溫度和鹽度對大竹蟶孵化及幼蟲生長變態(tài)的影響
2016-08-30 05:49:58郝詠芳曹琛邢佐平黃金勇匡少華宋松偉
水產(chǎn)養(yǎng)殖 2016年8期
郝詠芳,曹琛,邢佐平,黃金勇,匡少華,宋松偉
(1.營口市水產(chǎn)科學研究所,遼寧 營口 115004;2.大連海洋大學,遼寧 大連 116023)
溫度和鹽度對大竹蟶孵化及幼蟲生長變態(tài)的影響
郝詠芳1,2,曹琛1,邢佐平1,黃金勇1,匡少華1,宋松偉1
(1.營口市水產(chǎn)科學研究所,遼寧 營口 115004;2.大連海洋大學,遼寧 大連116023)
大竹蟶(Solen grandis Dunker)隸屬瓣鰓綱,異齒亞綱,簾蛤目,竹蟶科,竹蟶屬。貝殼較長,呈竹筒狀,一般殼長約為殼高的4~5倍,足極發(fā)達。營埋棲生活,生活在潮下帶至水下20 m水深處,生存底質(zhì)為沙底、泥沙底,主食浮游性較弱易于下沉或底棲的硅藻類[1]。大竹蟶廣泛分布于中國、朝鮮半島、日本以及菲律賓沿海,其肉質(zhì)營養(yǎng)價值高,蛋白質(zhì)含量高,必需氨基酸種類齊全,呈味氨基酸比例高,脂肪中含有大量的EPA及DHA等不飽和脂肪酸[2-3]。
目前,有關(guān)大竹蟶的研究主要集中在繁殖生物學[4-6]、人工育苗[7-10]等方面,培育密度及餌料種類[11]、環(huán)境因子[12-13]、鹽度等對大竹蟶稚貝生長發(fā)育影響的研究[14]也有報道,但溫度及鹽度對大竹蟶卵的孵化及幼蟲變態(tài)影響的相關(guān)研究尚未發(fā)現(xiàn)。筆者進行這方面的試驗,為大竹蟶人工育苗幼體孵化及變態(tài)階段積累基礎(chǔ)數(shù)據(jù),以期為其苗種的規(guī)模化培育技術(shù)示范及推廣提供理論依據(jù)。
1 材料與方法
1.1材料
1.1.1試驗場地及主要設(shè)施場地位于營口現(xiàn)代漁業(yè)科技產(chǎn)業(yè)園貝類研發(fā)車間。天然海水沉淀、砂濾后使用,鹽度22,pH值7.5~8.6,水溫17~18℃,水質(zhì)符合我國漁業(yè)水質(zhì)標準[3]。試驗用容器為35 cm×50 cm×15 cm的白塑料槽。
1.1.2試驗用大竹蟶受精卵及D型幼體大竹蟶種貝購自營口市白沙灣海區(qū),為2~3齡成熟大竹蟶親貝,個體健壯、生長旺盛、無損傷、肥滿度高、殼長8 cm以上。種貝暫養(yǎng)至性腺成熟自然產(chǎn)卵,300目篩絹網(wǎng)收集同一批受精卵及250目篩絹網(wǎng)選育出的同一批D型幼體。
1.2方法
