內(nèi)生真菌對高羊茅種子耐干旱能力的影響
2016-06-06 06:13:36陳志豪劉學鋒鮑根生李曉霞紀劍強陳水紅
塔里木大學學報 2016年2期
陳志豪 劉學鋒 鮑根生 李曉霞 紀劍強 陳水紅*
(1 塔里木大學動物科學學院, 新疆 阿拉爾 843300)(2 新疆生產(chǎn)建設(shè)兵團第十四師一牧場, 新疆 策勒 848306)(3 青海省畜牧獸醫(yī)科學院, 青海 西寧 810016)(4 塔里木大學生命科學學院, 新疆 阿拉爾 843300)
?
內(nèi)生真菌對高羊茅種子耐干旱能力的影響
陳志豪1劉學鋒2鮑根生3李曉霞1紀劍強4陳水紅4*
(1 塔里木大學動物科學學院, 新疆 阿拉爾 843300)(2 新疆生產(chǎn)建設(shè)兵團第十四師一牧場, 新疆 策勒 848306)(3 青海省畜牧獸醫(yī)科學院, 青海 西寧 810016)(4 塔里木大學生命科學學院, 新疆 阿拉爾 843300)
摘要禾草內(nèi)生真菌廣泛的存在于禾本科植物體內(nèi),他們不會引起植物任何感病癥狀,內(nèi)生真菌侵染對植株生長有促進作用、且具有提高植物抵抗生物和非生物脅迫能力。本研究以感染內(nèi)生真菌和非感染內(nèi)生真菌高羊茅種子為研究對象,研究不同濃度PEG6000脅迫處理下,帶菌和不帶菌種子萌發(fā)能力差異。結(jié)果表明:在高羊茅種子萌發(fā)時,用PEG-6000模擬干旱脅迫種子,感染內(nèi)生真菌高羊茅種子的發(fā)芽率和胚根長均高于非感染種子的,說明內(nèi)生真菌浸染能夠提高高羊茅種子萌發(fā)時的耐旱能力。帶菌高羊茅將會更好的適應(yīng)干旱環(huán)境。
關(guān)鍵詞內(nèi)生真菌; 高羊茅; 抗旱性; 發(fā)芽率
干旱化趨勢已成為全球關(guān)注的焦點問題,有關(guān)植物對干旱生境的適應(yīng)機理及抗旱本質(zhì)等研究已成為植物生理學研究的重要課題之一。內(nèi)生真菌是一類在植物體內(nèi)渡過大部分或全部生命周期,而寄主植物不顯示外部癥狀的一類真菌,菌絲生長于植物組織的細胞間,其中葉鞘和種子中菌絲含量最多[1]。至今已在許多高等植物中發(fā)現(xiàn)內(nèi)生真菌的存在。與植物共生的真菌按照共生部位的不同可分為兩類: 一類是存在于植物根部的菌根真菌; 另一類是存在于健康植物莖葉中的內(nèi)生真菌[2]。在自然情況下,禾草內(nèi)生真菌分別與高羊茅(Festuca arundanacea)和多年生黑麥草(Lolaum perenne)構(gòu)成共生關(guān)系,因其寄生于牧草導致家畜中毒而最早被發(fā)現(xiàn)和研究,是牧草中研究得最多的兩種內(nèi)生真菌[3]。
