腦CYP2E1參與脂多糖誘導(dǎo)的神經(jīng)元損傷
2016-08-10 06:56:40那淑芳楊哲瓊
中國藥理學(xué)通報 2016年7期
那淑芳,姚 慧,李 杰,楊哲瓊,樂 江
(武漢大學(xué)基礎(chǔ)醫(yī)學(xué)院藥理學(xué)系,湖北 武漢 430071)
?
腦CYP2E1參與脂多糖誘導(dǎo)的神經(jīng)元損傷
那淑芳,姚慧,李杰,楊哲瓊,樂江
(武漢大學(xué)基礎(chǔ)醫(yī)學(xué)院藥理學(xué)系,湖北 武漢430071)
目的研究脂多糖(LPS)所致炎癥與腦細(xì)胞色素P450 2E1和CYP2E1之間的相互影響。方法采用具有膽堿能神經(jīng)元特征的IMR-32人神經(jīng)母細(xì)胞瘤細(xì)胞系,分別給予低劑量(0.1 mg·L-1)和高劑量(1.0 mg·L-1)LPS處理24 h,檢測LDH和SOD活性。在LPS處理前45 min,分別加入p38抑制劑SB203580和ERK抑制劑U0126處理IMR-32細(xì)胞,觀察MAPK信號系統(tǒng)對神經(jīng)元CYP2E1表達(dá)的影響。采用具有多巴胺能神經(jīng)元特征的SH-SY5Y人神經(jīng)母細(xì)胞瘤細(xì)胞系建立高表達(dá)CYP2E1細(xì)胞系,并與正常SH-SY5Y細(xì)胞同時給予低劑量(0.1 mg·L-1)和高劑量(1.0 mg·L-1)LPS處理24 h后檢測LDH和SOD活性。結(jié)果與對照組相比,高劑量LPS處理IMR-32細(xì)胞,SOD的活力下降15.0%(P<0.01),LDH上升1.38倍(P<0.01),CYP2E1 mRNA升高1.25倍(P<0.01),蛋白水平升高1.19倍(P<0.05)。p38和ERK抑制劑可拮抗高劑量LPS對CYP2E1的誘導(dǎo)作用。低劑量LPS處理CYP2E1高表達(dá)SH-SY5Y細(xì)胞,LDH的升高幅度較非高表達(dá)的對照組上升了1.28倍(P<0.01),SOD活力下降幅度增加了3.53倍(P<0.01);高劑量LPS使得CYP2E1高表達(dá)SH-SY5Y細(xì)胞LDH的升高幅度較非高表達(dá)的對照組上升了1.54倍(P<0.01),SOD活力下降幅度增加了2.17倍(P<0.01)。結(jié)論LPS可上調(diào)神經(jīng)元所表達(dá)的CYP2E1水平,其調(diào)控作用可能與ERK和p38信號傳導(dǎo)通路相關(guān)。高表達(dá)CYP2E1加劇LPS對神經(jīng)元的損傷,提示CYP2E1參與了炎癥所致神經(jīng)細(xì)胞損傷的病理過程。
脂多糖;炎癥;神經(jīng)元;ERK信號傳導(dǎo)通路;p38信號傳導(dǎo)通路
隨著人口老年化,神經(jīng)退行性疾病已成為危害人類健康的重要疾病。研究提示,帕金森病、阿爾茨海默病、多發(fā)性硬化、肌萎縮側(cè)索硬化癥等神經(jīng)退行性疾病的病理生理學(xué)改變與持續(xù)的炎癥反應(yīng)過程密切相關(guān)[1-3],然而其病理機(jī)制尚需深入研究。
