擴展青霉拮抗菌的選育及抑菌機制初探
2016-09-10 06:15:32付瑞敏邢文會張麗琴陳五嶺
食品工業科技 2016年5期
付瑞敏,邢文會,張 紅,張麗琴,陳五嶺
(1.河南教育學院,河南鄭州 450046;2.西北大學 生命科學學院,陜西西安 710069)
?
擴展青霉拮抗菌的選育及抑菌機制初探
付瑞敏1,2,邢文會1,張紅1,張麗琴1,陳五嶺2,*
(1.河南教育學院,河南鄭州 450046;2.西北大學 生命科學學院,陜西西安 710069)
為選育有效抑制擴展青霉(Penicilliumexpansum)的拮抗菌,并初步探討其抑菌機制。從蘋果表面分離到拮抗擴展青霉的菌株BA-16,經形態學、生理生化及16S rRNA基因序列分析,對該菌進行鑒定,并采用低能N+注入技術對其進行誘變選育。采用雙酶反應體系檢測野生株和突變株對擴展青霉分泌磷脂酶的抑制效果以檢測突變效果并探究其抑菌機制。經鑒定,BA-16-8該菌被鑒定為解淀粉芽孢桿菌(Bacillusamyloliquefaciens)。低能N+注入技術誘變選育出的突變株BA-16-8抑菌性能顯著提高且遺傳性能穩定。磷脂酶活性結果顯示,相對于野生株,突變株代謝產物可顯著抑制病原菌所分泌的磷脂酶A的活性,且其抑制效果隨濃度的增高而增強,故推測拮抗菌可能通過該機制起到抑制擴展青霉的作用。本研究對于蘋果采后青霉病的生物防治具有良好應用開發前景。
擴展青霉,解淀粉芽胞桿菌,低能N+注入,磷脂酶A
蘋果采摘后病害的感染會造成果實腐爛嚴重,給蘋果種植業及副產品加工行業帶來巨大的經濟損失。其中,青霉病是蘋果采摘后致其腐爛的重要病害之一,該病的有效防治成為蘋果產業亟待解決的問題。研究表明,引起青霉病的主要致病菌是擴展青霉(Penicilliumexpansum)[1]。目前控制擴展青霉的主要方式仍是以化學殺菌劑和冷藏等物理手段為主,化學殺菌劑雖然能夠有效控制病原菌擴展青霉的生長,但是隨著病原菌抗藥性能和人們健康環保意識的不斷增強,化學防治方法已逐漸不被人們認可。諸如冷藏等物理方法雖然一定程度上可以阻止病原菌的生長但容易影響水果的風味和營養價值,且處理不當會造成凍害,此外,處理成本也相對較高。而采用拮抗性生物抑制病原菌生長從而達到水果防腐和保鮮的生物防治方法作為水果采摘后病害防治的新方法,不僅不會改變水果的營養和風味,還可以消除由于化學藥劑對人體健康所造成的隱患,故近幾年已成為水果防腐保鮮的研究熱點。
近年來已經有不少關于蘋果青霉病的生物防治的研究報道,采用的生物防治菌主要集中于生防酵母。Li 等人[2]研究了粘質紅酵母防治蘋果采后灰霉病和青霉病的機制,發現該酵母通過誘導蘋果中過氧化物酶和多酚氧化酶的活性和高度抑制蘋果脂質過氧化作用從而誘導蘋果中防御機制依賴性酶的活化,此外,該酵母通過和病原菌的空間和營養競爭抑制病原菌生長。后來,人們陸續發現芽孢桿菌屬的一些菌株可以抑制擴展青霉的生長。Lee[3]等人從鹽生植物根際分離出枯草芽孢桿菌S16并研究了該菌對采摘后蘋果和水蜜桃青霉病的生物防治作用,發現該菌可有效抑制擴展青霉的生長。磷脂酶A作為病原性真菌的致病因子,對于真菌在植物表面的定殖和入侵有重要輔助作用,有報道說明芽孢桿菌的代謝產物如豐原素等脂肽類產物可對磷脂酶A的活性產生影響[4]。
本實驗室從蘋果表面分離篩選到一株對擴展青霉具有拮抗作用的菌株BA-16,結合該菌株的形態特征、生理生化和分子系統發育等特征分析結果對其進行鑒定,并采用低能N+注入技術對其進行誘變,以期獲得可有效拮抗青霉病的菌株,為日后將其應用于采摘后蘋果青霉病的生物防治奠定理論基礎。
1 材料與方法
1.1材料與儀器
致病菌擴展青霉(Penicilliumexpansum)陜西省微生物研究所惠贈,供試蘋果采收于陜西洛川縣。
N+離子注入機BNU-400中科院北京市輻射中心研制,N+離子能量為25 KeV,束流0.4 mA,真空度為5×10-3Pa。
1.2培養基
牛肉膏蛋白胨培養基(YEB)用于拮抗菌的培養[5]:牛肉膏3 g,蛋白胨10 g,NaCl 5 g,瓊脂15~20 g,水1000 mL,pH7.4~76.