1.2.1溫度對幼蟲孵化及生長變態(tài)的影響試驗海水鹽度22,水溫在15~36℃范圍內(nèi),3℃為一個梯度,設(shè)置8個組。以種蟶產(chǎn)卵時的水溫為基點,按每10 min升高或降低2℃的速率通過冰袋降溫或加熱棒升溫調(diào)節(jié),控溫儀控制,溫差不超過±0.1℃。1.2.2鹽度對幼蟲孵化及生長變態(tài)的影響試驗海水水溫23~24℃,鹽度6~38,以4為梯度,共設(shè)9個鹽度組。試驗用水由鹽度100的濃縮海水、鹽度22的天然海水、充分曝氣的自來水按比例配制而成,用YSI水質(zhì)快速分析儀標定。不同試驗組海水一次配制,分裝于20 L純凈水塑料桶中保鮮膜封口備用。以突變方式調(diào)節(jié)各組鹽度,鹽度對變態(tài)率影響的試驗時間為28 d。
1.2.3日常管理2014—2015年生產(chǎn)和試驗同步進行。每個試驗組設(shè)置兩個平行,各組微量充氣,保證溶氧,自然光照1 000 lx以內(nèi)。孵化試驗受精卵密度4個/mL,變態(tài)試驗D型幼體密度2個/mL。試驗在裝有2 L不同梯度水溫、鹽度海水的塑料槽中進行。定時取樣觀察、鏡檢、記錄幼蟲的發(fā)育情況及各階段形態(tài)變化和幼蟲數(shù)量。
孵化率試驗時間較短,要定時觀察記錄。變態(tài)率試驗中,日換水1次,換水量70%;日投餌2次,初期金藻(Isochrysis zhanjiangensis)開口,后與角毛藻(Chaetoceros sp)搭配投喂,前期投喂密度2 000 cell/mL,隨著各組幼體生長發(fā)育和數(shù)量的變化情況適當調(diào)整投喂量,以保持水體單胞藻細胞密度2 000 cell/mL以上,從而保證其足量攝食。鏡檢幼蟲存活、變態(tài)情況,測量殼長,及時撈出死亡幼體(死亡的界定以貝殼張開為標準)。試驗結(jié)束時,統(tǒng)計各組幼體孵化率、變態(tài)率和殼長日增長率。
1.3數(shù)據(jù)統(tǒng)計及處理
孵化試驗:受精卵孵化為D形幼蟲所需時間為孵化時間,凡D形幼蟲出現(xiàn)殼變形扭曲、左右殼不對稱、鉸合線彎曲、殼緣有缺刻、面盤或纖毛異常者均計為畸形個體,不計入孵化率。
變態(tài)試驗:大竹蟶的個別D型幼體殼長230 μm時即開始變態(tài),發(fā)育至眼點圓大、面盤完全脫落、足形成可爬行時視為完成變態(tài),此時幼體殼長大多在250~300 μm之間,此階段所需時間為變態(tài)時間,面盤及外殼異常者記為畸形個體,不算入變態(tài)率。計算公式如下:
孵化率:Rh=dh1/dh0×100%
變態(tài)率:Rs=ds1/ds0×100%
殼長平均日增長量:Qg=(L1-L0)/(t1-t0)
