miR-222通過調控BCL2L13基因促進HBx-HepG2細胞生長*
2016-06-01 11:31:17余桂芳陳樹娣陳雪竹侯開連
中國病理生理雜志 2016年8期
余桂芳, 陳樹娣, 陳雪竹, 侯開連, 梁 敏
(廣州醫科大學附屬第五醫院腫瘤科,廣東 廣州 510700)
miR-222通過調控BCL2L13基因促進HBx-HepG2細胞生長*
余桂芳△, 陳樹娣, 陳雪竹, 侯開連, 梁 敏
(廣州醫科大學附屬第五醫院腫瘤科,廣東 廣州 510700)
目的: 探討HBx-HepG2 細胞中微小RNA-222(miR-222)對BCL2L13基因表達的調控作用以及對細胞生長和凋亡的影響,并研究其潛在的分子作用機制。方法: 利用實時熒光定量PCR檢測miR-222的表達水平;MTT和集落形成實驗檢測細胞的生長;流式細胞術檢測細胞周期和凋亡;構建BCL2L13 3’UTR雙螢光素酶報告載體,通過采用雙螢光素酶報告實驗驗證miR-222的靶基因。結果: 與正常的肝細胞 L02相比, miR-222在HBx-HepG2細胞過量表達(P<0.05)。miR-222過表達可促進HBx-HepG2細胞的生長,改變細胞周期,降低細胞的凋亡率;miR-222表達下調可抑制HBx-HepG2細胞的生長,改變細胞周期,增加細胞的凋亡率,和對照組相比差異有統計學顯著性(P<0.05)。與正常肝細胞 L02相比,BCL2L13在HBx-HepG2細胞表達下調(P<0.05);miR-222表達下調可促進BCL2L13的表達(P<0.05)。雙螢光素酶報告實驗和pcDNA3.1-BCL2L13轉染實驗結果提示miR-222可以通過作用于BCL2L13的3’UTR區,負向調控其表達,從而促進細胞的生長。結論: miR-222可以通過靶向調控BCL2L13基因進而促進HBx-HepG2細胞的生長。
微小RNA-222; HBx-HepG2細胞; 細胞生長; 細胞周期; 細胞凋亡;BCL2L13基因
微小RNA(microRNA,miRNA)是一種內源性的小分子單鏈RNA,長度介于22~25個堿基。miRNA能夠在轉錄后水平上對基因表達進行調控,其表達與多種人類腫瘤的發展有著密切的關系,miRNA既可作為癌基因促進腫瘤生長,又可作為抑癌基因抑制相關下游靶基因的表達[1-4]。研究發現miRNA-222(miR-222)在多種腫瘤上有著過表達;在肝癌組織中,miR-222 過量表達并且在腫瘤形成過程中具有促進肝癌細胞的生長、侵襲和轉移的能力[5-8]。目前miR-222在肝癌細胞中,特別是表達乙肝病毒X蛋白(hepatitis B virus X protein, HBx)的肝癌細胞HBx-HepG2中的作用機制尚不清楚,本研究通過生物信息學方法預測miR-222的靶基因,利用雙螢光素酶報告系統進行檢測,通過分子生物學技術證實了miR-222通過作用于BCL2L13基因對HBx-HepG2細胞生長產生影響,從而進一步解釋了miR-222影響HBx-HepG2生長的分子作用機制。
材 料 和 方 法
1 主要材料