PDA(馬鈴薯培養基)用于病原性真菌的培養[5]:馬鈴薯200 g,葡萄糖20 g,瓊脂15~20 g,蒸餾水1000 mL,pH7.2.
搖瓶發酵培養基用于發酵拮抗菌胞外抗菌性脂肽蛋白[6]:葡萄糖20 g,L-谷氨酸鈉5 g,MgSO40.5 g,KCl 0.5 g,KH2PO41 g,FeSO40.15 g,MnSO45 g,CuSO40.16 mg,蒸餾水1000 mL,pH7.2.
1.3細菌的分離、篩選
從感染青霉病蘋果的果面分離菌株。用無菌水清洗蘋果健康部位,將所得液體經過10-1~10-6的梯度稀釋,將10-4、10-5、10-6梯度稀釋液分別涂布在YEB平板上,37 ℃培養24 h。挑取各平板上的菌株分別進行純化,得到單克隆,將其分別編號以作進一步研究。
采用平板對峙法[7]對分離菌株進行拮抗能力的篩選。以擴展青霉為指示菌,將待測菌株和病原菌接種于PDA平板上,病原菌接種于平板中央,待測菌株同病原菌間的距離為2.5 cm 28 ℃培養48 h觀察其生長狀況,并測量抑菌帶寬度,實驗均重復三次,挑取抑菌條帶寬的菌株至YEB平板上,保存備用。
將初篩所得菌株接種于液體發酵培養基中,搖瓶震蕩培養72 h(37 ℃,150 r/min)。參照韓欣宇等的方法[8]制備菌株無細胞發酵液,將所制的發酵液200 μL接入牛津杯中,采用牛津杯法[9]檢測菌株無細胞發酵液拮抗擴展青霉的能力。28 ℃培養24 h后,測量記錄牛津杯周圍出現的抑菌圈直徑,挑選直徑最大的菌株做進一步研究。整個實驗以無菌水作為對照,實驗均重復三次。
1.4菌株的形態學與生理生化鑒定
將復篩所選菌株經染色、光學顯微鏡觀察菌體個體及群體形態。參照《微生物學實驗指導》[10]和《伯杰細菌鑒定手冊》[11]就菌株的生理生化特征進行初步鑒定。
1.516S rRNA基因序列分析
提取菌株的總基因組DNA為模板[12],利用細菌16S rRNA基因的通用引物27F(5′-AGAGTTTGATCATGGCTCAG-3′)和1492R(5′-CTACGGTTACCTT GTTACGAC-3′)進行PCR擴增[12]。PCR反應體系25 μL,熱循環參數如下:94 ℃預變性3 min,94 ℃變性1 min,55 ℃退火1 min,72 ℃延伸2 min,循環30次,72 ℃延伸10 min。所得PCR產物經回收純化后提交至上海生工進行測序,測序結果與NCBI數據庫中相關菌種的16S rRNA基因序列用BLAST程序進行比對分析,利用MEGA4.1構建系統發育樹[13]。
1.6高效拮抗菌的低能N+注入誘變選育
1.6.1樣品處理以復篩所得的菌株為出發菌株,將其轉接至新鮮斜面并培養24 h后,用5 mL的生理鹽水清洗斜面菌苔,并將其倒入含有玻璃珠的錐形瓶內,充分振蕩后,采用分光光度計法測量其600 nm處的光吸收值,調整菌懸液濃度至OD600=0.986,此時菌懸液濃度為108cfu/mL。用微量移液器吸取0.1 mL菌懸液將其接種于90 mm的無菌平皿中,無菌條件下涂布均勻并風干制成菌膜[14]。
1.6.2離子注入將待處理平皿置于離子注入室,打開皿蓋,室內抽真空,注入能量為25 KeV的低能,N+離子束,注入劑量分別為1×1015、1.5×1015、2.0×1015、2.5×1015、3.0×1015ions/cm2。