式中:dh1為D型幼體數(shù)量,dh0為受精卵數(shù)量;ds1和ds0分別表示截止試驗結(jié)束成功變態(tài)幼體密度和初始D型幼蟲數(shù)量;L1和L0分別為試驗結(jié)束時幼體殼長和初始D型幼蟲殼長(μm);t1和t0分別為試驗結(jié)束時間和D型幼體形成時間。試驗期間的死亡個體僅用于計算病死率指標。
試驗數(shù)據(jù)采用平均值(M)±標準差(SD)表示;Excel(2003)對數(shù)據(jù)進行處理、作圖,SPSS17.0對數(shù)據(jù)進行單因素方差分析(ANOVA),用于檢驗處理間的差異顯著性(P<0.05)。
2 結(jié)果與分析
2.1溫度對大竹蟶幼蟲孵化及生長變態(tài)的影響
溫度對大竹蟶幼蟲孵化及生長變態(tài)的影響見圖1、圖2。
試驗結(jié)果表明,水溫對于大竹蟶受精卵孵化率影響具有統(tǒng)計學意義(P<0.05)。大竹蟶受精卵對低溫比對高溫適應(yīng)性略強。水溫12℃,幼體分裂至多細胞即停止發(fā)育或畸形,無法完成孵化;15℃時,受精卵胚胎發(fā)育進程緩慢,大部分發(fā)育畸形,只有28%的受精卵可以正常孵化,且所需時間較長;15~24℃時,隨著水溫升高,受精卵孵化至D型幼蟲時間縮短,孵化率顯著升高(P<0.05);水溫24℃時,孵化率最高為79.25%,略高于27℃的72.75%,變態(tài)所需時間也最短,僅為5 d;27℃起,水溫上升孵化時間逐步增長,孵化率降低顯著(P<0.05);33℃時受精卵大部分發(fā)育至多細胞便開始解體,少數(shù)畸形,孵化率僅15.25%,但孵化時間和18℃相同,均略低于15℃;36℃時極個別幼蟲能發(fā)育至擔輪幼蟲,但也無法正常孵化。因此,大竹蟶受精卵孵化的適宜溫度范圍18~30℃,最適溫度范圍21~27℃,在此范圍內(nèi),幼蟲孵化較快且孵化率較高。因此,大竹蟶受精卵孵化過程中,應(yīng)將水溫控制在21~27℃。
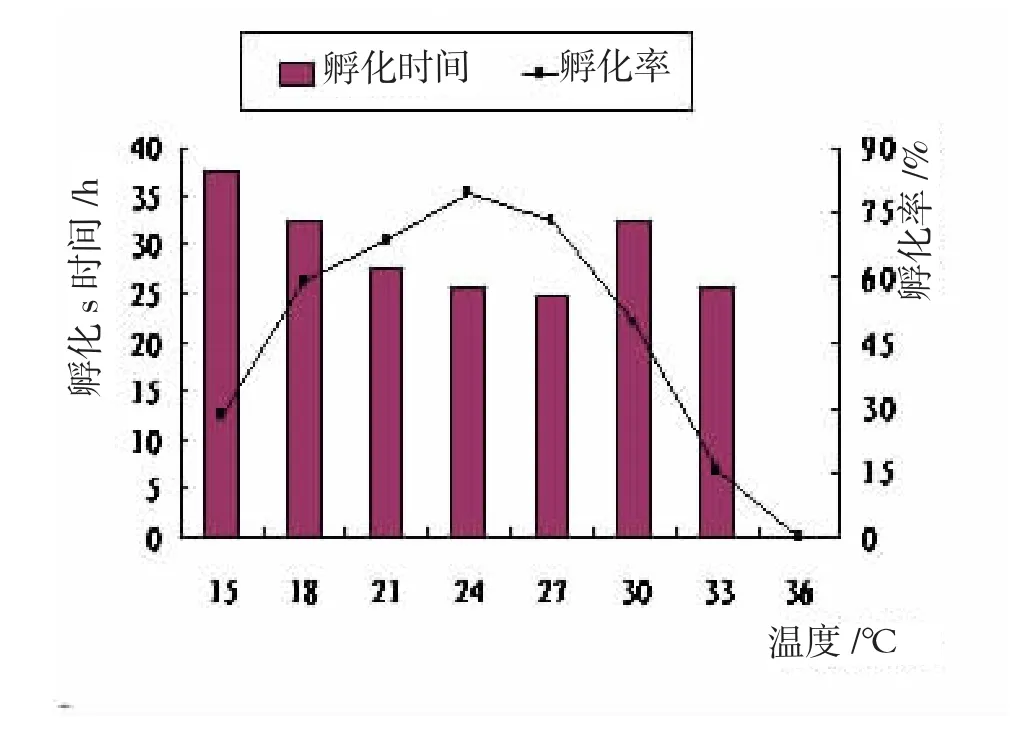
圖1 溫度對幼體孵化的影響
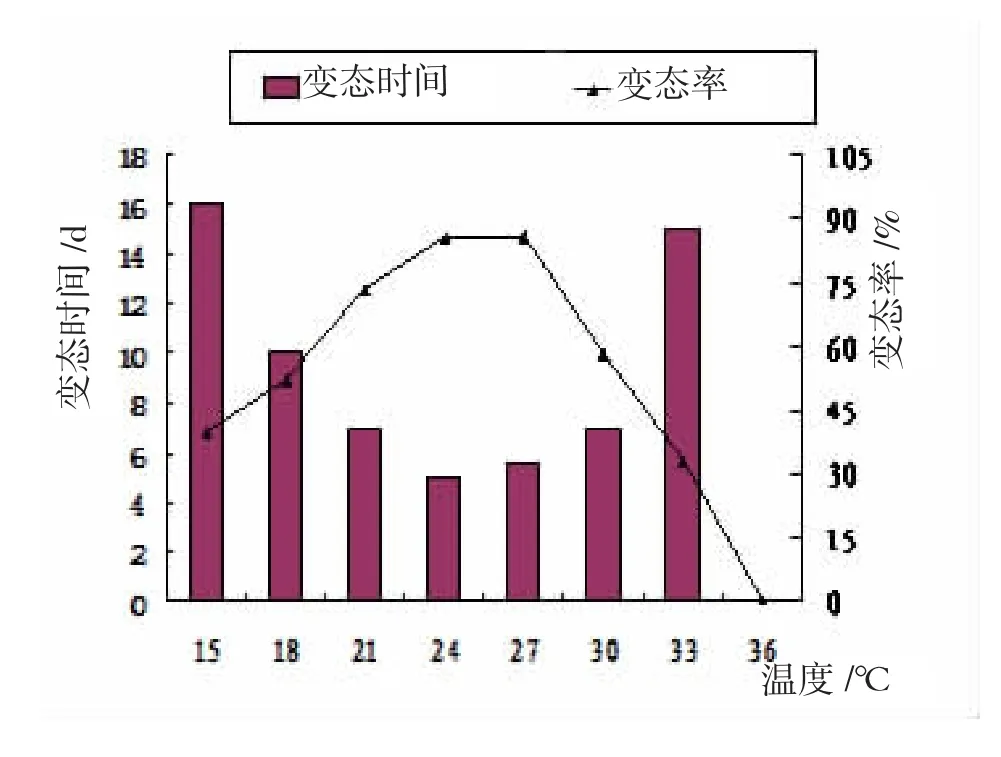
圖2 溫度對幼體變態(tài)的影響
水溫對大竹蟶幼體變態(tài)率的影響顯著(P<0.05),12℃無法完成變態(tài),少數(shù)發(fā)育至殼頂期即死亡;15℃時D型幼體活力弱,攝食差,部分幼體面盤伸縮異常,48 h開始出現(xiàn)面盤解體,72 h后有死亡個體,變態(tài)時間較長,變態(tài)率僅39.75%;15~24℃時,隨著溫度上升,D型幼體殼長日增長增大極顯著,變態(tài)率顯著上升,24℃時殼長日增長23.4 μm/ d,變態(tài)率85.45%;27℃殼長日增長、變態(tài)時間及變態(tài)率與24℃差異均無統(tǒng)計學意義(P>0.05);水溫30℃時,12 h內(nèi)幼蟲表現(xiàn)異常活潑,面盤伸縮頻繁,濾食能力強,但此后逐漸出現(xiàn)沉底、纖毛脫落等現(xiàn)象,殼長日增長量下降顯著(P<0.05);36℃,12 h后幼蟲空胃率70%以上,幾乎停止發(fā)育,面盤解體、逐漸沉底,無法完成變態(tài)。因此,大竹蟶D型幼體變態(tài)的適宜溫度范圍18~30℃,最適溫度范圍21~27℃。
2.2鹽度對大竹蟶幼蟲孵化及生長變態(tài)的影響
如圖3所示,水溫23~24℃時,鹽度對于大竹蟶受精卵的孵化率及發(fā)育速度均影響顯著(P<0.05)。鹽度過高或過低,受精卵無法正常孵化,鹽度為6時大竹蟶受精卵分裂至原腸期終止,鹽度38時受精卵也無法完成孵化;鹽度10時,大部分幼蟲發(fā)育至擔輪幼體即終止,孵化率僅2.75%,且孵化時間較長;鹽度10~34,各組均有D型幼蟲孵出,但孵化率差異具有統(tǒng)計學意義(P<0.05);鹽度10~26時,隨著鹽度升高孵化率顯著升高(P<0.05),孵化時間顯著降低(P<0.05);鹽度26孵化率最大為82.25%,且孵化時間最短,鹽度22次之;鹽度26~34,鹽度升高孵化率顯著下降(P<0.05),孵化時間顯著增長(P<0.05);鹽度34時受精卵多數(shù)發(fā)育至多細胞,分裂球分裂畸形,最終死亡,孵化率僅13.85%。由此可見,大竹蟶受精卵孵化適宜鹽度范圍為18~30,最適范圍為22~30,在此范圍內(nèi),大竹蟶的孵化率較高,發(fā)育速度較快,初孵D形幼蟲活力較好。
由圖4可知,鹽度變化對大竹蟶幼體的變態(tài)率影響顯著(P<0.05)。鹽度過高、過低,大竹蟶幼蟲無法正常變態(tài)。鹽度6時,D型幼體發(fā)育12 h后陸續(xù)閉殼沉底,鏡檢發(fā)現(xiàn)幼蟲面盤纖毛和胃部仍在活動,60 h后全部死亡;鹽度10時變態(tài)所需時間較長,變態(tài)率僅3.5%;鹽度14~26,隨著鹽度升高幼體變態(tài)率顯著增大(P<0.05),攝食、活力較強,生長速度也逐漸提高;鹽度26時殼長平均日增長量和變態(tài)率最大,變態(tài)時間最少,變態(tài)規(guī)格相對較小(殼長231 μm),鹽度22時次之;鹽度26~34,變態(tài)率隨鹽度上升顯著下降(P<0.05),生長發(fā)育減慢,變態(tài)時間增長;鹽度38時,幼蟲高滲反應(yīng)明顯,游動異常,24 h出現(xiàn)死亡,3 d后病死率40%,存活的幼蟲以殼頂中期為主,6 d后全部死亡;各鹽度組對大竹蟶幼體變態(tài)率的影響依次為:鹽度26>鹽度22>鹽度30>鹽度18>鹽度34>鹽度14>鹽度10。因此,大竹蟶D型幼體變態(tài)的適宜鹽度范圍為18~30,最適鹽度范圍22~30。

