beclin 1表達,引起自噬水平下降。miRNAs在調控自噬的發生過程中,大多為抑制自噬發生,也有少部分miRNAs能激活自噬。抑制自噬發生的相關miRNAs如miR-375在人類肝癌細胞中表達下調,轉染miR-375后能下調自噬相關基因Atg7信使RNA及蛋白表達水平,Atg7參與LC3I向LC3II的轉變所必需的Atg10和Atg12、Atg3和LC3復合物形成過程,當Atg7下調時自噬溶酶體的形成受阻,阻斷了自噬的發生,從而避免腫瘤治療中自噬引起的耐藥[14]。miR-100[15]、miR-18a[16]、miR-7[17]等都促進自噬的發生。
本實驗成功構建了肝卵圓細胞miR-21慢病毒轉染模型,Hoechst 33258染色miR-21增強慢病毒轉染的細胞染色增多(凋亡增多),miR-21-inhibition慢病毒轉染的細胞染色減少(凋亡減少),MDC染色miR-21增強慢病毒轉染的細胞染色減少(自噬減少),miR-21-inhibition慢病毒轉染的細胞染色增多(自噬增多)。miR-21過表達在本文中證實抑制HOC的自噬,抑制miR-21過表達可以增強缺血缺氧引起的肝卵圓細胞的自噬,有利于肝卵圓細胞在缺血缺氧微環境中穩定細胞內環境,維持細胞的存活,具體可能機制有待進一步研究。Lu等[18]研究miR-21的過表達能上調PTEN表達,抑制P13K-Akt-mTOR信號通路異常活化,抑制自噬。我們以miRNA為突破點研究肝卵圓細胞的自噬,為研究自噬提供一種新的思路。
參考文獻:
[1]FELDMANN G.Liver transplantation of hepatic stem cells:potential use for treating liver diseases[J].Cell Biol Toxicol, 2001, 17(2):77-85.
[2]TSUCHIHARA K, FUJII S, ESUMI H.Autophagy and cancer:dynamism of the metabolism of tumor cells and tissues[J].Cancer Lett, 2009, 278(2):130-138.
[3]MEHRPOUR M, ESCLATINE A, BEAU I, et al.Autophagy in health and disease.1.Regulation and significance of autophagy:an overview[J].AM J physid Cell Physiol, 2010, 298(4):C776-785.
[4]TANIDA I.Autophagosome formation and molecular mechanism of autophagy[J].Antioxid Redox Signal, 2011, 14(11):2201-2214.
[5]段文彪, 高勝強, 周春光, 等.自噬對肝卵圓細胞在缺血缺氧微環境中增殖的影響[J].肝膽胰外科雜志, 2015, 27(1):37-41.
[6]KABEYA Y, MIZUSHIMA N, UENO T, et al.LC3, a mammalian homologue of yeast Apg8p, is localized in autophagosome membranes after processing[J].EMBO J, 2000, 19(21):5720-5728.
[7]REN J H, HE W S, NONG L, et al.Acquired cisplatin resistance in human lung adenocarcinoma cells is associated with enhanced autophagy[J].Cancer Biothe Radiopharm, 2010, 25(1):75-80.
[8]BARTEL D P.MicroRNAs:genomics, biogenesis, mechanism, and function[J].Cell, 2004, 116(2):281-297.
[9]MARQUEZ R T, WENDLANDT E, GALLE C S, et al.MicroRNA-21 is upregulated during the proliferative phase of liver regeneration, targets Pellino-1, and inhibits NF-kappaB signaling[J].Am J Physiol Gastrointest Liver physiol, 2010, 298(4):G535-541.
[10]陳楨坤, 單云峰, 何彬, 等.miR-21在大鼠肝卵圓細胞活化和增殖過程中作用研究[J].中華肝臟病雜志, 2014, 22(11):854-859.
[11]HE C, KLIONSKY D J.Regulation mechanisms and signaling pathways of autophagy[J].Annu Rev genet, 2009, 43:67-93.
[12]XIAO J, ZHU X, HE B, et al.MiR-204 regulates cardiomyocyte autophagy induced by ischemia-reperfusion through LC3-II[J].J Biomed Sci, 2011, 18:35.
[13]ZHU H, WU H, LIU X, et al.Regulation of autophagy by a beclin 1-targeted microRNA, miR-30a, in cancer cells[J].Autophagy, 2009, 5(6):816-823.
[14]CHANG Y, YAN W, HE X, et al.miR-375 inhibits autophagy and reduces viability of hepatocellular carcinoma cells under hypoxic conditions[J].Gastroenterol, 2012, 143(1):177-187 e8.
[15]GE Y Y, SHI Q, ZHENG Z Y, et al.MicroRNA-100 promotes the autophagy of hepatocellular carcinoma cells by inhibiting the expression of mTOR and IGF-1R[J].Oncotarget, 2014, 5 (15):6218-6228.
[16]QASED A B, YI H, LIANG N, et al.MicroRNA-18a upregulates autophagy and ataxia telangiectasia mutated gene expression in HCT116 colon cancer cells[J].Mol Med Rep, 2013, 7 (2):559-564.
[17]TAZAWA H, YANO S, YOSHIDA R, et al.Genetically engineered oncolytic adenovirus induces autophagic cell death through an E2F1-microRNA-7-epidermal growth factor receptor axis[J].Int J Cancer, 2012, 131(12):2939-2950.
[18]LU X, FAN Q, XU L, et al.Ursolic acid attenuates diabetic mesangial cell injury through the up-regulation of autophagy via miRNA-21/PTEN/Akt/mTOR suppression[J].PloS one, 2015, 10(2):e0117400.
(本文編輯:張和,魯翠濤)
·論著 基礎研究·
[通訊作者簡介]單云峰,主任醫師,副教授,博士,碩士生導師,E-mail:shanyfeng@126.com。
作者簡介][第一高勝強(1990-),男,安徽馬鞍山人,在讀碩士。
[基金項目]浙江省自然科學基金項目(LY12H03006);溫州市科委課題(Y20110078)。
[收稿日期]2015-08-17
[中圖分類號]R329.2+6
[文獻標識碼]A
DOI:10.11952/j.issn.1007-1954.2016.01.011