1.6.3突變株的篩選用無菌生理鹽水1 mL沖洗離子注入后的平板,將所得液體進行梯度稀釋到10-5、10-6和10-7,分別取各稀釋的菌懸液0.1 mL將其接種于YEB平板上,37 ℃培養48 h,觀察所得突變株的菌落形態并進行菌落計數。
1.6.4計算存活率與正變率將突變株的發酵液接種于PDA平板上,采用牛津杯法檢測各突變株抗擴展青霉能力,規定抑菌圈直徑大于出發菌株的為正變株,參照下列公式計算存活率與正變率:
存活率(%)=誘變后的活菌數/誘變前的活菌×100
正變率(%)=正突變的菌株數/誘變后的活菌×100
1.7正變株的篩選及遺傳穩定性檢測
選取抑菌圈直徑較大的突變株,將其轉接至YEB平板上,37 ℃培養24 h,作為第一代,后每隔24 h傳一代,每代均隨機選取3個克隆采用牛津杯法檢測其無細胞發酵液抗擴展青霉的活性,進行遺傳穩定性檢測。
1.8擴展青霉菌絲分泌的磷脂酶的提取
將培養了48 h的病原菌擴展青霉培養液10000 g離心10 min,去除菌絲體,所得上清液用70%的飽和硫酸銨沉淀,而后10000×g離心5 min,取沉淀,用磷脂酶A提取液溶解,透析后所得產物即為磷脂酶A粗品。
1.9磷脂酶活性的測定
磷脂酶活力測定參照參考文獻[17],具體采用雙酶體系:1,2-二亞油酰基卵磷脂10 μL,Tris-Hcl 125 μL,大豆氧合酶5 μL,脫氧膽酸鈉溶液50 μL,磷脂酶A粗提液10 μL。雙酶反應的基本原理是:以1,2-二亞油酰基卵磷脂為第磷脂酶A的底物,反應后生成亞油酸,亞油酸由脂氧合酶催化發生氧化反應,生成氫過氧化物,其對應的紫外吸收光譜在234 nm有吸收峰,單位時間內(1 h),該波長下的吸光值每增加0.1即為1個酶活力單位。
1.10拮抗菌發酵液對病原菌分泌磷脂酶A的影響
將野生型BA-16和突變株BA-16-8的發酵24 h的菌株發酵液10000×g離心10 min,取上清,用超濾管過濾后即為無細胞發酵液,將所得無細胞發酵液分別做梯度稀釋,將不同稀釋度的無細胞發酵液分別定量加入雙酶反應體系中,各自反應1 h,通過測定各體系在234 nm處的吸光值,以不加無細胞發酵液的磷脂酶反應吸光值為100,通過所得值與其的比值來衡量各稀釋度無細胞發酵液作用下的磷脂酶活性。
2 結果與分析
2.1擴展青霉拮抗菌株的分離和篩選
經過初篩從蘋果表面分離獲得18株菌,其中酵母4株,細菌14株。通過平板對峙實驗,篩選出可拮抗擴展青霉的6株。采用牛津杯法檢測六株菌的無細胞發酵液抗擴展青霉活性,結果如表1所示。結果表明:六株菌的菌株細胞和無細胞發酵液拮抗擴展青霉的效果表現出了顯著的差異。其中,BA-16的拮抗效果最強,故將其挑選出來做后續研究。

表1 擴展青霉拮抗菌篩選結果Table 1 The screening results of antagonistic strains against Penicillium expansum
2.2擴展青霉拮抗菌株的形態學與生理生化鑒定
菌株BA-16經革蘭氏染色陽性,顯微鏡鏡檢為桿菌,有芽孢,有莢膜,具有運動性。該菌株在LB平板上形成乳白色菌落。菌落邊緣不整齊,干燥不透明,中間有凸起。生理生化結果見表2。