據(jù)報道內(nèi)生真菌也可提高宿主植物對干早[4-5]、低溫[6]、離子毒害[7]、鹽堿[8-10]等非生物脅迫的抗性。從而使內(nèi)生真菌感染的植株比未感染植株對有限的資源更具竟爭力。當植物受到水分脅迫時會導致植物氣孔關(guān)閉來減少蒸騰作用而短期保護植物免受缺水傷害。據(jù)研究高羊茅感染內(nèi)生真菌會影響植株根莖和氣孔[11-12],帶菌高羊茅比非帶菌高羊茅氣孔導度下降的更早更快[11,13]。帶菌導致這種現(xiàn)象的具體機理還不清楚,但科研人員研究發(fā)現(xiàn)水分脅迫時從內(nèi)生真菌傳來的生物化學信號轉(zhuǎn)導或激素的改變將會更快的傳到宿主植物葉片[14-15]。而對內(nèi)生真菌在高羊茅種子萌發(fā)時的影響研究較少,本研究用PEG6000模擬干旱滲透脅迫,探討內(nèi)生真菌對種子萌發(fā)的影響。
1實驗材料與方法
1.1實驗材料
實驗材料為高羊茅(Festuca arundanacea)凌志。購買于百綠草種公司。
非帶菌種子的獲得方法:用帶菌的高羊茅種子處理得到,先50 ℃水浴處理20 min。經(jīng)處理的高羊茅種子種植到花盆里,4周后每顆植株用苯胺藍多次鏡檢,留下不帶菌的植株收獲種子即為非帶菌種子。
1.2實驗方法
1.2.1種子PEG脅迫
在培養(yǎng)皿底(直徑12 cm)的口部鋪兩層濾紙,濾紙中間開一小孔,通過燈芯連接濾紙和下層培養(yǎng)皿里的液體,供試種子均勻擺于濾紙上,然后蓋上培養(yǎng)皿蓋。PEG-6000處理濃度分別為0%、5%、10%、15%、20%,每個處理5次重復。培養(yǎng)皿中裝入相應(yīng)的PEG-6000濃度,確保每個培養(yǎng)皿中裝入液體在同一個高度,并用記號筆標記,對照加入同等液面的蒸餾水。為了控制PEG-6000濃度保持不變,每天補加蒸發(fā)的水分到記號線。種子開始發(fā)芽時(實驗開始的第5天)統(tǒng)計發(fā)芽數(shù),以后每隔兩天統(tǒng)計直到發(fā)芽最多的那天停止統(tǒng)計。發(fā)芽試驗總共持續(xù)16天。
1.2.2計算方法
發(fā)芽率=發(fā)芽終期全部正常種苗數(shù)/供試種子數(shù)×100%[16]。
16 d 后, 每個重復隨機選取10株幼苗, 測量胚芽長與胚根長。
1.2.3數(shù)據(jù)處理
數(shù)據(jù)采用excel 2007 和SPSS19處理
2結(jié)果與分析
2.1高羊茅種子中內(nèi)生真菌檢測結(jié)果
根據(jù)李春杰的檢菌方法[17],用氫氧化鈉浸泡種子后壓片,苯胺藍染色鏡檢。鏡檢結(jié)果如圖1。

圖1 高羊茅種子內(nèi)生真菌菌絲照片(40×)
種子中能在顯微鏡中看到如圖1箭頭所指示的菌絲即為帶菌種子。供試種子帶菌率結(jié)果如表1。直接購買的市售種子帶菌率為98%。經(jīng)溫度處理后再從非感染植株收獲的二代種子,完全不帶菌。

表1 高羊茅種子內(nèi)生真菌帶菌率鏡檢結(jié)果
2.2PEG脅迫對帶菌和不帶菌種子發(fā)芽率影響