HBx-HepG2細胞株是之前實驗構建的穩定表達HBx的HepG2細胞株[9]; L02 和 HEK293T細胞株購自上海中科院細胞庫;RPMI-1640培養液和胎牛血清購自HyClone; miR-222模擬物(miR-222 mimics)、miR-222抑制物(miR-222 inhibitor)、無關序列模擬物對照(miR-control,miR-Ctrl)、抑制物對照(miR inhibitor-Ctrl)以及miR-222的引物均購于廣州瑞博生物科技有限公司;High Pure miRNA Isolation Kit 購于Roche;miRNA-222 cDNA Synthesis Kit和Lipofectamine 2000購于Invitrogen;miR-222熒光定量PCR試劑盒購于TaKaRa;pmirGLO質粒和pcDNA3.1-BCL2L13質粒購于上海捷瑞生物工程有限公司;雙螢光素酶報告基因試劑盒購于Promega;MTT購于Sigma; Annexin V-FITC/PI凋亡試劑盒購于東仁化學科技有限公司。
2 主要方法
2.1 細胞培養 HBx-HepG2細胞,L02細胞用含有10%胎牛血清的RPMI-1640培養液,置于37 ℃、5% CO2的細胞培養箱內常規培養,消化傳代。當6孔板里每孔細胞數量達2×105細胞時,用不含雙抗的培養液培養24 h, 細胞融合率達80%時采用Lipofectamine 2000試劑進行轉染,在轉染miR-222的實驗中,實驗分為miR-222 mimics組和miR-222 inhibitor組,以及各自的對照組miR-Ctrl和 miR inhibitor-Ctrl;在轉染pcDNA3.1-BCL2L13實驗中,實驗分為pcDNA3.1空質粒組和pcDNA3.1-BCL2L13組。轉染后24 h收集細胞進行各項檢測。
2.2 實時熒光定量PCR(RT-qPCR)檢測細胞miR-222以及BCL2L13的表達 使用High Pure miRNA Isolation Kit以及TRIzol分別提取miRNA和mRNA, 然后將miRNA或者mRNA逆轉錄成cDNA; 采用熒光定量PCR 儀擴增檢測miR-222和BCL2L13目的片段。miR-222以U6為內參照,BCL2L13以GAPDH為內參照,采用2-ΔΔCt方法分析結果,每組設3個復孔,實驗重復3次。
2.3 MTT法檢測細胞活力 取對數生長期的HBx-HepG2細胞每孔1×104個細胞接種在96孔板中預培養24 h, 然后按照2.1分組轉染細胞,轉染24 h的HBx-HepG2細胞每孔加入MTT 20 μL 繼續培養4 h 后小心吸棄孔內培養上清液,每孔加入DMSO 150 μL,用酶聯免疫標記分析儀在570 nm 波長處測定各孔的吸光度A值,按下列公式計算細胞生長率(%)=實驗組A值/對照組A值×100%。為了檢測BCL2L13對細胞活力的影響,取對數生長期的HBx-HepG2細胞以每孔1×104接種在96孔板中預培養24 h, 然后按照2.1分組(pcDNA3.1組與pcDNA3.1-BCL2L13組)轉染細胞,對轉染24 h的HBx-HepG2細胞進行MTT檢測。
2.4 細胞平板克隆形成實驗 將轉染24 h 的HBx-HepG2細胞接種于6 孔培養板,每孔2 mL,充分混勻,常規培養10 d,每組3個復孔。每隔3 d更換新鮮培養液繼續培養。倒盡培養基,生理鹽水洗滌2遍,空氣干燥后75%乙醇固定15 min,0.5%結晶紫染色2 h,清水洗凈干燥。顯微鏡下觀察拍照,形成的克隆(≥50個細胞為1個克隆)計數。
