長鏈非編碼GATS表達干擾質(zhì)粒構建抑制乳腺癌細胞增殖和侵襲的研究
2016-05-25 06:58:14易剪梅錢立元陳學東
中國現(xiàn)代醫(yī)學雜志 2016年8期
易剪梅,錢立元,陳學東
(中南大學湘雅三醫(yī)院乳疝外科,湖南長沙410018)
?
長鏈非編碼GATS表達干擾質(zhì)粒構建抑制乳腺癌細胞增殖和侵襲的研究
易剪梅,錢立元,陳學東
(中南大學湘雅三醫(yī)院乳疝外科,湖南長沙410018)
摘要:目的探討長鏈非編碼GATS對乳腺癌細胞增殖和侵襲的影響。方法構建的人LncRNA-GATS-shRNA1~4慢病毒載體轉染乳腺癌MDA-MB-231細胞后,qPCR篩選出干擾效率為50.0%和54.0%的sh1組和sh3組,MTT法和Transwell實驗分別檢測轉染后細胞增殖和侵襲能力;流式細胞術檢測轉染sh1組慢病毒后細胞周期分布及凋亡變化。結果測序證實,LncRNA-GATS的shRNA寡聚核苷酸序列已被克隆到pLV-GATS載體;轉染MDA-MB-231細胞后,與陰性對照組比較,sh1組和sh3組的乳腺癌細胞增殖和侵襲能力均降低(P<0.01),以sh1組更顯著;sh1組細胞周期被阻滯于S期(P<0.01);細胞凋亡率無明顯變化(P> 0.05)。結論LncRNA-GATS可能下調(diào)乳腺癌細胞侵襲力,并通過抑制細胞周期進展降低細胞增殖能力。
關鍵詞:人LncRNA-GATS;shRNA干擾載體;MDA-MB-231細胞;增殖;侵襲
乳腺癌是婦女發(fā)病率最高的惡性腫瘤,其發(fā)病率占全身各種惡性腫瘤的7%~10%,近年來發(fā)病率不斷上升[1-3],其中三陰乳腺癌(triple negative breast cancer,TNBC)占20%[4]。三陰乳腺癌作為一種特殊的高危乳腺癌亞型,其侵襲性強、預后差、青年女性好發(fā),尋找其有效治療靶點迫在眉睫。長鏈非編碼RNA(long non-coding RNA,lncRNA)是一類長度> 200 nt的基因轉錄產(chǎn)物,占ncRNA的80%,既參與表觀遺傳、可變剪接、入核轉運等過程,也能以細胞微結構原件、小RNA前體等發(fā)揮功能[5]。近年,大量研究證實lncRNA在白血病、前列腺癌、乳腺癌、結腸癌、神經(jīng)膠質(zhì)瘤、肺癌等多種腫瘤中異常表達[6-10]。課題組前期基因芯片篩選乳腺浸潤性導管癌中18條差異表達人lncRNA(P<0.01),其中GATS較正常組織明顯下調(diào)[11]。本試驗構建GATS-lncRNA-shRNA慢病毒載體,轉染人乳腺癌細胞MDA-MB-231,檢測其對乳腺癌細胞增殖、侵襲的影響。
1 材料與方法
1.1材料
1.1.1細胞、菌株與質(zhì)粒人乳腺癌細胞MDA-MB-231、HEK-293FT脂質(zhì)體轉染細胞、pLV-GATS-shRNA慢病毒載體及慢病毒包裝系統(tǒng)、慢病毒對照LV-shNC等均由長沙贏潤生物技術有限公司提供。
1.1.2主要儀器及試劑熒光定量PCR儀(BioRad公司)、總RNA提取試劑Trizo(l美國Invitrogen公司),Transwell小室(美國Corning costar公司)、流式細胞儀(美國BD公司),Annexin V-APC/PI雙染細胞凋亡檢測試劑盒(南京凱基生物公司),DMEM培養(yǎng)基(Gibco公司),胎牛血清(杭州四季青公司),THUNDERBIRD SYB R?qPCR Mix(日本TOYOBO公司)、dNTP、RevertAidTMReverse Transcriptase、RiboLockTMRNase Inhibitor、Taq DNA Polymerase(MBI公司)、培養(yǎng)基:RPMI 1640(美國Invitrogen公司)+10% FBS(美國Gibco公司)+1% p/s(雙抗誘液,碧云天公司)、胰酶液(美國Gibco公司)。
1.2方法
1.2.1細胞培養(yǎng)MDA-MB-231細胞用含10%胎牛血清的DMEM培養(yǎng)基,在37℃飽和濕度、含5%二氧化碳CO2的孵箱中培養(yǎng)。