由圖2可知,隨著PEG6000模擬干旱脅迫的加劇,帶菌高羊茅種子(E+)和不帶菌高羊茅種子(E-)的發(fā)芽率變化相似,均呈下降趨勢,在5%的輕度干旱脅迫下發(fā)芽率均接近對照,與對照差異不顯著。在干旱脅迫濃度達到10%后,E+和E-的發(fā)芽率均急劇下降,且呈現(xiàn)出E+發(fā)芽率高于E-,在15% PEG脅迫處理時,E+種子發(fā)芽率顯著高于E-種子(P<0.05)。 當PEG6000脅迫濃度達到20%時,E+和E-種子幾乎不能萌發(fā),因為每皿萌發(fā)種子數(shù)都少于10,未對其進行胚芽長與胚根長統(tǒng)計。

圖2 不同濃度 PEG6000脅迫后高羊茅種子發(fā)芽率
注:不同處理中,同一E+或E-中,標有不同小寫字母者差異顯著(p<0.05)。
E+、E-相同處理中兩兩差異顯著者用“*”標出,下同
2.3PEG脅迫對胚芽長和胚根長的影響

圖3 不同濃度PEG6000 脅迫后高羊茅種子胚芽長度
如圖3所示,高羊茅的胚芽長均隨PEG 脅迫濃度的升高呈現(xiàn)出明顯下降趨勢。根據(jù)統(tǒng)計分析結(jié)果顯示,高羊茅種子萌發(fā)時胚芽長度受到PEG 濃度的顯著影響,每增加5%的PEG 濃度均顯著的降低了種子胚芽長(P<0. 05)。而內(nèi)生真菌沒有表現(xiàn)出明顯促進胚芽長,經(jīng)單變量兩因素統(tǒng)計分析,影響高羊茅種子胚芽長的主效應(yīng)因素是PEG(F=151. 986, P=0. 000),而內(nèi)生真菌對芽長的影響不顯著(F=0. 198, P=0. 659),但PEG 和內(nèi)生真菌之間存在交互作用(F=4. 994,P=0. 06)。
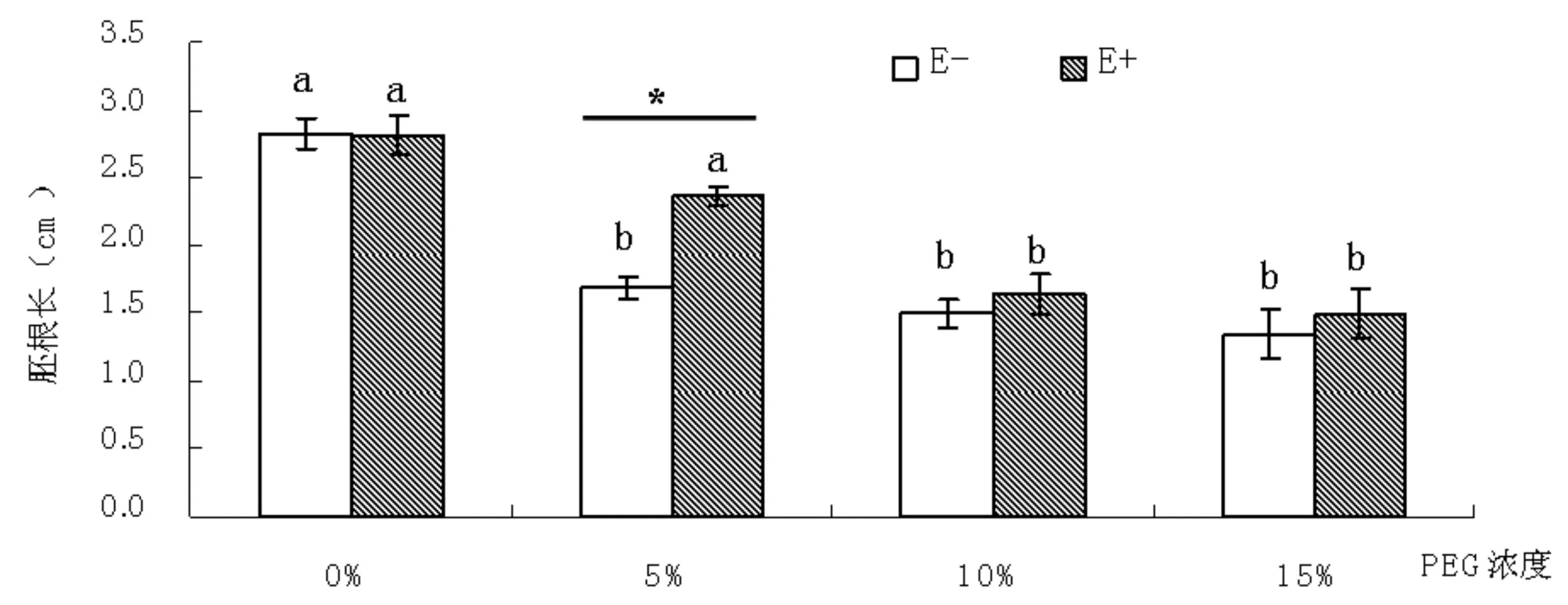
圖4 不同濃度PEG6000 脅迫后高羊茅種子胚根長度
如圖4所示,胚根長也隨著PEG6000脅迫濃度的增加呈現(xiàn)出下降趨勢。不帶菌的高羊茅種子,在5%濃度PEG脅迫開始,胚根長就顯著的低于對照。未處理時,帶菌和不帶菌種子的胚根長度無顯著差異,但PEG6000脅迫時,萌發(fā)種子的胚根長度呈現(xiàn)出帶菌種子胚根長大于不帶菌種子的。在5% PEG 6000處理下帶菌的胚根長顯著高于不帶菌的,說明內(nèi)生真菌感染的高羊茅種子,能在輕度干旱脅迫條件下幼苗胚根長顯著提高。用SPSS進行了單變量雙因素分析,結(jié)果為PEG 為第一影響胚根長的因素(F=34. 319,P=0. 000),內(nèi)生真菌也顯著影響了高羊茅種子萌發(fā)的胚根長(F=5. 999,P=0. 02),但PEG 和內(nèi)生真菌間沒有交互作用(F=1. 731,P=0. 18)。
3討論
3.1試驗采用PEG6000模擬干旱脅迫高羊茅種子發(fā)芽,結(jié)果為內(nèi)生真菌感染能提高種子在PEG模擬干旱脅迫下的發(fā)芽率,實驗結(jié)果為在15%濃度的PEG模擬干旱脅迫下帶內(nèi)生真菌高羊茅種子萌發(fā)率顯著高于不帶菌種子,試驗結(jié)果與宋梅玲[5]的研究結(jié)果內(nèi)生真菌可以提高禾草在溫度和干旱脅迫下的發(fā)芽率一致。