2.5 流式細胞術檢測細胞凋亡 將轉染48 h的HBx-HepG2細胞用PBS洗滌,加入70% 冷乙醇固定后重新收集細胞,PBS洗去固定液,加入RNA 酶反應過夜,與碘化丙啶(propidium iodide,PI)染液混勻后,用流式細胞儀作流式細胞分析,激發波長為488 nm,并用ModFit LT 2.0 軟件分析細胞周期分布及凋亡情況。計算各期細胞所占比例。Annexin V-FITC/PI雙染后檢測細胞凋亡率。
2.6 雙螢光素酶報告基因系統驗證miR-222對BCL2L13 基因的調控作用 根據TargetScan靶基因預測數據庫預測miR-222的靶基因,我們發現BCL2L13為miR-222的潛在靶基因并尋找出與miR-222 互補配對的BCL2L13 3’UTR區。提取正常基因組,PCR擴增包含有miR-222結合位點的BCL2L13 3’UTR區,回收純化后插入到pmirGLO載體多克隆位點中,載體命名為pmirGLO-BCL2L13-wide type(BCL2L13-3’UTR-WT); 設計針對BCL2L13 3’UTR“種子區”的突變引物,插入到pmirGLO載體多克隆位點中,載體命名為pmirGLO-BCL2L13-mutant(BCL2L13-3’UTR-Mut)。螢光素酶報告載體BCL2L13-3’UTR-WT和BCL2L13-3’UTR-Mut分別與miR-222 mimics和miR-222 inhibitor,以及各自的對照(miR-Ctrl和miR inhibitor Ctrl)兩兩組合共轉染到HEK293T 細胞中。每組設3個復孔。轉染48 h后收集細胞按照螢光素酶報告基因檢測試劑盒說明書測定各組細胞螢火蟲螢光素信號及海腎螢光素信號。
3 統計學處理
采用SPSS 16.0軟件進行數據分析。所有數據以均數±標準差(mean±SD)表示,兩組間均數的比較采用Student’st檢驗,多組間均數的比較采用單因素方差分析(one-way ANOVA)。以P<0.05為差異有統計學意義。
結 果
1 HBx-HepG2細胞中miR-222的表達
RT-qPCR結果發現HBx-HepG2細胞中的miR-222表達顯著高于對照細胞株L02中miR-222的表達 (P<0.05),見圖1A。 miR-222 mimics轉染組HBx-HepG2細胞中的miR-222表達水平與miR-Ctrl相比顯著升高(P<0.05); 而miR-222 inhibitor轉染組HBx-HepG2細胞中的miR-222表達水平與miR inhibitor-Ctrl相比顯著降低(P<0.05),見圖1B。
2 miR-222對HBx-HepG2細胞生長、細胞周期以及凋亡的影響
MTT法檢測結果顯示,miR-222 mimics 轉染組HBx-HepG2細胞的生存率顯著高于miR-Ctrl組(P<0.05);而miR-222 inhibitor轉染組HBx-HepG2細胞的生存率顯著低于miR inhibitor-Ctrl組(P<0.05),見圖1C。細胞平板克隆形成實驗的結果可見,miR-222 mimics轉染組的HBx-HepG2集落形成數目顯著高于miR-Ctrl組(P<0.05);而miR-222 inhibitor轉染組的HBx-HepG2集落形成數目顯著低于miR inhibitor-Ctrl組(P<0.05),見圖1D。