1.2.2構建慢病毒載體及細胞感染針對人GATS-lncRNA序列(NCBI Gen Bank,基因編號:NR028038),利用YRgene shRNA庫設計干擾序列(見表1),構建慢病毒重組載體pLV-GATS-lncRNA-shRNA1~4。取重組載體轉化至感受態(tài)細胞DH5α,同時設計空載體作為對照,挑選陽性克隆測序。將重組載體和包裝質(zhì)粒共轉染HEK293FT細胞,包裝慢病毒,4 d后用熒光顯微鏡技術觀察熒光表達,計數(shù)病毒滴度。包裝好的GATS-lncRNA-shRNA 1~4慢病毒和陰性對照組(NC-shRNA),以MOI值50轉染MDA-MB-231細胞。

表1 GATS的lncRNA序列及人工合成的shRNA oligo序列
1.2.3逆轉錄(RT-PCR)檢測lncRNA GATS表達按Trizol裂解液說明書進行細胞總RNA的抽提;按逆轉錄試劑盒說明進行逆轉錄。GAPDH為內(nèi)參基因,GATS lncRNA的正向引物為5'-TAAGCAGAGGGCTACAGAA-3',反向引物5'-CAGGGAGATC AGGAGGAC-3';GAPDH的正向引物為5'-TGCACCACCAACTGCTTAGC-3',反向引物5'-GGCATGGAC TGTGGTCATGAG-3',按熒光定量PCR試劑盒說明配置反應體系,熒光信號實時檢測,數(shù)據(jù)分析采用2-ΔΔCT(CT代表循環(huán)閾值)分析法。
1.2.4MTT法檢測轉染后MDA-MB-231細胞增殖篩選出干擾效率分別為50.0%和54.0%的sh1 和sh3組,取sh1、sh3及陰性對照組(NC組)3組MDA-MB-231細胞接種到96孔板,每孔接種8 000個細胞(100μl),每組設3個復孔。于慢病毒感染后48、72和96 h加入MTT溶液(20μl,5 mg/ml)。酶聯(lián)免疫監(jiān)測儀測定A490值。
1.2.5Transwell法檢測轉染后MDA-MB-231細胞侵襲轉染24 h后將60μl基質(zhì)膠稀釋于300μl無血清培養(yǎng)基中,取上述稀釋液100μl加入Transwell上室中,37℃培養(yǎng)箱中,孵育4~5 h。下室中加入600μl含有20%FBS的培養(yǎng)基。37℃培養(yǎng)箱孵育20~24 h,Transwell PBS洗2次,顯微鏡下計數(shù)5個高倍鏡視野細胞,計算平均數(shù)。每組設2個復孔,實驗重復3次。
1.2.6流式細胞儀檢測細胞周期分布各組細胞慢病毒感染72 h后,經(jīng)0.2%胰酶處理,離心收集細胞,用4℃預冷PBS洗滌2次,在離心管中留約0.5 ml PBS,加入預冷的70%乙醇1 ml混勻固定,4℃放置過夜;離心離去乙醇,PBS清洗1次,在離心管中留1 ml PBS,打散細胞團,加入Rnase 5μl(10 mg/ml),37℃放置1 h,400目篩網(wǎng)過濾,加入PI(100μg/ml)染液,室溫避光染色30 min,流式細胞儀(FACS)檢測細胞周期。實驗重復3次。
1.2.7流式細胞儀檢測細胞凋亡各組細胞慢病毒感染72 h后,經(jīng)0.25%胰酶處理,離心收集細胞,用4℃預冷PBS洗滌2次,然后用500μl結合緩沖液重懸細胞,調(diào)節(jié)其濃度為106/ml,然后取100μl細胞懸浮于5 ml流式管中,加入5μl Annexin V-APC混勻后,加入5μl PI混勻,于室溫避光孵育15 min,重懸于400μl PBS,400目篩網(wǎng)過濾,加樣于流式細胞儀(FACS)檢測細胞凋亡。激發(fā)波長Ex=488 nm,發(fā)射波長Em=530 nm。獲得細胞凋亡數(shù)據(jù)后用Mod Fit LT軟件進行細胞凋亡相對定量分析。實驗重復3次。
1.3統(tǒng)計學方法