細(xì)胞色素P450(cytochrome P450,CYP)超家族酶系參與內(nèi)源性物質(zhì)(如脂肪酸、維生素、膽酸)和外源性物質(zhì)(如藥物、毒物、前致癌物)的代謝失活及活化。CYP超家族酶系雖主要表達(dá)在肝臟,但也表達(dá)于肝外器官,如腦。在CYP亞型中,CYP2E1蛋白空間結(jié)構(gòu)具有可變性,故易出現(xiàn)底物與酶解偶聯(lián)而產(chǎn)生活性自由基[4]。2012年,德國學(xué)者對帕金森病患者大腦皮層和殼核的14495個基因的CpG島的甲基化水平進(jìn)行分析,發(fā)現(xiàn)僅CYP2E1基因甲基化水平明顯下降,mRNA水平明顯升高[5]。
脂多糖(LPS)為革蘭陰性細(xì)菌外壁層中一種特有的化學(xué)成分[6],能夠刺激體內(nèi)多種細(xì)胞合成和釋放眾多內(nèi)源性生物活性因子,導(dǎo)致炎癥反應(yīng)發(fā)生[7]。文獻(xiàn)報道,LPS腹腔注射、顱內(nèi)注射均可用于建立動物神經(jīng)炎癥模型。IMR-32細(xì)胞為具有膽堿能神經(jīng)元特征的人神經(jīng)母細(xì)胞瘤細(xì)胞系; SH-SY5Y細(xì)胞為具有多巴胺能神經(jīng)元特征的人神經(jīng)母細(xì)胞瘤細(xì)胞系。文獻(xiàn)報道,腦CYP2E1在IMR-32細(xì)胞和SH-SY5Y細(xì)胞中均具有表達(dá)。本文擬利用上述人神經(jīng)母細(xì)胞瘤細(xì)胞系,觀察腦CYP2E1在LPS所致神經(jīng)元損傷中的作用。
1 材料與方法
1.1細(xì)胞培養(yǎng)和試劑IMR-32和SH-SY5Y人神經(jīng)母細(xì)胞瘤細(xì)胞,置于恒溫37℃,含5% CO2的培養(yǎng)箱中進(jìn)行培養(yǎng)。培養(yǎng)基分別為含10%胎牛血清(Gibco公司)及1%青霉素-鏈霉素雙抗生素(碧云天生物技術(shù)研究所)的MEM培養(yǎng)基或DMEM培養(yǎng)基(杭州吉諾生物醫(yī)藥技術(shù)有限公司)。含EDTA胰蛋白酶用于細(xì)胞消化、傳代(杭州吉諾生物醫(yī)藥技術(shù)有限公司)。考馬斯亮蘭試劑盒、總超氧化物歧化酶(T-SOD)測試盒、乳酸脫氫酶(LDH)測定試劑盒,購于南京生物建成生物工程研究所。LPS(來源于大腸桿菌055:B5)、DMSO、SB203580、U0126,購于Sigma公司;所用兔抗大鼠多克隆CYP2E1抗體,購自Millipore公司。辣根過氧化物酶(HRP)標(biāo)記的羊抗兔IgG單克隆抗體購自碧云天生物技術(shù)研究所。
1.2細(xì)胞分組及處理選取生長良好細(xì)胞制成細(xì)胞懸液,接種于6孔和24孔細(xì)胞培養(yǎng)板。待細(xì)胞貼壁后,換液,分別給予低劑量(0.1 mg·L-1)LPS、高劑量(1.0 mg·L-1)LPS或等體積PBS溶液,孵育24 h。抑制性實(shí)驗中,細(xì)胞給予ERK抑制劑U0126(10 μmol·L-1)或p38抑制劑SB203580(20 μmol·L-1),45 min后分別加入低或高劑量LPS,共孵育24 h。