流式細胞術實驗結果發現,miR-222 mimics轉染組HBx-HepG2的G0/G1細胞比例下降和S期細胞比例上升(P<0.05)并且細胞凋亡率與miR-Ctrl組相比顯著降低(P<0.05);而miR-222 inhibitor轉染組HBx-HepG2的G0/G1細胞比例上升和S期細胞比例下降(P<0.05);細胞凋亡率與miR inhibitor-Ctrl組相比顯著升高(P<0.05),見圖2。
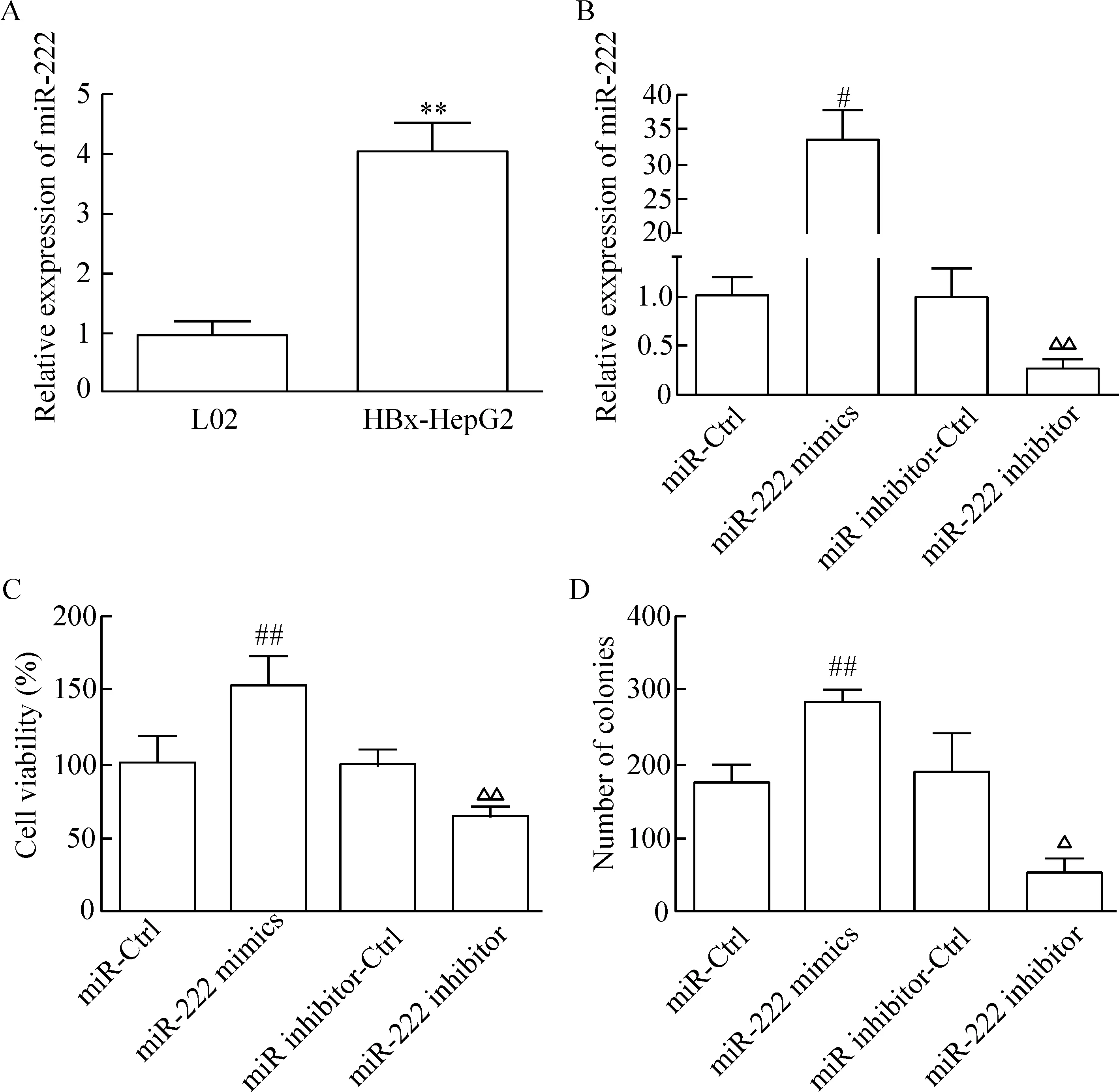
Figure 1.The effect of miR-222 on the growth of HBx-HepG2 cells. A: the expression of miR-222 in L02 and HBx-HepG2 cells; B: the effect of miR-222 mimics or miR-222 inhibitor transfection on miR-222 expression; C: the effect of miR-222 on the viability of HBx-HepG2 cells; D: the effect of miR-222 on colony formation of HBx-HepG2 cells. Mean±SD.n=3.**P<0.01vsL02;#P<0.05,##P<0.01vsmiR-Ctrl;△P<0.05,△△P<0.01vsmiR inhibitor-Ctrl group.
圖1 miR-222 對HBx-HepG2細胞生長的影響
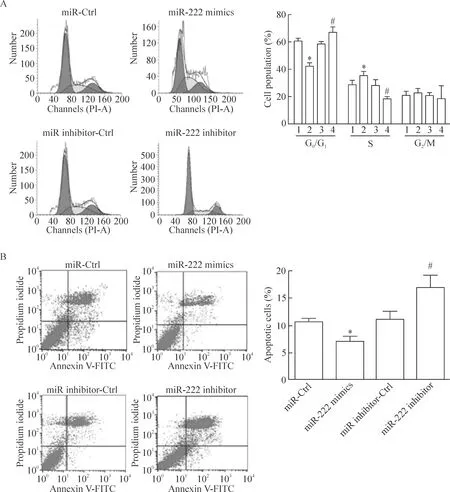
Figure 2.The effect of miR-222 on the cell cycle and apoptotic rate in the HBx-HepG2 cells. A: the effect of miR-222 mimics or miR-222 inhibitor transfection on the cell cycle; B: the effect of miR-222 mimics or miR-222 inhibitor transfection on the apoptotic rate. 1: miR-Ctrl; 2: miR-222 mimics; 3: miR inhibitor-Ctrl; 4: miR-222 inhibitor. Mean±SD.n=3.*P<0.05vsmiR-Ctrl;#P<0.05vsmiR inhibitor-Ctrl group. 圖2 miR-222對HBx-HepG2細胞周期以及凋亡率的影響
3 miR-222對BCL2L13 3’UTR表達的影響
采用雙螢光素酶報告基因檢測各組螢光信號的變化。實驗結果發現,miR-222 mimics和BCL2L13-3’ UTR-WT共轉染HEKT293細胞后,細胞的螢光素酶與對照組相比明顯受到抑制;miR-222 inhibitor和BCL2L13-3’ UTR-WT共轉染細胞后,細胞的螢光素酶與對照組相比明顯升高(P<0.05)。而miR-222 mimics或者miR-222 inhibitor和BCL2L13-3’ UTR-Mut共轉染細胞后, 螢光素酶的表達水平與對照組相比差異無統計學顯著性(P>0.05),見圖3。