采用SPSS 18.0統(tǒng)計軟件進行數(shù)據(jù)分析,計量資料用均數(shù)±標準差(±s)表示,各組間比較用單因素方差分析,兩兩比較用Dunnett法。方差不齊用非參數(shù)檢驗的Kruskal-Wallis H檢驗,兩兩比較用Dunnett's法,P<0.05為差異有統(tǒng)計意義。
2 結果
2.1重組人GATS-lncRNA-shRNA慢病毒載體測序和鑒定
經(jīng)測序分析,比對4種重組載體中插入的片段序列與設計的pLV-GATS-lncRNA-shRNA寡核苷酸序列完全一致。熒光顯微鏡下同一視野的熒光和可見光照片比較,可見大部分細胞發(fā)出熒光,病毒滴度分別為2×109TU/ml、6×108TU/ml、2×109TU/ml 和4×108TU/ml。
2.2RT-qPCR檢測GATS lncRNA表達
GATS-lncRNA-shRNA慢病毒轉染人乳腺癌細胞MDA-MB-231細胞,熒光顯微鏡證實轉染成功(見圖1)。RT-qPCR結果顯示,pLV-GATS-lncRNA-shRNA1~4慢病毒對GATS lncRNA干擾效率分別為50.0%、40.9%、54.0%和8.0%。
2.3MTT檢測轉染后MDA-MB-231細胞增殖能力變化
各組不同時相吸光度見表2,吸光度變化曲線見圖2。MTT檢測結果顯示,轉染后3組MDA-MB-231細胞的增殖水平在48、72和96 h差異有統(tǒng)計學意義(P<0.001);與對照組比較,sh1組的細胞增殖水平在48、72和96h差異有統(tǒng)計學意義(P=0.000),sh3組的細胞增殖水平除在72 h差異有統(tǒng)計學意義(P= 0.000),48和96 h差異無統(tǒng)計學意義(P=0.182和0.110)。
2.4Transwell法檢測轉染后MDA-MB-231細胞侵襲能力變化
非參數(shù)檢驗的Kruskal-Wallis H檢驗結果顯示,慢病毒轉染72 h后,3組的細胞侵襲力差異有統(tǒng)計學意義(P<0.01),兩兩比較結果顯示與NC組比較,sh1組和sh3組的細胞侵襲能力有所下降(P<0.01),但差異無統(tǒng)計學意義(P>0.05)。見圖3和表3。
2.5流式細胞儀分析細胞周期分布及凋亡
NC組細胞G1、S、G2期分別占(51.57±1.08)%、(28.04±1.23)%、(20.39±0.25)%,sh1組細胞G1、S、G2期分別占(37.60±1.00)%、(43.20±1.65)%、(19.20±0.66)%。與NC組比較,sh1組S期細胞比例明顯增高(P<0.01),說明轉染GATS-lncRNA-shRNA1后,MDA-MB-231細胞周期發(fā)生S期阻滯,見圖4。慢病毒轉染72 h后,3組的細胞凋亡率差異無統(tǒng)計學意義(P>0.05),不能認為轉染GATS-lncRNA-shRNA會影響MDA-MB-231細胞的凋亡率。見表4。
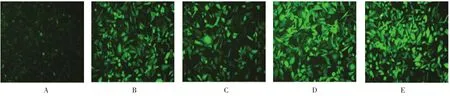
圖1 GATS-lncRNA-shRNA慢病毒感染MDA-MB-231細胞(×200)
表2 各組不同感染時相細胞吸光值比較(n=3,±s)

表2 各組不同感染時相細胞吸光值比較(n=3,±s)
注:?與NC組比較,P=0.000
組別 細胞增殖48 h 72 h 96 h NC組 0.430±0.006 0.533±0.008 0.617±0.008 sh1組 0.349±0.010? 0.422±0.012? 0.504±0.007?sh3組 0.416±0.012 0.464±0.009? 0.602±0.009 F值 61.408 97.783 178.359 P值 0.000 0.000 0.000
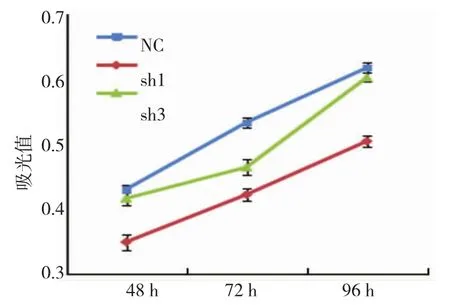
圖2 MTT檢測轉染GATS-lncRNA-shRNA對MDA-MB-231細胞增殖的影響

圖3 轉染GATS-lncRNA-shRNA對MDA-MB-231細胞侵襲力的影響
表3 各組穿膜細胞數(shù)比較(±s)
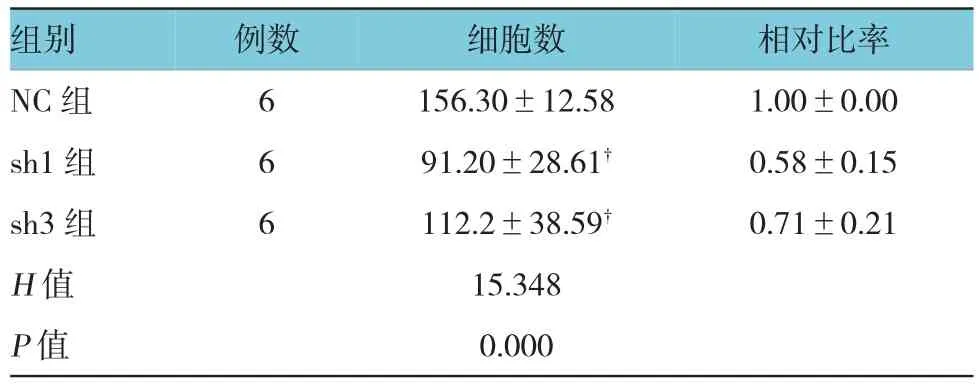