1.3構(gòu)建高表達(dá)CYP2E1的SH-SY5Y人神經(jīng)母瘤細(xì)胞系構(gòu)建pcDNA3.1-CYP2E1重組質(zhì)粒,利用LipofectamineTM3000轉(zhuǎn)染試劑,將人CYP2E1 cDNA表達(dá)質(zhì)粒轉(zhuǎn)入SH-SY5Y細(xì)胞中。轉(zhuǎn)染48 h后,胰蛋白酶消化細(xì)胞,接種至培養(yǎng)皿,用含10%胎牛血清和100 mg·L-1G418的DMEM培養(yǎng)基繼續(xù)培養(yǎng)[8]。兩周后,將孤立克隆轉(zhuǎn)移到6孔板中,繼續(xù)傳代培養(yǎng)。通過RT-PCR和Western blot鑒定每個克隆CYP2E1表達(dá),以獲得穩(wěn)定表達(dá)CYP2E1的細(xì)胞系。
1.4乳酸脫氫酶(LDH)和超氧化物歧化酶(SOD)活性測定將IMR-32細(xì)胞、高表達(dá)CYP2E1的SH-SY5Y細(xì)胞制成細(xì)胞懸液分別接種24孔板,培養(yǎng)24 h待貼壁。細(xì)胞給予低濃度(0.1 mg·L-1)、高濃度LPS(1.0 mg·L-1)或等體積PBS溶液處理,每組8孔,繼續(xù)培養(yǎng)24 h。收集培養(yǎng)基上清液,按試劑盒說明書檢測培養(yǎng)基中LDH水平,可見光分光光度計測定440 nm處吸光度[9]。
按SOD檢測試劑盒說明書檢測培養(yǎng)基中SOD水平。黃嘌呤及黃嘌呤氧化酶系產(chǎn)生超氧陰離子,氧化羥胺生成亞硝酸銨鹽,在顯色劑作用下呈紫紅色,利用可見光分光光度計550 nm處測定吸光度。
1.5Real-time RT-PCRTRIzol(Incitrogen公司)提取IMR-32細(xì)胞總RNA,使用Revert Aid First Strand cDNA synthesis試劑盒將總RNA逆轉(zhuǎn)錄為cDNA。設(shè)計人CYP2E1引物序列:5′-CCGCTTCCCATCATCGGGAAC,3′-GCGCTTCCTTCACCGCCTTGT。RPLP0作為內(nèi)參,引物序列為:5′-GGCGACCTGGAAGTCCAACT,3′-CCATCAGCACCACAGCCTTC[10]。退火溫度為60℃,循環(huán)數(shù)為30。1.6Western blot收集細(xì)胞,PBS洗滌細(xì)胞,加入RIPA裂解液,冰上裂解30 min。使用考馬斯亮藍(lán)法測定蛋白濃度,并將各組樣品稀釋至相同蛋白濃度。取30 μg總蛋白,加入上樣緩沖液等體積混合,置于100℃沸水中變性處理5 min。樣品經(jīng)SDS-PAGE凝膠電泳,電泳結(jié)束后轉(zhuǎn)印至PVDF膜上。用5% PBST配制的脫脂奶粉,在37℃封閉2 h,加入兔抗大鼠多克隆CYP2E1抗體(1 ∶1 500)孵育2 h。PBST洗滌3次,每次10 min。加入辣根過氧化物酶(HRP)標(biāo)記的羊抗兔IgG單克隆抗體(1 ∶7 000),37℃孵育1 h。PBST洗滌3次,每次10 min。采用辣根過氧化物酶-ECL化學(xué)發(fā)光顯影,曝光膠片。掃描膠片,用灰度分析軟件Alpha EaseFC分析目的條帶。
2 結(jié)果
2.1LPS引發(fā)IMR-32神經(jīng)細(xì)胞出現(xiàn)損傷與對照組相比,低劑量(0.1 mg·L-1)、高劑量(1.0 mg·L-1)LPS可使得IMR-32神經(jīng)細(xì)胞LDH分別升高1.18倍(P<0.05)和1.38倍(P<0.01)。然而,與對照組相比,高劑量LPS致SOD的活力下降15.0%(P<0.01),但低劑量LPS對SOD的活力無明顯影響(Fig 1)。