4BCL12L13在HBx-HepG2中的表達及其對細胞生長的影響
RT-qPCR實驗結果發現,BCL12L13在HBx-HepG2細胞中的表達水平顯著低于正常肝細胞L02的表達水平(P<0.05),見圖4A。同時,在HBx-HepG2轉染miR-222 inhibitor后,BCL2L13的表達水平明顯高于只轉染miR inhibitor-Ctrl的細胞(P<0.05),見圖4B。轉染pcDNA3.1-BCL2L13載體能夠顯著提高HBx-HepG2細胞中BCL2L13的表達水平(P<0.05),見圖4C。同時轉染pcDNA3.1-BCL2L13載體的HBx-HepG2細胞生存率明顯低于轉染了空載體的HBx-HepG2細胞(P<0.05),見圖4D。
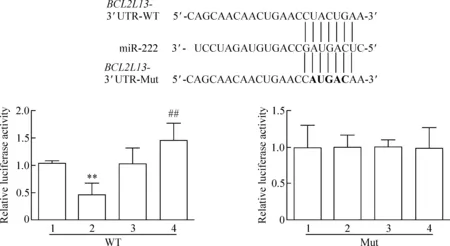
Figure 3. The targeted inhibition ofBCL2L13 expression in the HEKT293 cells by miR-222 detected by dual-luciferase reporter assay. 1: miR-Ctrl; 2: miR-222 mimics; 3: miR inhibitor-Ctrl; 4: miR-222 inhibitor. Mean±SD.n=3.**P<0.01vsmiR-Ctrl;##P<0.01vsmiR inhibitor-Ctrl.
圖3 雙螢光素酶報告系統檢測提示miR-222靶向抑制HEKT293細胞中BCL2L13基因表達
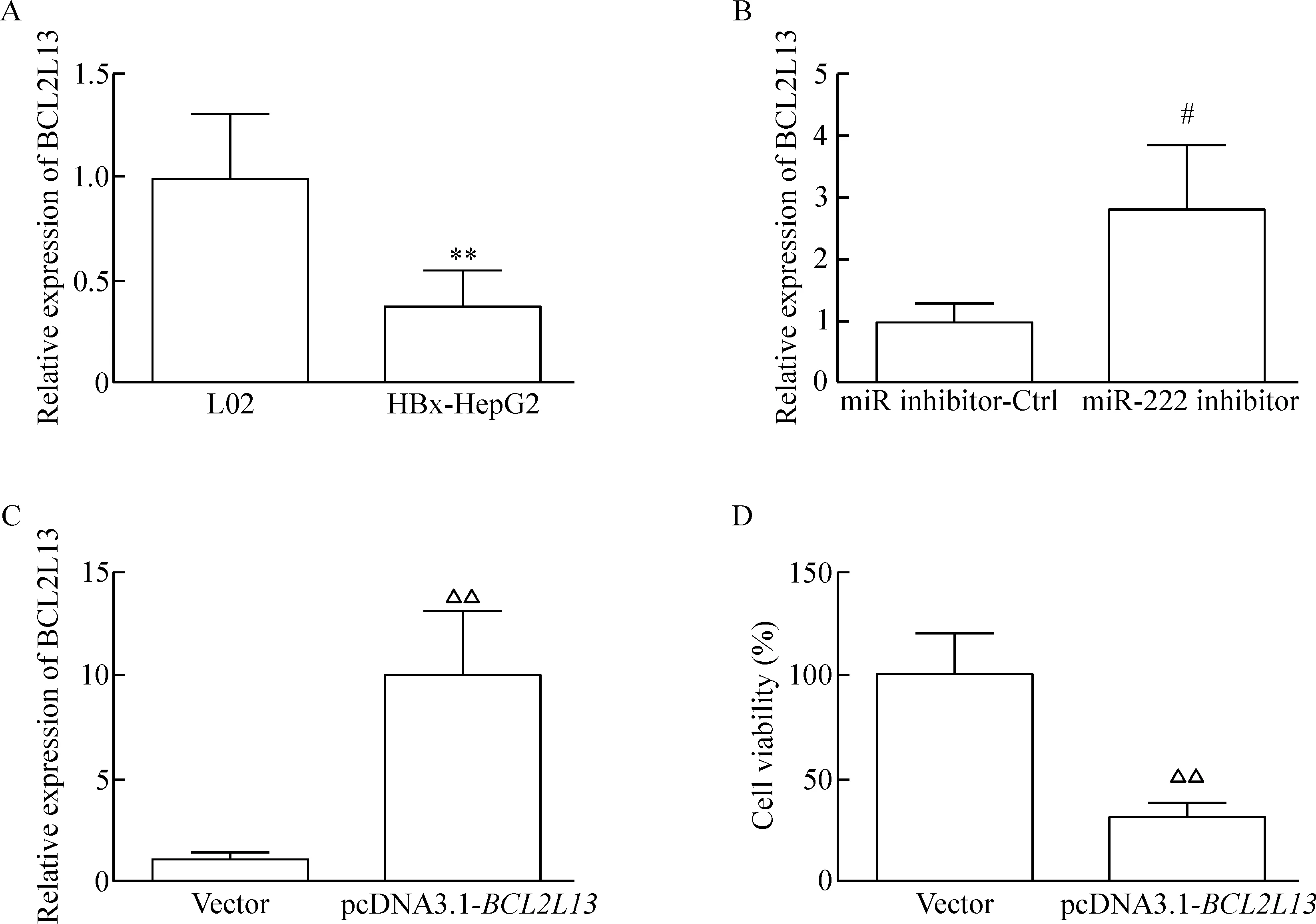
Figure 4.BCL2L13 was down-regulated in the HBx-HepG2 cells and inhibited cell growth. A: the expression ofBCL2L13 in the L02 and HBx-HepG2 cells; B: the effect of miR-222 inhibitor transfection on the expression level ofBCL2L13; C: the effect of pcDNA3.1-BCL2L13 transfection on the expression ofBCL2L13; D: the effect of pcDNA3.1-BCL2L13 transfection on the viability of HBx-HepG2 cells. Mean±SD.n=3.**P<0.01vsL02;#P<0.05vsmiR inhibitor-Ctrl;△△P<0.01vsvector.
圖4BCL2L13在HBx-HepG2細胞中下調并且可以抑制細胞生長
討 論