表3 各組穿膜細胞數(shù)比較(±s)
注:?與NC組比較,P<0.01
組別 例數(shù) 細胞數(shù) 相對比率NC組 6 156.30±12.58 1.00±0.00 sh1組 6 91.20±28.61? 0.58±0.15 sh3組 6 112.2±38.59? 0.71±0.21 H值 15.348 P值 0.000

圖4 轉染GATS-lncRNA-shRNA1對MDA-MB-231細胞周期的影響
表4 各組細胞凋亡率的比較(±s)

表4 各組細胞凋亡率的比較(±s)
組別 例數(shù) 凋亡率/% NC組 3 1.94±0.32 sh1組 3 1.37±0.27 sh3組 3 1.93±0.24 F值 P值4.178 0.073
3 討論
國內(nèi)外學者認為,乳腺癌MDA-MB-231細胞系能較好反應TNBC特性[12-13],故本研究選擇該細胞作為細胞模型。Jin等[14]研究發(fā)現(xiàn),MALAT1在TNBC組織中上調(diào),體外下調(diào)MALAT1可以抑制乳腺癌細胞增殖、遷移和增加癌細胞凋亡;在活體下調(diào)MALAT1抑制腫瘤生長和轉移,其很可能通過miR-1/SLUG軸而產(chǎn)生作用。同源異型框基因反義基因間RNAs(HOX antisense intergenenic RNAs,HOTAIR),通過ERS和ER共調(diào)節(jié)因子,HOTAIR反轉錄的雌二醇誘導下的轉錄失衡,導致乳腺癌變,并發(fā)現(xiàn)HOTAIR在轉錄水平被E2調(diào)節(jié),對于乳腺癌細胞的生存發(fā)育是必需的[15-16]。
人lncRNA-GATS為基質(zhì)抗原3(stromal antigen 3,STAG3)基因的反向鏈(opposite strand),又名STAG3OS,定位于7q22.1,外顯子計數(shù)為8,長度71 580 bp。STAG3含有C-末端結構域,一個28個氨基酸的保守的結構域和一個86個氨基酸的結構域。STAG3作為多亞基蛋白復合物的內(nèi)聚蛋白,在同源染色體分離的關鍵步驟中裝配/拆卸染色質(zhì),目前,確認其在偶線期復合物或著絲粒著絲點中的功能尚不明確,其在腫瘤中異常激活的相關機制待闡明[17-18]。
本實驗成功構建人GATS-lncRNA-shRNA慢病毒載體[19-20],轉染乳腺癌MDA-MB-231細胞,MTT實驗和Transwell實驗發(fā)現(xiàn),GATS-lncRNA-shRNA1 和GATS-lncRNA-shRNA3組增殖和侵襲能力較均降低,以sh1組更為顯著;流式分析發(fā)現(xiàn)轉染GATS-lncRNA-shRNA1后細胞周期進展被阻滯于S期,細胞凋亡率無明顯變化。該結果提示,GATS-lncRNA干擾后,細胞增殖受限可能并不是通過誘導凋亡來實現(xiàn),而是影響細胞周期來實現(xiàn)。
綜上所述,本研究首次探討lncRNA GATS在乳腺癌MDA-MB-231細胞增殖和侵襲過程中發(fā)揮的作用,提示GATS-lncRNA可能為乳腺癌治療的新的有效靶點。為進一步研究GATS-lncRNA與STAG3在乳腺癌發(fā)生、發(fā)展和轉移的過程中的相互關系奠定基礎。
參考文獻:
[1] Chao YL, Shepard CR, Wells A. Breast carcinoma cells re-express E-cadherin during mesenchymal to epithelial reverting transition[J]. Mol Cancer, 2010, 9(7): 179.
[2] Hou G, Zhang S, Zhang X, et al. Clinical pathological characteristics and prognostic analysis of 1013 breast cancer patients with diabetes[J]. Breast Cancer Res Treat, 2013, 137(3): 807-816.