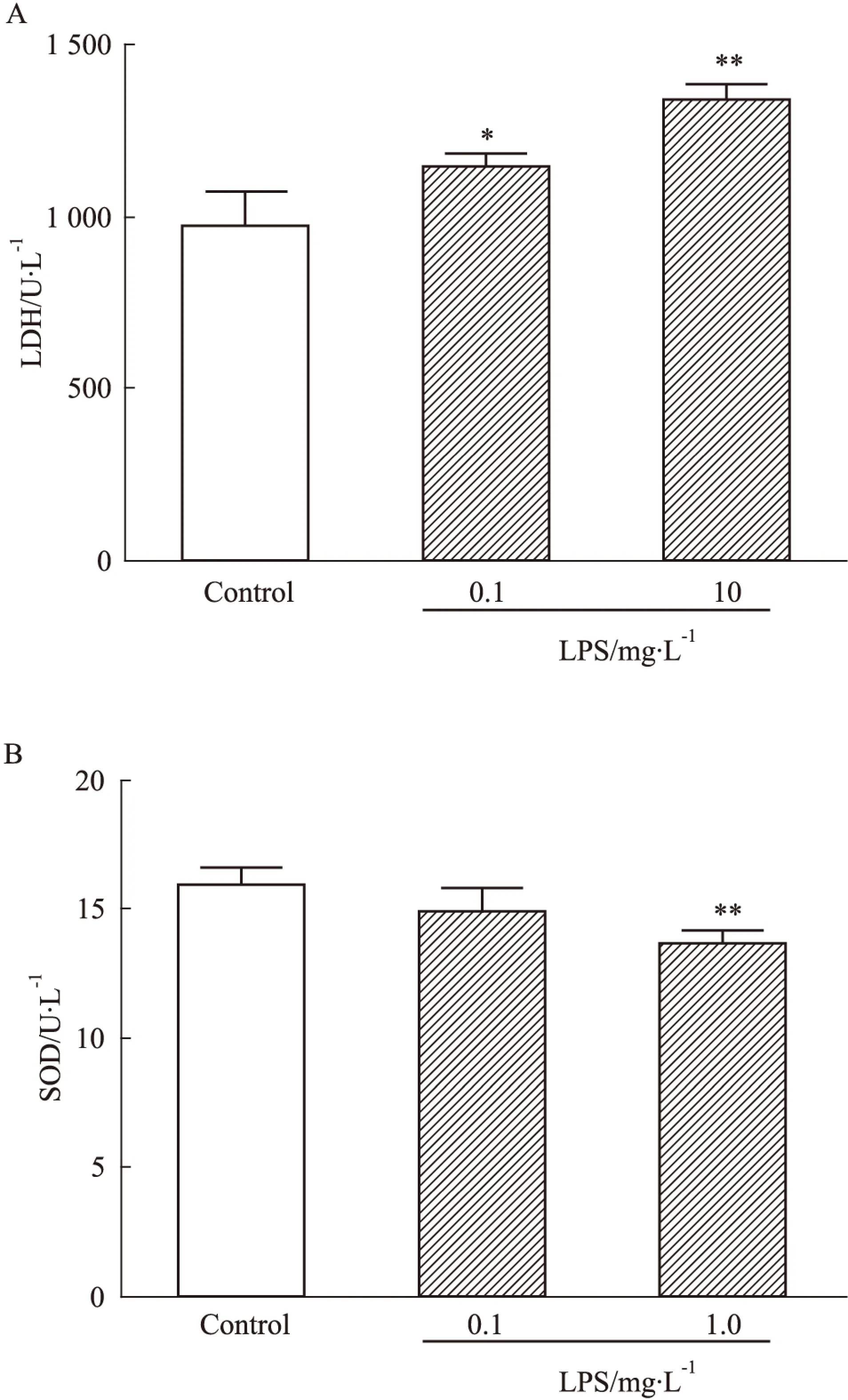
Fig 1 Effect of LPS treatment on levels of LDH(A) and SOD(B) in IMR-32 neuroblastoma cells
*P<0.05,**P<0.01vscontrol
2.2LPS對IMR-32神經(jīng)細(xì)胞CYP2E1 表達(dá)水平的影響與正常對照組相比,低劑量(0.1 mg·L-1)LPS對IMR-32細(xì)胞CYP2E1 mRNA水平、蛋白表達(dá)水平無明顯影響。高劑量(1.0 mg·L-1)LPS可明顯誘導(dǎo)IMR-32細(xì)胞CYP2E1 mRNA水平,其mRNA水平較對照組升高1.25倍(P<0.01)(Fig 2A)。Western blot實(shí)驗結(jié)果表明,高劑量LPS處理組細(xì)胞CYP2E1蛋白水平較對照組升高1.19倍(P<0.05)(Fig 2B)。

Fig 2 Alteration of CYP2E1 mRNA and protein in IMR-32 neuroblastoma cells after LPS treatment
A: Effect of LPS on the levels of CYP2E1 mRNA in IMR-32 neuroblastoma cell. B: Effect of LPS on the levels of CYP2E1 protein in IMR-32 neuroblastoma cell. Representative Western blot of CYP2E1 protein is shown above, and the protein relative density is shown as the histogram.*P<0.05,**P<0.01vscontrol.
2.3ERK和p38抑制劑可拮抗LPS對CYP2E1 mRNA的誘導(dǎo)作用ERK抑制劑U0126(10 μmol·L-1)和p38抑制劑SB203580(20 μmol·L-1)單獨(dú)處理IMR-32細(xì)胞,未見CYP2E1mRNA水平出現(xiàn)變化(Fig 3A)。IMR-32細(xì)胞分別給予不同抑制劑45 min后,加入1.0 mg·L-1LPS共孵育,U0126和SB203580使得細(xì)胞CYP2E1 mRNA水平分別下降20.6%(P<0.01)和24.2%(P<0.01)(Fig 3B)。結(jié)果提示,LPS可能通過激活ERK和p38通路誘導(dǎo)CYP2E1表達(dá)。
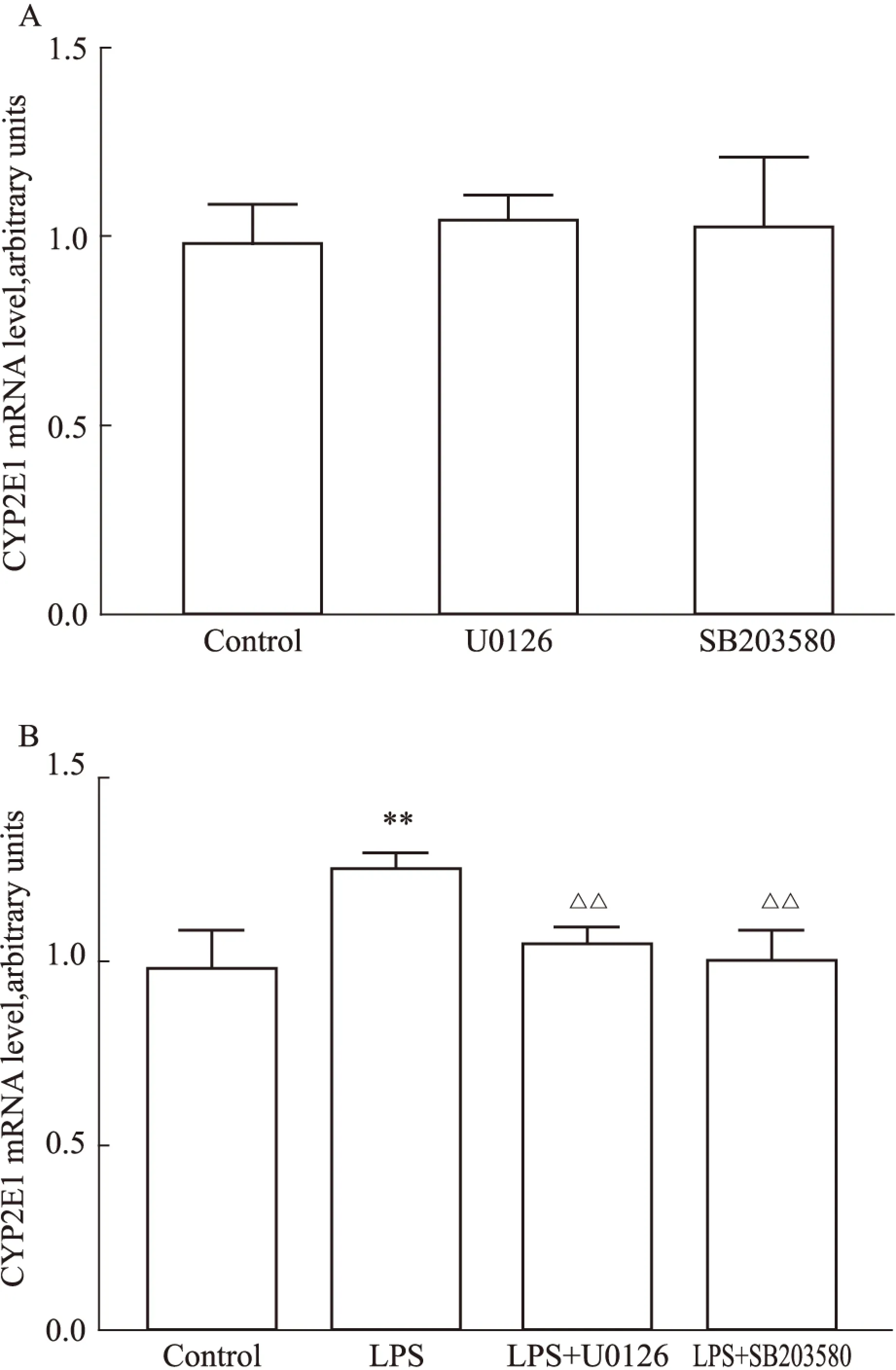
Fig 3 Effects of ERK and p38 inhibitors on LPS-induced CYP2E1 mRNA levels in IMR-32 neuroblastoma cells
A: The levels of CYP2E1 mRNA had no obvious change after the treatment of SB203580 and U0126. B: The up-regulation of CYP2E1 by LPS could be attenuated by SB203580 and U0126 pretreatment.**P<0.01vscontrol;△△P<0.01vsLPS