3.2植物的生長不僅受到自身遺傳特性的控制,還受到眾多環(huán)境因子的影響,如光照、溫度、水分和土壤營養(yǎng)物質(zhì)等在時空上的差異[18],其中限制植物生長的最普遍因素之一即是水分,尤其在干旱的環(huán)境[19]。植物蒸騰失水與其根部吸水之間的收支關(guān)系稱為水分平衡。Turner等[20]將植物忍受干旱脅迫的方式劃分為三個基本類型,即:避旱(短生植物)、延遲脫水耐旱、忍耐脫水耐旱。延遲脫水耐旱型:這類植物根系生長迅速,擴展范圍深廣,增加了與土壤的接觸面和吸收面積,主要通過限制水分的損失或保持水分的吸收來延遲脫水的發(fā)生。本試驗結(jié)果在5%的PEG濃度下,感染內(nèi)生真菌種子的胚根長度顯著高于不帶菌的,且統(tǒng)計分析結(jié)果為內(nèi)生真菌能顯著影響胚根長度(F=5. 999,P=0. 02),這一結(jié)果與鮑根生等[21]的研究結(jié)果一致。說明內(nèi)生真菌也是通過促進幼苗根的長度來減緩幼苗受到的水分脅迫。而高羊茅幼苗的胚芽長沒有出現(xiàn)顯著高于非帶菌的,分析原因可能由于營養(yǎng)分配導致的,種子萌發(fā)時營養(yǎng)都來自于種子自己胚乳的營養(yǎng),而這個營養(yǎng)是有限的,內(nèi)生真菌首先促進了胚根生長,而胚芽的長度就沒有顯著變化。
3.3種子萌發(fā)關(guān)系到植物是否能成苗,內(nèi)生真菌不但會在苗期或生長后期感染植物,而且植物被感染之后,內(nèi)生真菌還可能留在種子里形成種子內(nèi)生真菌這對于農(nóng)業(yè)制種和生態(tài)保育非常有意義,尤其是像高羊茅等禾本科牧草,他們所帶的內(nèi)生真菌為種帶內(nèi)生真菌,即內(nèi)生真菌的遺傳是通過種子進行垂直傳播[22]。如果能研究出提高抗旱性的帶新菌系高羊茅種子,為南疆沙漠和荒漠地的恢復和綠化將是一大貢獻。
3.4本實驗只是進行了PEG脅迫高羊茅凌志種子萌發(fā)方面的試驗,初步得出內(nèi)生真菌對高羊茅種子萌發(fā)具有促進作用,內(nèi)生真菌具體通過什么機理促進了胚根長來適應(yīng)干旱脅迫還有待于進一步深入研究。
4結(jié)論
內(nèi)生真菌感染可以顯著提高15%PEG6000干旱脅迫下高羊茅種子的發(fā)芽率和5%PEG 濃度下的胚根長。表明內(nèi)生真菌的存在可以使其宿主禾草在干旱條件下種子發(fā)芽能力具有優(yōu)勢, 以保證禾草在逆境中的生長。如果高羊茅草坪建植在干旱條件下帶菌種子與不帶菌種子相比,內(nèi)生真菌感染種子具有萌發(fā)能力強的特性。尤其是內(nèi)生真菌感染提高了幼苗的胚根長度進而促進高羊茅植株根的生長,使其在干旱環(huán)境下將更多的營養(yǎng)用于根系生長,從而提高了高羊茅的抗旱性。
參考文獻