圖3 鹽度對幼體孵化的影響

圖4 鹽度對幼體變態(tài)的影響
3 討論
3.1溫度大竹蟶幼蟲孵化及變態(tài)的影響
貝類屬于變溫動物,其體溫及代謝隨環(huán)境溫度的改變而變化。溫度作為影響貝類生長發(fā)育最重要的環(huán)境因子之一,在一定范圍內(nèi)緩慢提高可以促進貝類的生長發(fā)育。本研究表明,大竹蟶受精卵孵化和變態(tài)的適宜溫度為18~30℃。低溫時胚胎發(fā)育緩慢且畸形率高,無法正常孵化或順利完成變態(tài)。最適溫度為21~27℃,在此范圍內(nèi),胚胎發(fā)育正常且發(fā)育較快,孵化率和變態(tài)率較高。這與陳愛華[12]的大竹蟶稚貝存活和生長的適宜溫度范圍6~30℃,最適溫度范圍21~30℃略有不同,可能是由于大竹蟶受精卵和浮游幼蟲較稚貝胚胎發(fā)育不完善,對低溫抵抗力差有關(guān)。與青蛤胚胎、浮游幼蟲的適宜溫度24~32℃,最適生長溫度26~30℃[15]不同,與毛蚶孵化的適宜溫度25~30℃[16]、幼蟲生長最適溫度30℃也不同,這可能與它們的生活習性有關(guān),大竹蟶在遼東灣一帶5月下旬至6月為繁殖盛期,此時天然海水溫度范圍在18~22℃;而青蛤繁殖盛期是每年的7月至9月,正處于盛夏季節(jié)[15],海水溫度較高;毛蚶的產(chǎn)卵大多在6月底至7月份,海水溫度也較高。因此大竹蟶幼體的最適孵化及變態(tài)溫度范圍應(yīng)該在21~27℃。
3.2鹽度對大竹蟶幼蟲孵化及變態(tài)的影響
海洋貝類屬于變滲動物,能夠根據(jù)周圍環(huán)境海水鹽度不同而調(diào)節(jié)其滲透壓,以利于攝食和生長[12]。貝類對鹽度的耐受力與滲透調(diào)節(jié)能力、營養(yǎng)狀況和遺傳變異(包括抗逆基因的表達)有關(guān),其中滲透調(diào)節(jié)能力直接決定了貝類適合生長的環(huán)境[17]。尤仲杰等[18]研究發(fā)現(xiàn),墨西哥灣扇貝幼蟲和稚貝在不適的鹽度下,其圍心腔壓力增大,心跳減慢,對外界的刺激反應(yīng)遲鈍等,處于這種狀態(tài)下的幼蟲抵抗力下降,代謝緩慢或停止,易死亡。