[3] Hassan MS, Ansari J, Spooner D, et al. Chemotherapy for breast cancer (Review). Oncol Rep, 2010, 24(5): 1121-1131.
[4] Beg S, Siraj AK, Prabhakaran S, et al. Loss of PTEN expression is associated with aggressive behavior and poor prognosis in Middle Eastern triple-negative breast cancer [J]. Breast Cancer Res Treat, 2015, 151(3): 541-553.
[5] Di GF, Capaccioli S, Lulli M. A pathophysiological view of the long non-coding RNA world[J]. Oncotarget, 2014, 305(22): 10976-10996.
[6] Liu Y, Zhao J, Zhang W, et al. LncRNA GAS5 enhances G1cell cycle arrest via binding to YBX1 to regulate p21 expression in stomach cancer[J]. Sci Rep, 2015, 5(11): 10159.
[7] Kiren YU, Mark RP, Gwyn T. Williams. Reciprocal regulation of GAS5 lncRNA levels and mTOR inhibitor action in prostate cancer cells[J]. The Prostate, 2015, 75(7): 693-705.
[8] Dong S, Qu X, Li W, et al. The long non-coding RNA, GAS5, enhances gefitinib-induced cell death in innate EGFR tyrosine kinase inhibitor-resistant lung adenocarcinoma cells with wide-type EGFR via downregulation of the IGF-1R expression[J]. Hematol Oncol, 2015, 8: 43.
[9] Sun M, Liu XH, Wang KM, et al. Downregulation of BRAF activated non-coding RNA is associated with poor prognosis for non-small cell lung cancer and promotes metastasis by affecting epithelial-mesenchymal transition[J]. Mol Cancer, 2014, 13(1): 68.
[10] Sun T, Ye HH, Wu CL, et al. Emerging players in prostate cancer: long non-coding RNAs[J]. Am J Clin Exp Urol, 2014, 2(4): 294-299.
[11] Chen XD, Yang JY, Qian LY, et al. Aberrantly expressed mRNAs and long non-coding RNAs in patients with invasive ductal breast carcinoma: a pilot study[J]. Molecular medicine reports, 2015, 11(3): 2185-2190.