[1]李飛,李春杰. 內(nèi)生真菌對禾草類植物抗旱性的影響[J]. 草業(yè)科學,2006(03):57-62.
[2]張殿忠,汪沛洪,趙會賢. 測定小麥葉片游離脯氨酸含量的方法[J]. 植物生理學通訊,1990(04):62-65.
[3]梁瑋莎,易建平,周而勛. 高羊茅和多年生黑麥草內(nèi)生真菌的分子檢測[J]. 菌物研究,2006(03):96-97.
[4]Kane K H. Effects of endophyte infection on drought stress tolerance of Lolium perenne accessions from the Mediterranean region[J]. Environmental and Experimental Botany , 2011(71): 337- 344.
[5]宋梅玲,李春杰,彭清青等. 溫度和水分脅迫下內(nèi)生真菌對野大麥種子發(fā)芽的影響[J]. 草地學報,2010(06):833-837.
[6]陳娜. 醉馬草遺傳多樣性及內(nèi)生真菌對其抗寒性影響[D].蘭州大學,2008.
[7]Malinowski DP, Belesky DP. Adaptation of endophyte-infected cool-season grasses to environment stresses: Mechanisms of drought and mineral stress tolerance[J]. Crop Science ,2000, 40(4): 923-940.
[8]Helmut B,Jozsef F,Borbala D H,et al. Salt tolerance of barley induced by the root endophyte Piriformospora indica is associated with a strong increase in antioxidants[J].New phytologist,2008(180):501-510.
[9]Sabzalian MR and Mirlohi A. Neotyphodium endophytes trigger salt resistance in tall and meadow fescues[J].J.Plant Nutr.Soil Sci,2010(173):952-957.
[10]王正鳳,李春杰,金文進等. 內(nèi)生真菌對野大麥耐鹽性的影響[J]. 草地學報,2009(01):88-92.
[11]Elmi A A, West C.P, Endophyte infection effects on stomatal conductance, osmotic adjustment and drought recovery of tall fescue[J]. New Phytol. 1995(131):61-67.