2.4SH-SY5Y神經(jīng)細(xì)胞高表達(dá)CYP2E1加劇LPS所致細(xì)胞損傷Real-time PCR和Western blotting證實(shí),高表達(dá)CYP2E1的SH-SY5Y細(xì)胞系構(gòu)建成功(Fig 4A,B)。與正常對照組相比,低劑量(0.1 mg·L-1)LPS處理SH-SY5Y細(xì)胞,LDH水平升高1.55倍(P<0.01),SOD活力下降8.3%;而高表達(dá)CYP2E1的SH-SY5Y細(xì)胞,LDH水平升高1.98倍(P<0.01),SOD活力下降29.3%(P<0.01)。與此同時,與正常對照組相比,高劑量(1.0 mg·L-1)LPS使得LDH水平升高2.17倍(P<0.01),SOD活力下降20.4%(P<0.01);而高表達(dá)CYP2E1的SH-SY5Y細(xì)胞,LDH水平升高3.34倍(P<0.01),SOD活力下降44.2%(P<0.01)。研究結(jié)果提示,高表達(dá)CYP2E1可加劇LPS所致神經(jīng)細(xì)胞損傷。
3 討論
實(shí)驗結(jié)果表明,LPS可在轉(zhuǎn)錄水平上調(diào)神經(jīng)元CYP2E1表達(dá),其誘導(dǎo)機(jī)制可能與激活MAPK信號通路有關(guān),而高表達(dá)CYP2E1可加重LPS所致神經(jīng)元損傷。目前,由炎癥介導(dǎo)的CYP2E1的變化在肝內(nèi)的調(diào)控機(jī)制已較為明確,但是對中樞內(nèi)CYP2E1的變化情況研究較少。在大鼠肝癌細(xì)胞Fao細(xì)胞系的體外實(shí)驗中,證實(shí)炎癥因子通過多重機(jī)制下調(diào)CYP2E1基因的表達(dá)[11]。炎癥因子刺激人肝臟原代培養(yǎng)細(xì)胞后也發(fā)現(xiàn),CYP2E1 mRNA水平下降。以往研究提示,炎癥因子可能調(diào)控HNF-1α和其他轉(zhuǎn)錄因子作用于CYP2E1的5′-端上游調(diào)控區(qū)域,或影響CYP2E1 mRNA的穩(wěn)定性而下調(diào)其表達(dá)。有報道[12],卡介苗制備的免疫性肝炎模型中,因NF-κB信號轉(zhuǎn)導(dǎo)通路活化而抑制CYP2E1的下調(diào)。
本文數(shù)據(jù)表明,炎癥對腦CYP2E1表達(dá)調(diào)控機(jī)制不同于肝臟。不同劑量(0.1 mg·L-1和1.0 mg·L-1)LPS處理IMR-32細(xì)胞,可導(dǎo)致細(xì)胞出現(xiàn)損傷,且誘導(dǎo)CYP2E1 mRNA和蛋白水平表達(dá)。分裂原激活的蛋白激酶(MAPKs)是絲氨酸/蘇氨酸蛋白激酶家族,包括細(xì)胞外信號調(diào)節(jié)激酶(ERK)、c-Jun Np-末端蛋白激酶(JNK)和p38 MAPK等[13]。MAPK信號通路參與細(xì)胞增殖、存活和凋亡。本研究表明,ERK和p38抑制劑均可不同程度拮抗LPS對CYP2E1的誘導(dǎo)效應(yīng)。結(jié)果提示,LPS可能通過激活ERK和p38信號傳導(dǎo)通路,繼而影響下游轉(zhuǎn)錄因子,上調(diào)神經(jīng)元CYP2E1表達(dá)水平。文獻(xiàn)報道[14],MAPK信號通路參與了乙醇對神經(jīng)膠質(zhì)細(xì)胞CYP2E1的表達(dá)調(diào)控。