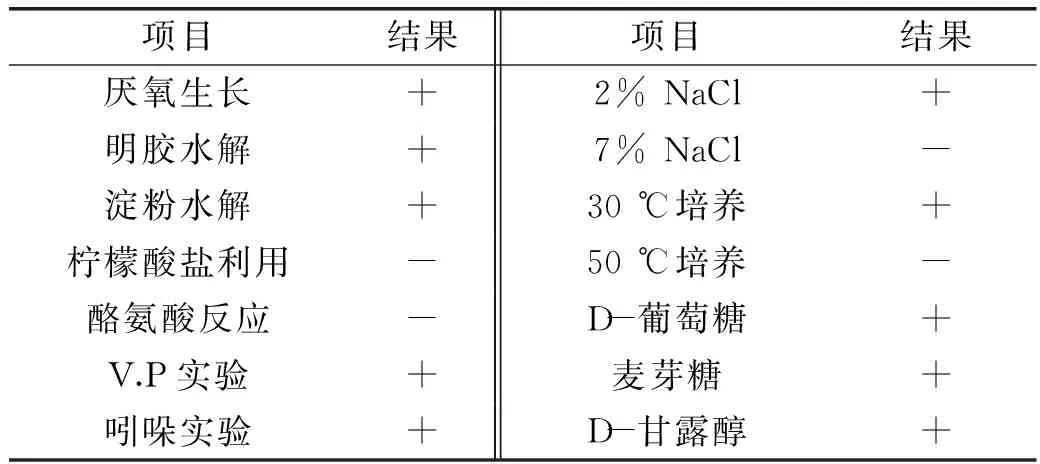
表2 菌株BA-16的生理生化特征Table 2 Characteristics of physiology and biochemistry of BA-16 strains
注:+/-表示在生理生化鑒定中陽性/陰性反應。+:陽性;-:陰性。
參照《常見細菌系統鑒定手冊》和《伯杰氏細菌鑒定手冊》,結合菌株BA-16的形態、染色及生理生化特征,可將該菌株初步鑒定為解淀粉芽孢桿菌(Bacillusamyloliquefaciens)。
2.316S rRNA 基因序列分析
提取菌株BA-16的總DNA進行PCR擴增,凝膠電泳回收擴增序列,得到一段1468 bp長度的16S rRNA基因,所獲序列提交至GenBank,登錄號No.KR31430。將其同GenBank中相近種屬進行BLAST比對,對BLAST結果進行分析,通過MEGA 4.1軟件構建系統發育樹(圖1)。發現菌株BA-16與BacillusamyloliquefaciensAY603658的相似性達到99%,結合上述的生理生化鑒定結果,可將菌株BA-16最終鑒定為解淀粉芽孢桿菌(Bacillusamyloliquefaciens)。
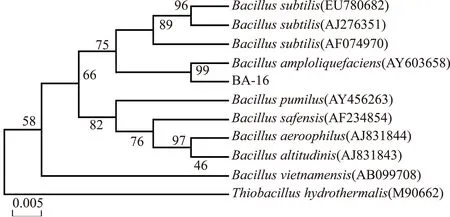
圖1 菌株BA-16 16SrRNA基因序列發育進化樹Fig.1 Phylogenetic tree of 16s rRNA gene sequences of isolated strain BA-16
2.4離子注入誘變結果
2.4.1最佳誘變劑量的確定采用低能N+注入誘變BA-16,野生株在該離子能量作用下發生突變,不同注入劑量對菌株的存活率和正變率影響如圖2所示。
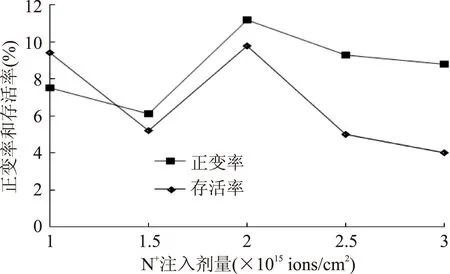
圖2 不同N+注入劑量對突變株的影響Fig.2 The effect of different does of N+ implantation on mutants
由圖中可以看出,一定范圍內,高劑量的離子注入可導致菌株正突變率的增加,同時也會導致菌株存活率的下降,一般認為,誘變菌株存活率在10%~20%,所得菌株正變率相對較高[15]。本實驗中,注入劑量在1×1015ions/cm2和2×1015ions/cm2時存活率接近該范圍,而后者的正變率遠高于前者,綜合考慮,以2×1015ions/cm2注入劑量為最佳誘變劑量。
研究發現,剛開始注入離子時,存活率急劇下降和正變率有所上升。推測其原因是帶能的氮離子最初作用于菌體細胞,可導致菌體細胞損傷從而導致存活率下降;同時,能量在胞內的積累也可導致菌體發生突變,故正變率有所上升。而當注入劑量到達一定值(1.5×1015ions/cm2)時,本來下降的存活率有了回升,而后又繼續下降。推測其原因可能是當離子注入劑量達到一定程度,可通過作用于菌體細胞DNA而激活其修復機制,因而一直下降的存活率會有少量回升。而當離子注入劑量進一步加大,所致損傷已無法修復,故存活率又進一步下降。