本研究表明:大竹蟶受精卵孵化及變態(tài)的適宜鹽度為18~30,最適鹽度范圍22~30。這與文蛤孵化期最適鹽度17.3~28.4[19]相近,與文蛤浮游幼體的最適鹽度15.9~22.6[19]不同。由于大竹蟶分布在潮下帶至水下20 m水深的淺海,鹽度較近海高,故其對低鹽的耐受能力較差;而文蛤長期生活在有淡水注入的河口附近,自然進化成對鹽度的廣泛適應(yīng)性,并表現(xiàn)出對低鹽的適應(yīng)性強于高鹽[14]。本結(jié)果與陳愛華[12]對大竹蟶稚貝的研究一致;與青蛤20~25為最適鹽度(變態(tài)率在84.1%~88.0%)[15]相近,與毛蚶受精卵孵化最適鹽度22~26[16],毛蚶幼蟲生長最適鹽度20~25相近。這也表示,大竹蟶受精卵及浮游幼蟲與大多數(shù)海洋雙殼貝類相同,對鹽度要求較高,在適鹽范圍內(nèi)生長發(fā)育較快。
3.3耐鹽耐溫實踐指導意義
海水溫度和鹽度是影響海洋生物存活和生長的重要環(huán)境因子,當二者中的一個因子控制在安全范圍,通過單因素實驗?zāi)軌蚍从成飳囟然螓}度的適應(yīng)性[20],這也是本試驗研究的主要內(nèi)容。貝類對溫度和鹽度的適應(yīng)能力與種類的遺傳性和個體的生理狀態(tài)有關(guān),而與其個體大小關(guān)系不明顯,同種類的所有大小的個體都以相同方式對溫度和鹽度作出反應(yīng)[21],這與本研究的結(jié)果一致。許多學者[6-7]認為生物體對環(huán)境的適應(yīng)能力受許多環(huán)境因子的共同影響,其中溫度和鹽度作為養(yǎng)殖環(huán)境的主要因子,對雙殼貝類的生長、存活和繁殖有顯著影響[22-24]。有關(guān)研究表明,溫度和鹽度不但影響貝類的胚胎發(fā)育速度和孵化率,也影響所孵化幼蟲的健康狀態(tài)。當溫度或鹽度超過貝類胚胎發(fā)育的忍受范圍時,將引起孵化率明顯下降,部分能發(fā)育到幼蟲的,其畸形率也明顯增大[25-26]。
孵化率及變態(tài)率的高低對苗種繁育影響重大,故其幼體孵化及變態(tài)的適宜溫度和鹽度范圍在其人工苗種生產(chǎn)中具有十分重要的指導意義。建議在大竹蟶苗種繁育的過程中,將溫度控制在21~27℃內(nèi),鹽度范圍22~30內(nèi)為宜。
參考文獻:
[1]潘星光.縊蟶的生態(tài)觀察與食性分析 [J].動物學雜志,1959,3(8):355-357.
[2]戴中杰.大竹蟶軟體部營養(yǎng)成分分析及評價[J].集美大學學報,2002,7(4):304-308.
[3]戴中杰.大竹蟶軟體部的氨基酸分析[J].莆田學院學報,2002,9(3):32-35.
[4]肖國強,柴雪良,邵艷卿,等.大竹蟶的繁殖生物學[J].海洋科學,2009,33(10):21-25.
[5]陳愛華,姚國興,吳楊平,等.呂四漁場海區(qū)大竹蟶的繁育與生長[J].海洋漁業(yè),2012,34(3):277-284.
[6]吳楊平,陳愛華,姚國興,等.大竹蟶生物學特性的初步研究[J].水產(chǎn)科學,2011,30(12):764-767.