低劑量(0.1 mg·L-1)LPS處理SH-SY5Y細(xì)胞,高表達(dá)CYP2E1后LDH的升高幅度上升了1.28倍(P<0.01),SOD活力下降幅度增加了3.53倍(P<0.01);高劑量(1.0 mg·L-1)LPS處理細(xì)胞,高表達(dá)CYP2E1后LDH的升高幅度上升了1.54倍(P<0.01),SOD活力下降幅度增加了2.17倍(P<0.01)。實(shí)驗數(shù)據(jù)表明,CYP2E1表達(dá)量增加可加劇神經(jīng)細(xì)胞的損傷。CYP2E1具有較高的NADPH氧化活性,代謝過程中會產(chǎn)生活性氧,如羥自由基(·OH)、超氧陰離子(O2-)和過氧化氫(H2O2)等,使得活性氧ROS產(chǎn)生增多。當(dāng)活性氧含量超過機(jī)體清除能力時,則發(fā)生氧化損傷,破壞膜結(jié)構(gòu)的完整性,改變膜結(jié)合酶,膜通透性增加,流動性改變,損傷線粒體,破壞DNA修飾,使細(xì)胞內(nèi)環(huán)境紊亂,能量衰竭,膜功能喪失[5]。在腹腔注射神經(jīng)毒素1-甲基-4-苯基-1,2,3,6-四氫吡啶(MPTP)制備的帕金森病動物模型中,CYP2E1基因敲除小鼠所出現(xiàn)的多巴胺能神經(jīng)元損傷程度明顯弱于對照組[15]。腦CYP2E1可能是神經(jīng)炎癥引發(fā)神經(jīng)細(xì)胞損傷的重要中央環(huán)節(jié)。腦CYP2E1在與炎癥相關(guān)的神經(jīng)退行性疾病病理生理過程中的作用仍需進(jìn)一步研究。
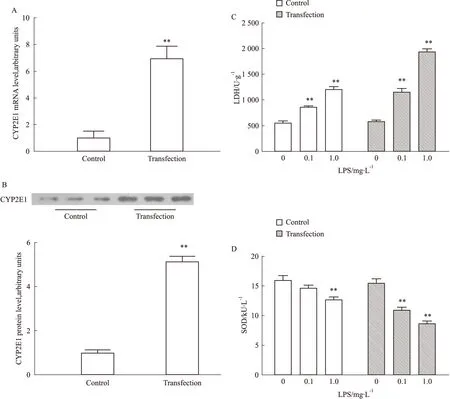
Fig 4 Effect of LPS treatment on levels of LDH and SOD in SH-SY5Y neuroblastoma cells overexpressing CYP2E1
A: Transfection increased CYP2E1 mRNA levels in SH-SY5Y cells. B: Transfection increased CYP2E1 protein levels in SH-SY5Y cells. Representative Western blot of CYP2E1 protein is shown above, and the protein relative density is shown as the histogram. C: The levels of LDH were aggrandized in SH-SY5Y cells overexpressing CYP2E1 after LPS treatment. D: The SOD activity in SH-SY5Y cells overexpressing CYP2E1 were reduced in LPS treatment.**P<0.01vscontrol.