2.4.2BA-16突變株的篩選及遺傳穩定性分析采用不同劑量的N+注入共篩選出8 株抗擴展青霉高于野生株解淀粉芽孢桿菌BA-16的正突變株,分別對這8 株突變株進行平板對峙實驗和牛津杯實驗,結果如表3所示。選取抑菌帶寬和抑菌圈直徑在12以上的2 株突變型菌株:amyloliquefaciensBA-16-7和BacillusamyloliquefaciensBA-16-8進行傳代培養,并通過牛津杯法檢測各代拮抗病原菌的遺傳穩定性,結果如表4所示。
由表4可知,經過傳代,突變株BA-16-8抗菌性能差異不顯著,說明該突變株的遺傳穩定性較好,而BA-16-7的抗菌能力在不同世代差異顯著且呈現逐漸下降的趨勢,說明該突變株的遺傳穩定性較差,菌株退化較為嚴重。故選取突變株BA-16-8進行后續研究。
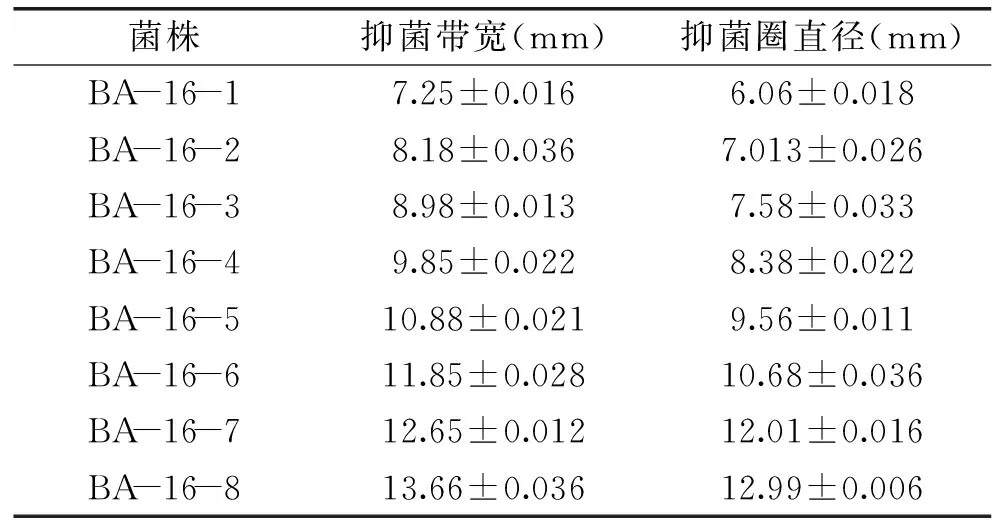
表3 突變株抗擴展青霉篩選結果Table 3 The screening results of mutants against Penicillium expansum
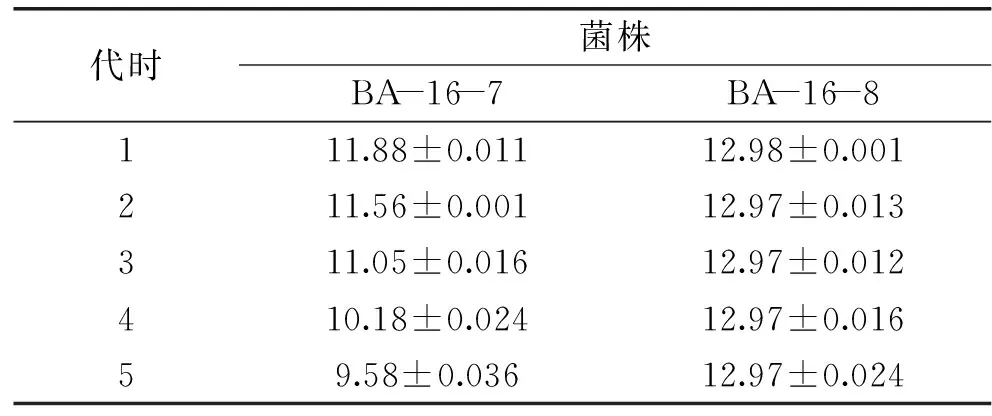
表4 突變株拮抗擴展青霉的遺傳穩定性分析結果Table 4 The Results of genetic stability analysis of mutants against Penicillium expansum
2.5磷脂酶A抑制實驗
將拮抗菌的野生株和突變株的無細胞發酵液分別以不同梯度稀釋度的濃度定量加入磷脂酶反應體系中,測定其酶活力比值,所得結果如圖3所示,隨著發酵液濃度的減少,所測得病原菌產磷脂酶A的活性直線上升,說明病原菌所產磷脂酶活受拮抗菌無細胞發酵液的抑制,且通過對比突變株和野生株的無細胞發酵液的抑制效果,發現突變株抑制效果顯著優于野生株,當用突變株的發酵液原液處理病原菌時,其磷脂酶A活性幾乎完全被抑制。
據報道,芽孢桿菌代謝產物如豐原素等可抑制磷脂酶A的活性,本研究中,發現拮抗菌代謝產物對病原菌所產磷脂酶A有抑制作用,且抑制作用隨濃度變化而變化,這一定程度上與前人報道結論是一致的[17-18],此外,由于磷脂酶A在病原菌侵染和定殖中起重要作用,因此,拮抗菌代謝產物對磷脂酶的抑制可緩解或阻止作物被病原菌感染的進程。此外,研究發現,與野生株相比,突變株對磷脂酶A的抑制作用要更明顯,這說明在誘變過程中突變株的代謝產物比如一些脂肽類抗生素的表達量有了增加,這種猜測將在進一步的研究中加以驗證。

圖3 不同濃度拮抗菌發酵液 對擴展青霉分泌的磷脂酶A活性影響 Fig.3 Effects of fermentation broth in different concentration on the activity of PLA secreted by Penicillium expansum
3 結論