肝細胞癌(hepatocellular carcinoma,HCC)是最常見的惡行腫瘤之一,死亡率位于世界第4,慢性乙肝病毒感染是其發病的主要原因之一[10-11]。HBx是HBV 4個開放讀碼框中的X-ORF編碼的病毒蛋白,為含有154個氨基酸的多肽,分子量為17.5 kD,研究發現HBx促進了與其相關的HCC的發展[12-13]。HBx蛋白通過與不同宿主因素相互作用來調節細胞信號轉導,轉錄調控,細胞周期進程,DNA修復,細胞凋亡和遺傳穩定性[14]。前期的實驗中我們成功建立了穩定表達HBx的HCC細胞模型,并且發現穩定表達的HBx促進HepG2細胞的增殖,本實驗在前面基礎上進一步研究其分子機制,為尋找新的早期診斷標志和治療靶點提供依據。
miRNA既可作為癌基因促進腫瘤生長,又可作為抑癌基因抑制相關下游靶基因的表達[2]。研究發現miR-222在多種腫瘤上有著過表達;在肝癌組織中,miR-222 過量表達并且在腫瘤形成過程中具有促進肝癌細胞的生長、侵襲和轉移的能力[5-8]。進一步的研究發現miR-222的過表達與肝癌細胞的轉移有關,而且miR-222是通過AKT信號通路來影響肝癌細胞轉移的[15]。同時研究發現miR-222能夠通過抑制GNAI3的表達來促進肝癌細胞的轉移和入侵[16]。miR-222的過量表達能夠下調p27的表達從而來促進肝癌細胞的增長[5]。本研究第一次在HBx-HepG2細胞上證明miR-222的過表達可以促進細胞的增長,而miR-222影響HBx-HepG2細胞的增長可能是通過調控細胞周期,降低細胞的凋亡率而實現的。
我們通過生物信息學分析發現BCL2L13基因是miR-222的一個潛在靶點,雙螢光素酶報告實驗和pcDNA3.1-BCL2L13轉染實驗結果提示miR-222可以通過作用于BCL2L13的3’UTR區,負向調控其表達,從而促進細胞的生長。BCL2L13屬于Bcl-2家族中的一員,BCL2L13的過表達可以促進細胞的凋亡[17]。盡管BCL2L13基因與細胞的凋亡有關,但是其具體的分子機制還是有很多爭議。研究發現BCL2L13基因的過表達與小孩急性白血病的生存率呈負相關[18]。在多型性神經膠母細胞中,BCL2L13能夠抑制化療藥物引起的細胞凋亡[19]。而本實驗發現BLC2L13 的過表達可以抑制HBx-HepG2的生長。這些具有爭議的結果可能因為不同的細胞系或是不同的癌癥種類。BCL2L13基因在 HBx-HepG2細胞中的分子機制可能還需要進一步的研究。
綜上所述,miR-222可以通過靶向調控BCL2L13基因進而促進HBx-HepG2細胞的生長。為了進一步闡述miR-222在HBx-HepG2中的機制,我們將來的實驗有必要觀察miR-222在臨床樣本中的表達情況,從而進一步確認miR-222 在慢性乙肝病毒感染肝癌中的臨床意義。
[1] Cheng G. Circulating miRNAs: roles in cancer diagnosis, prognosis and therapy[J]. Adv Drug Deliv Rev, 2015, 81:75-93.
[2] Croce CM, Calin GA. miRNAs, cancer, and stem cell division[J]. Cell, 2005, 122(1):6-7.