(致謝:本研究在武漢大學(xué)基礎(chǔ)醫(yī)學(xué)院藥理系完成,得到國家自然基金項目資助,在此表示感謝!)
[1]Deleidi M, Isacson O. Viral and inflammatory triggers of neurodegenerative diseases[J].SciTranslMed, 2012, 4(121): 121ps3.
[2]Phani S, Loike J D, Przedborski S. Neurodegeneration and inflammation in Parkinson′s disease[J].ParkinsonismRelatDisord, 2012, 18 Suppl 1: S207-09.
[3]Glass C K, Saijo K, Winner B, et al. Mechanisms underlying inflammation in neurodegeneration[J].Cell, 2010, 140(6): 918-34.
[4]劉晨暉,樂江. 細(xì)胞色素P450 CYP2E1酶構(gòu)型特征及其表達(dá)調(diào)控機(jī)制的研究進(jìn)展[J]. 中國藥理學(xué)與毒理學(xué)雜志,2010,24(02): 155-60.
[4]Liu C H, Yue J. Progress of research on characteristics of cytochrome P-450 CYP2E1 in protein structure and regulation mechanisms[J].ChinJPharmacolToxicol,2010,24(02):155-60.
[5]Kaut O, Schmitt I, Wullner U. Genome-scale methylation analysis of Parkinson′s disease patients′ brains reveals DNA hypomethylation and increased mRNA expression of cytochrome P450 2E1[J].Neurogenetics,2012,13(1): 87-91.
[6]Wang G, Namura S. Effects of chronic systemic treatment with peroxisome proliferator-activated receptor alpha activators on neuroinflammation induced by intracerebral injection of lipopolysaccharide in adult mice[J].NeurosciRes, 2011, 70(2): 230-37.
[7]張雪梅,熊煥章. LPS誘導(dǎo)的炎癥反應(yīng)信號傳導(dǎo)通路研究進(jìn)展[J]. 中國獸醫(yī)雜志, 2010, 46(7):45-7.
[7]Zhang X M, Xiong H Z. Research progress in LPS induced inflammatory response of signaling transduction pathway[J].ChinJVeterMed,2010,46(7): 45-7.
[8]Leon-Buitimea A, Rodriguez-Fragoso L, Lauer F T,et al. Ethanol-induced oxidative stress is associated with EGF receptor phosphorylation in MCF-10A cells overexpressing CYP2E1[J].ToxicolLett, 2012, 209(2): 161-5.
[9]Zhong Y, Dong G, Luo H, et al. Induction of brain CYP2E1 by chronic ethanol treatment and related oxidative stress in hippocampus, cerebellum, and brainstem[J].Toxicology,2012,302(2-3): 275-84.
[10]Barzon L, Masi G, Pacenti M, et al. Expression of aromatase and estrogen receptors in human adrenocortical tumors[J].VirchowsArch,2008,452(2):181-91.
[11]Hakkola J, Hu Y, Ingelman-Sundberg M. Mechanisms of down-regulation of CYP2E1 expression by inflammatory cytokines in rat hepatoma cells[J].JPharmacolExpTher, 2003, 304(3): 1048-54.
[12]賈金雪, 秦金東, 李學(xué)峰, 等.鈍化NF-κB的活化對免疫性肝損傷大鼠CYP2E1的影響[J].中國藥理學(xué)通報,2015,31(8):1076-80.
[12]Jia J X, Qing J D, Li X F, et al. Effect of blunting NF kappa B activation on CYP2E1 in immunological liver injury rats[J].ChinPharmacolBull, 2015, 29(8):1076-80.
[13]Davis R J. The mitogen-activated protein kinase signal transduction pathway[J].JBiolChem,1993,268(20): 14553-56.