[12] Cailleau R, Olivé M, Cruciger QV. Long-term human breast carcinoma cell lines of metastatic origin: preliminary characterization[J]. In Vitro, 1978, 14(11): 911-915.
[13] Marc L, Guy L. Relevance of breast cancer cell lines as models for breast tumours: an update[J]. Breast Cancer Research and Treatment, 2004, 83(3): 249-289.
[14] Jin C, Yan BC, Qin L, et al. Reciprocal regulation of Hsa-miR -1 and long noncoding RNA MALAT1 promotes triple-negative breast cancer development[J]. Tumour Biol, 2015, (12): 1-12.
[15] Bhan A, Hussain I, Ansari K, et al. Antisense transcript long noncoding RNA(lncRNA) HOTAIR is transcriptionally induced by estradiol[J]. J Mol Biol, 2013, 425(19): 3707-3722.
[16] Bhan A, Mandal SS. Estradiol-Induced Transcriptional Regulation of Long Non-Coding RNA, HOTAIR [M]. 1st ed. Humanna Press, 2016: 395-412.
[17] Prieto I, Suja JA, Pezzi N, et al. Mammalian STAG3 is a cohesin specific to sister chromatid arms inmeiosis I[J]. Nat Cell Biol, 2001, 3(8): 761-766.
[18] Strunnikov A. Cohesin complexes with a potential to link mammalian meiosis to cancer[J]. Cell Regen (Lond), 2013, 2(1): 4.
[19] Heilbronn R, Weger S. Viral vectors for gene transfer: current status of gene therapeutics[J]. Handb Exp Pharmacol, 2010(197): 143-170.
[20]丁治國,何蓓,陳曉珩,等.人VEGF基因慢病毒介導RNA干擾有效靶點的設計及篩選[J].中國現(xiàn)代普通外科進展, 2013, 16(2): 85-88.
(張蕾編輯)
論著
Inhibitory regulation of human long non-coding RNA GATS on proliferation and invasion of breast cancer cells
Jian-mei Yi, Li-yuan Qian, Xue-dong Chen
(Department of Breast, Thyroid and Hernia Surgery, the 3rd Affiliated Hospital of Xiangya, Central South University, Changsha, Hunan 410018, China)
Abstract:Objective To investigate the effect of human long non-coding RNA GATS on proliferation and invasion of breast cancer MDA-MB-231 cells. Methods Four human GATS-lncRNA-specific oligonucleotide sequences were designed and synthesized previously, then were inserted into the pLV-GATS vector. After infecting the MDAMB-231 cells, transfection efficiencies of GATS-lncRNA-shRNA1 and GATS-lncRNA-shRNA3 were analyzed and screened by qPCR, which were 50.0% and 54.0% respectively. MTT and Transwell assays were used to detect the proliferation and invasion ability of the transfected cells, respectively. Flow cytometry was used to search the changes of cell cycle distribution and apoptosis of the MDA-MB-231 cells transfected GATS-lncRNA-shRNA1 lentiviral vector. Results Compared with the cells transfected with scrambled shRNA, the proliferation and invasion ability of cells the sh1 group and sh3 group was inhibited, especially t-hose the sh1 group. Flow cytometry showed the sh1 group cell cycle was arrested in S phase, and the cell apoptosis rate had no significant change (P > 0.05). Conclusions lncRNA GATS inhibits the invasion of breast cancer cells, and down regulates the proliferation of tumor cells through arresting cell cycle in S phase.
Keywords:Human lncRNA GATS; short hairpin RNA interference vector; MDA-MB-231 cells; proliferation; invasion
收稿日期:2016-01-27
文章編號:1005-8982(2016)08-0017-05
DOI:10.3969/j.issn.1005-8982.2016.08.004
中圖分類號:R655.8
文獻標識碼:A
猜你喜歡
中老年保健(2022年6期)2022-08-19 01:41:48
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2020年12期)2021-01-18 06:57:46
中學生數(shù)理化·七年級數(shù)學人教版(2020年12期)2021-01-18 06:57:46
中國生殖健康(2019年2期)2019-08-23 08:11:42
中國生殖健康(2019年6期)2019-01-06 09:20:12
中國生殖健康(2019年5期)2019-01-06 09:16:40
祝您健康(2018年5期)2018-05-16 17:10:16