[12]Buck G W, West C.P, and H.W. Elbersen. Endophyte effect on drought tolerance in diverse Festuca species[M]. In C.W. Bacon and N.S. Hill (ed.) Neotyphodium/grass interactions. Plenum Press, New York. 1997. p. 141-143.
[13]Elbersen H W, Buck G W, West C P, and R.E. Joost. Water loss from tall fescue leaves is decreased by endophyte[J]. Arkansas Farm Res. 1994(43):8-9.
[14]Belesky D P, Devine O J, Pallas J E, et al. Stringer. Photosynthetic activity of tall fescue as influenced by a fungal endophyte[J]. Photosynthetica.1987a.21:82-87.
[15]West C P, Physiology and drought tolerance of endophyte-infected grasses[M]. CRC Press, Boca Raton, FL. 1994:87-99.
[16]韓建國.實用牧草種子學[M].北京:中國農(nóng)業(yè)大學出版社,1997:133.
[17]李春杰, 南志標, 劉勇等.醉馬草內(nèi)生真菌檢測方法的研究[J].中國食用菌, 2008,27( suppl.):16-19.
[18]肖春旺,周廣勝,馬鳳云.施水量變化對毛烏素沙地優(yōu)勢植物形態(tài)與生長的影[J].植物生態(tài)學報,2002,26(1):69-76.
[19]Lange O L,KapPen L,Schulze E D.Water and Plant life: Problems and modern approaches[M].Berlin:Heidelbergs Springer Verlag,l976.
[20]Turner N C, Stern W R, Evans P. Water Relations and Osmotic Adjustment of Leaves and Roots of lupins inresponse to water deficits [J].Crop scienee,1987(27):977-983.
[21]Bao Gensheng Saikkonen, Kari Wang Hongsheng, et al.. Does endophyte symbiosis resist allelopathic effects of an invasive plant in degraded grassland [J]. Fungal Ecology,2015(17): 114-125.
[22]Siegel M R , Johnson M C , Varney D R , et al. A fungal endophyte in tall fescue : incidence and dissemination[J]. Phytopathology , 1984, 74(3) : 932~937.
Effects of Endophytic Fungi on Drought Tolerance Ability of Tall Fescue Seed
Chen Zhihao1Liu Xuefeng2Bao Gensheng3Li Xiaoxia1Ji Jianqiang4Chen Shuihong4*
(1 College of Znimal Science of Tarim University, Alar, Xinjiang 843300 )(2 The 1st Pasturage Farm of No.14 Agriculture Division of Xinjiang Production and Construction Crops,Cele, Xinjiang 848306 )(3 Qinghai Academy of Animal Science and Veterinary Medicine, Xining, Qinghai 810016)(4 College of life science, Tarim University, Alar, Xinjiang 843300 )
AbstractIn their evolution, grasses have developed symbiotic associations with fungi that systemically infect grass shoots. Among the latter are grass endophytes, defined as fungi that live their entire life cycle within the aerial portion of the host grass, they do not cause any plant disease symptoms. Endophyte infected can improve biotic and abiotic stress resistance on the host plant. we studied the tall fescue seed of endophyte infection (E+) and endophyte free (E-) on germination under different concentrations PEG6000 stress. The results showed that the endophyte can improve the germination activity, and increased root length under PEG-6000 drought stress. The endophyte infected seed will be better adapted to drought environments.
Key wordsEndophyte; tall fescue; drought resistance; germination rate
中圖分類號:S543+9
文獻標識碼:A
DOI:10.3969/j.issn.1009-0568.2016.02.003
文章編號:①1009-0568(2016)02-0012-06
作者簡介:陳志豪(1991-),男,2016級本科生,研究方向為草業(yè)科學。E-mail:569701832@qq.com*為通訊作者E-mail:rainbow4@126.com
基金項目:塔里木大學大學生創(chuàng)新項目(2015013)。
收稿日期:①2015-09-02