[3] Thorgeirsson SS, Lee JS, Grisham JW. Functional geno-mics of hepatocellular carcinoma[J]. Hepatology, 2006, 43 (2 Suppl 1): S145-S150.
[4] 嵇曉輝,范秉琳,張紅鴿,等. MicroRNA-100對肝癌細胞增殖活力和細胞周期的影響[J]. 中國病理生理雜志, 2013, 29(1):108-111.
[5] Yang YF, Wang F, Xiao JJ, et al. miR-222 overexpression promotes proliferation of human hepatocellular carcinoma HepG2 cells by downregulating p27[J]. Int J Clin Exp Med, 2014, 7(4):893-902.
[6] Zhong C, Ding S, Xu Y, et al. MicroRNA-222 promotes human non-small cell lung cancer H460 growth by targeting p27[J]. Int J Clin Exp Med, 2015, 8(4): 5534-5540.
[7] Liu S, Sun X, Wang M, et al. A microRNA 221- and 222-mediated feedback loop maintains constitutive activation of NFκB and STAT3 in colorectal cancer cells[J]. Gastroenterology, 2014, 147(4):847-859.e11.
[8] Liu W, Song N, Yao H, et al. miR-221 and miR-222 simultaneously target RECK and regulate growth and invasion of gastric cancer cells[J]. Med Sci Monit, 2015, 21:2718-2725.
[9] 余桂芳, 嚴躍紅, 王瑞鑫,等. 慢病毒介導HBV X基因穩定表達HepG2細胞系的建立[J]. 世界華人消化雜志, 2012, 20(8):638-643.
[10]Motavaf M, Safari S, Saffari Jourshari M, et al. Hepatitis B virus-induced hepatocellular carcinoma: the role of the virus x protein[J]. Acta Virol, 2013, 57(4):389-396.
[11]Guerrieri F, Belloni L, Pediconi N, et al. Molecular mechanisms of HBV-associated hepatocarcinogenesis[J]. Semin Liver Dis, 2013, 33(2):147-156.
[12]Bouchard MJ, Schneider RJ. The enigmatic X gene of hepatitis B virus[J]. J Virol, 2004, 78(23):12725-12734.