[14]Jin M, Ande A, Kumar A, Kumar S. Regulation of cytochrome P450 2e1 expression by ethanol: role of oxidative stress-mediated pkc/jnk/sp1 pathway[J].CellDeathDis, 2013, 4(3): e554.
[15]Viaggi C, Vaglini F, Pardini C, et al. MPTP-induced model of Parkinson′s disease in cytochrome P450 2E1 knockout mice[J].Neuropharmacology,2009,56(8): 1075-81.
Involvement of brain CYP2E1 in damage to neuronal cells induced by lipopolysaccharide
NA Shu-fang, YAO Hui, LI Jie, YANG Zhe-qiong, YUE Jiang
(DeptofPharmacology,BasicMedicalSchool,WuhanUniversity,Wuhan430071,China)
AimTo investigate the interactions between the neuroinflammation caused by lipopolysaccharide(LPS) and brain CYP2E1.MethodsThe human cholinergic neuroblastoma cell line IMR-32 was treated with LPS(0.1 mg·L-1, 1.0 mg·L-1), and the LDH and SOD activities were determined after 24 h incubation. In order to determine the roles of MAPK signaling pathway in the regulation of CYP2E1 by LPS, the IMR-32 cells were treated with p38 pathway inhibitor SB203580 or ERK pathway inhibitor U0126 for 45 min before the incubation with LPS. The human dopaminergic neuroblastoma cell line SH-SY5Y with CYP2E1 over-expression was established. The LDH and SOD activities were determined in SH-SY5Y cells over-expressed CYP2E1 and control cells treated with LPS(0.1 mg·L-1, 1.0 mg·L-1) for 24 h.ResultsThe levels of LDH in IMR-32 cells treated with high-dose LPS were increased by 1.38-fold(P<0.01) compared with the control group, and the levels of SOD reduced by 15.0%(P<0.01).Compared with the control, CYP2E1 mRNA and protein levels in IMR-32 cells treated with high-dose LPS were increased by 1.25-fold(P<0.01) and 1.19-fold(P<0.05). The up-regulation of CYP2E1 by LPS could be attenuated by SB203580 and U0126 pretreatment. Compared with the control cells, the CYP2E1 over-expression increased LDH levels by 1.28-fold(P<0.01) and decreased SOD levels by 3.53-fold(P<0.01) after the low-dose of LPS treatment. The CYP2E1 over-expression increased LDH levels by 1.54-fold(P<0.01) and decreased SOD levels by 2.17-fold(P<0.01) after the high-dose of LPS treatment, compared with the control cells.ConclusionsLPS can induce CYP2E1 mRNA and protein levels, and the p38 and ERK signaling pathway may be involved in the regulation. The elevated CYP2E1 levels aggravate the damage to neuronal cells caused by LPS. Brain CYP2E1 may be an important contributing factor to the pathological process of neuroinflammatory injury.
cytochrome P450 2E1; lipopolysaccharide; inflammation; neuron; ERK signaling pathway; p38 signaling pathway
2016-01-22,
2016-04-28
國家自然科學(xué)基金資助項目(No 30600773、No 31271327)
那淑芳(1989-),女,碩士生,研究方向:藥物代謝,E-mail:nashufang2009@126.com;楊哲瓊(1975-),女,博士,副教授,碩士生導(dǎo)師,通訊作者,研究方向:神經(jīng)藥理學(xué),E-mail:yzq751007@126.com;樂江(1975-),女,博士,教授,博士生導(dǎo)師,通訊作者,研究方向:神經(jīng)藥理學(xué),E-mail:yuejiang@whu.edu.cn
10.3969/j.issn.1001-1978.2016.07.010
A
1001-1978(2016)07-0932-06
R322.81;R329.25;R338.1;R345.99;R364.5
網(wǎng)絡(luò)出版時間:2016-6-20 11:49網(wǎng)絡(luò)出版地址:http://www.cnki.net/kcms/detail/34.1086.R.20160620.1149.020.html
猜你喜歡
課堂內(nèi)外·初中版(科學(xué)少年)(2023年10期)2023-12-10 00:43:06
全科護(hù)理(2022年10期)2022-12-26 21:19:15
中國合理用藥探索(2022年1期)2022-11-26 00:22:32
美與時代·美術(shù)學(xué)刊(2022年3期)2022-04-27 01:18:15
鄉(xiāng)村科技(2021年33期)2021-03-16 02:26:54
國際放射醫(yī)學(xué)核醫(yī)學(xué)雜志(2021年10期)2021-02-28 08:41:58
火花(2019年12期)2019-12-26 01:00:28
人大建設(shè)(2019年12期)2019-05-21 02:55:32
學(xué)苑創(chuàng)造·A版(2015年11期)2016-01-14 09:03:27
中國合理用藥探索(2014年11期)2014-03-11 20:30:20