本研究主要針對蘋果采摘后主要致病菌擴展青霉,從蘋果表面分離篩選出一株可有效拮抗擴展青霉的細菌BA-16,經鑒定,該菌為解淀粉芽孢桿菌(Bacillusamyloliquefaciens)為進一步提高菌株BA-16防治擴展青霉的能力,我們采用低能N+注入技術對其進行誘變。并研究了不同注入劑量對突變株存活率和正變率的影響。結果顯示,2×1015ions/cm2注入劑量為最佳誘變劑量。通過檢測突變株的菌體細胞和無細胞發酵液拮抗擴展青霉的能力,篩選出抗菌性能最強且遺傳性能穩定的突變株BA-16-8。將所得突變株和野生株的菌懸液和無細胞發酵液分別處理病原菌并檢測其對病原菌分泌磷脂酶A活性的影響,結果顯示,野生株和突變株均可不同程度地抑制病原菌所分泌磷脂酶的活性,其中突變株的抑制能力要明顯高于野生株,此外,隨著發酵液濃度的降低其抑制力也逐漸下降,說明,病原菌的代謝產物中有部分物質比如Fengycin可影響磷脂酶A的活性,本研究結果有待進一步實驗加以證實。
[1]Zhao L,Zhang H,Lin H,et al. Effect of trehalose on the biocontrol efficacy of pichia caribbica against post-harvest grey mould and blue mould decay of apples[J]. Pest management science,2013,69(1):983-989.
[2]Li R,Zhang H,Liu W,et al. Biocontrol of postharvest gray and blue mold decay of apples with rhodotorula mucilaginosa and possible mechanisms of action[J]. International Journal of Food Microbiology,2011,146(1):151-156.
[3]Hong P,Hao W,Luo J,et al. Combination of hot water,bacillus amyloliquefaciens hf-01 and sodium bicarbonate treatments to control postharvest decay of mandarin fruit[J]. Postharvest Biol Tec,2014,88(1):96-102.
[4]David HE,Tania CS. Hexadecylphosphocholine(Miltefosine)has broad-spectrum fungicidal activity and is efficacious in a mouse model of cryptococcosis[J]. Antimicrob Agents Chemother,
2006,50(2):414-421.
[5]Afsharmanesh H,Ahmadzadeh M,Javan-Nikkhah M,et al. Improvement in biocontrol activity of bacillus subtilis utb1 against aspergillus flavus using gamma-irradiation[J].Crop Protection,2014,60(2):83-92.
[6]姚磊,徐良雄,薛璟花,等. 枝頂孢屬真菌的抑菌活性及其代謝產物研究[J]. 熱帶亞熱帶植物學報,2012,20(1):192-196.