[13] 崔利園,張釗瑞,費洪榮,等. 乙肝病毒X蛋白相互作用蛋白促進HepG2細胞遷移并調節β-catenin表達[J]. 中國病理生理雜志, 2012, 28(6): 1128-1131.
[14]Riviere L, Ducroux A, Buendia MA. The oncogenic role of hepatitis B virus[J]. Recent Results Cancer Res, 2014, 193:59-74.
[15]Wong QW, Ching AK, Chan AW, et al. miR-222 overexpression confers cell migratory advantages in hepatocellular carcinoma through enhancing AKT signaling[J]. Clin Cancer Res, 2010, 16(3):867-875.
[16]Zhang Y, Yao J, Huan L, et al. GNAI3 inhibits tumor cell migration and invasion and is post-transcriptionally regulated by miR-222 in hepatocellular carcinoma[J]. Cancer Lett, 2015, 356(2 Pt B):978-984.
[17]Kataoka T, Holler N, Micheau O, et al. Bcl-rambo, a novel Bcl-2 homologue that induces apoptosis via its unique C-terminal extension[J]. J Biol Chem, 2001, 276(22):19548-19554.
[18]Yang YL, Lin SR, Chen JS, et al. Expression and prognostic significance of the apoptotic genesBCL2L13,Livin, andCASP8AP2 in childhood acute lymphoblastic leukemia[J]. Leuk Res, 2010, 34(1):18-23.
[19]Jensen SA, Calvert AE, Volpert G, et al.BCL2L13 is a ceramide synthase inhibitor in glioblastoma[J]. Proc Natl Acad Sci U S A, 2014, 111(15): 5682-5687.
(責任編輯: 林白霜, 羅 森)
miR-222 enhances HBx-HepG2 cell growth via regulation ofBCL2L13 gene
YU Gui-fang, CHEN Shu-di, CHEN Xue-zhu, HOU Kai-lian, LIANG Min
(DepartmentofOncology,TheFifthAffiliatedHospitalofGuangzhouMedicalUniversity,Guangzhou510700,China.E-mail:guifangyu1023@sina.com)
AIM: To investigate the regulation of miR-222 onBCL2L13 gene and its effect on the growth and apoptosis of HBx-HepG2 cells, and to explore the underlying molecular mechanisms. METHODS: The expression level of miR-222 was detected by RT-qPCR. The HBx-HepG2 cell growth was examined by MTT and colony formation assays. The cell cycle and apoptosis were analyzed by flow cytometry. The recombination vector pmirGLO-BCL2L13 was constructed, and dual-luciferase reporter experiment was performed to validate the target of miR-222. RESULTS: The expression level of miR-222 in the HBx-HepG2 cells was significantly higher than that in the L02 cells (P<0.05). Over-expression of miR-222 enhanced HBx-HepG2 cell growth, changed cell cycle, and inhibited apoptosis (P<0.05). Knockdown of miR-222 reduced HBx-HepG2 cell growth, changed cell cycle, and increased cell apoptotic rate (P<0.05).BCL2L13 was down-regulated in the HBx-HepG2 cells as compared with L02 cells (P<0.05), and knockdown of miR-222 in the HBx-HepG2 cells increased the expression level ofBCL2L13 (P<0.05). The results of dual-luciferase reporter assay and restore experiment showed that miR-222 negatively regulated the expression ofBCL2L13 via targeting 3’UTR ofBCL2L13, resulting in the promotion of HBx-HepG2 cell growth. CONCLUSION: miR-222 enhances HBx-HepG2 cell growth via down-regulation ofBCL2L13.
MicroRNA-222; HBx-HepG2 cells; Cell growth; Cell cycle; Apoptosis;BCL2L13 gene
1000- 4718(2016)08- 1389- 06
2015- 11- 25
2016- 07- 06
廣東省科技計劃項目(No. 20130319c)
R363
A
10.3969/j.issn.1000- 4718.2016.08.008
雜志網址: http://www.cjpp.net
△通訊作者 Tel: 020-32208059; E-mail: guifangyu1023@sina.com
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小讀者(2021年2期)2021-03-29 05:03:48
華人時刊(2019年13期)2019-11-17 14:59:54
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
天津醫科大學學報(2019年3期)2019-08-13 06:53:08
文苑(2018年22期)2018-11-19 02:54:14
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