[7]曹琛,閆喜武,李曉冬,等.大竹蟶生產(chǎn)性人工育苗技術(shù)研究[J].齊魯漁業(yè),2012(6):4-7.
[8]王雪梅,路宜華,豐愛秀,等.大竹蟶健康苗種培育新模式的研究[J].水產(chǎn)養(yǎng)殖,2012(8):14-16.
[9]侯和要,牟乃海,宋全山,等.大竹蟶人工繁育技術(shù)研究[J].齊魯漁業(yè),2004,21(6):32-35.
[10]陳愛華,姚國興,張志偉.大竹蟶生產(chǎn)性人工繁育實驗[J].海洋漁業(yè),2009,31(1):66-72.
[11]閆喜武,趙生旭,張澎,等.培育密度及餌料種類對大竹蟶幼蟲生長、存活及變態(tài)的影響 [J].大連海洋大學學報,2010,25(5):386-390.
[12]陳愛華,張志偉,姚國興,等.環(huán)境因子對大竹蟶稚貝生長及存活的影響 [J].上海水產(chǎn)大學學報,2008,17(9):559-563.
[13]陳愛華,姚國興,張志偉,等.溫度、鹽度和底質(zhì)對大竹蟶稚貝生長及存活的影響 [J].熱帶海洋學報,2010(5):94-97.
[14]侯和要,王君霞,彭作波,等.不同鹽度對大竹蟶存活的影響[J].齊魯漁業(yè),2004,21(5):5-6.
[15]王丹,徐善良,尤仲杰,等.溫度和鹽度對青蛤孵化及幼蟲、稚貝存活與生長變態(tài)的影響[J].水生生物學報,2005,29 (5):495-501.
[16]沈偉良,尤中杰,施祥元.溫度與鹽度對毛蚶受精卵孵化及幼蟲生長的影響[J].海洋科學,2009,33(10):5-8.
[17]薛凌展,闕華勇,張國范,等.鹽度對近江牡蠣幼蟲生長及存活的影響[J].海洋科學,2007,31(9):73-76.
[18]馬洪明,劉曉偉,麥康森,等.鹽度突降對櫛孔扇貝(Chlamys farreri)抗病力指標的影響[J].高技術(shù)通訊,2006,16 (7):746-751.
[19]陳沖,王志松,隨錫林.鹽度對文蛤孵化及幼體存活和生長的影響[J].海洋科學,1999(3):16-18.
[20]Castagna M.Culture of the bayscallop,Argopecten irradinans Lamark,in Vinrginia[J].Marine Fisheries Review,1975 (37):19-24.
[21]何義朝,張福綏.墨西哥灣扇貝稚貝對鹽度的耐受力[J].海洋學報,1999,21(4):87-91.
[22]林筆水,吳天明.鹽度和溫度同縊蟶稚貝存活于生長的關(guān)系[J].水產(chǎn)學報,1986,10(1):41-45
[23]尤仲杰,徐善良,邊平江,等.海水溫度和鹽度對泥蚶幼蟲和稚貝生長及存活的影響 [J].海洋學報,2001,23(6):108-113.
[24]劉志剛,王輝,栗志民,等.墨西哥灣扇貝高起始致死溫度的研究[J].中國水產(chǎn)科學,2007,14(5):778-785.
[25]何義朝,張福綏.鹽度對海灣扇貝不同發(fā)育階段的影響[J].海洋與湖沼,1990,21(3):197-203.
[26]何義朝,張福綏.溫度對貽貝胚胎發(fā)育的影響[A].貝類學論文集(第一集)[C].北京:科學出版社,1983:133-142.
資助項目:農(nóng)業(yè)科技成果轉(zhuǎn)化資金項目(2014GB000363)
doi:10.3969/j.issn.1004-2091.2016.08.006
收稿日期:(2016-01-28)
猜你喜歡
中學生數(shù)理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
媽媽寶寶(2017年3期)2017-02-21 01:22:28
紅領(lǐng)巾·萌芽(2016年1期)2016-09-10 07:22:44
中國塑料(2016年3期)2016-06-15 20:30:00