[7]劉雪,葉婧,穆長青,等. 枯草芽孢桿菌 b-332 菌株發酵條件的研究[J]. 農業科學與技術:英文版,2013,12(1):81-85.
[8]韓立榮,張華姣,高保衛,等. 放線菌 11-3-1 對油菜菌核病的防治作用與菌株鑒定[J]. 植物保護學報,2012,39(1):97-102.
[9]韓欣宇,陳志厚,羅定棋,等. Tpb55菌株發酵液中活性代謝產物的抑菌作用及穩定性測定[J]. 中國農學通報,2012,28(1):260-264.
[10]韋婉,李少英,王夢姣,等. 益生乳酸菌對氟喹諾酮類藥物的敏感性檢測方法的研究[J]. 食品科技,2013,10(1):22-26.
[11]黃文芳,張松. 微生物學實驗指導[M]. 暨南大學出版社:2003.
[12]布坎南,吉本斯. 伯杰細菌鑒定手冊,第八版[M],北京:科學出版社:1984.
[13]孫卓,楊利民. 人參病原菌拮抗細菌的分離篩選與鑒定[J]. 植物保護學報,2015,10(1):79-86.
[15]蔣益,鄭惠華,劉廣建,等. 低能 N+離子注入誘變選育云芝高產抗病菌株的研究[J]. 食用菌,2014,36(1):18-20.
[16]張瑞,周俊,王舒雅,等. 產 γ-聚谷氨酸菌株的誘變選育及其種子液工藝優化[J]. 生物加工過程,2015,13(1):47-53.
[17]Ivanovska N. Phospholipases as a factor of pathogenicity in microorganisms[J]. Journal of Molecular Catalysis B:Enzymatic,2003(22):357-361.
[18]Ganendren R,Widmer F,Singhal V,et al.Invitroantifungal activities of inhibitors of phospholipases from the fungal pathogen cryptococcus neoformans[J]. Antimicrob Agents Chemother,2004(48):1561-1569.
Breeding of antagonistic bacteria againstPenicilliumexpansumand study on its inhibition mechanism
FU Rui-min1,2,XING Wen-hui1,ZHANG Hong1,ZHANG Li-qin1,CHEN Wu-ling2,*
(1.Henan Institute of Education,Zhengzhou 450046,China;2.Northwest University,Xi’an 710069,China)
To breed the antagonistic strain which can inhibitPenicilliumexpansumeffectively,and discuss its antibacterial mechanism preliminarily. An antagonistic strain BA-16,which could inhibitPenicilliumexpansum,was isolated from apple surface and identified based on phenotypic,physiological,biochemical and phylogenetic(16S rDNA)studies. In order to enhance its antagonistic capability,the mutation of BA-16 was carried out by using low energy N+implantation. Double enzyme reaction system was used to detect the phospholipase A(PLA)activity ofPenicilliumexpansumunder the action of wild-type antagonistic strain and its mutant. The strain BA-16 was identified as Bacillus amyloliquefaciens. After the low energy N+implantation,the mutant BA-16-8,which showed the strongest antagonist capability and stable hereditary stability was selected out. In phospholipase activity detection,mutant showed stronger inhibition against PLA activity secreted byPenicilliumexpansumthan its wild-type strain. Moreover,as the concentration of fermentation broth increased,the inhibition effect enhanced,which indicated that it was possibly the inhibitory mechanism of antagonistic strain againstPenicilliumexpansum. The mutant BA-16-8 will be a potential biological control agent against apple blue mold decay.
Penicilliumexpansum;Bacillus amyloliquefaciens;low energy N+implantation;phospholipase A
2015-04-27
付瑞敏(1981-),女,在讀博士,講師,主要從事農業及食品微生物研究,E-mail:angelaminmin@163.com。
陳五嶺(1954-),男,碩士,教授,主要從事農業、環境及食品微生物研究,E-mail:wulingchen@yeah.net。
農業部科技成果與轉化項目(2012GB2G000451);河南省科技計劃項目(132102310253);河南省高等學校重點科研項目(15B180002);陜西省重大科技創新項目(2009ZKC04-16);河南教育學院青年科研課題項目(20100103);河南省基礎與前沿技術研究(152300410092)。
TS201.3
B
1002-0306(2016)05-0154-05
10.13386/j.issn1002-0306.2016.05.021
