微生物誘導(dǎo)下甲烷厭氧氧化及碳酸鹽礦物生成實(shí)驗(yàn)
2016-09-12 01:54:06劉肖許天福魏銘聰溫玉娟金光榮曹玉清
關(guān)鍵詞:實(shí)驗(yàn)質(zhì)量
劉肖,許天福,魏銘聰,溫玉娟,金光榮,曹玉清
(吉林大學(xué) 地下水資源與環(huán)境教育部重點(diǎn)實(shí)驗(yàn)室,吉林 長(zhǎng)春,130021)
微生物誘導(dǎo)下甲烷厭氧氧化及碳酸鹽礦物生成實(shí)驗(yàn)
劉肖,許天福,魏銘聰,溫玉娟,金光榮,曹玉清
(吉林大學(xué) 地下水資源與環(huán)境教育部重點(diǎn)實(shí)驗(yàn)室,吉林 長(zhǎng)春,130021)
從淤泥樣品中,培育甲烷氧化菌并對(duì)其進(jìn)行 DNA測(cè)序;進(jìn)而在自行研制的反應(yīng)系統(tǒng)中開展甲烷氧化菌-水-巖石相互作用實(shí)驗(yàn),檢測(cè)微生物的數(shù)量和 HCO3-質(zhì)量濃度的變化,最后對(duì)巖石樣品進(jìn)行掃描電鏡分析。研究結(jié)果表明:所培養(yǎng)的淡水甲烷氧化菌為假單胞菌屬;微生物注入反應(yīng)器后有HCO3-的生成,HCO3-質(zhì)量濃度變化與微生物數(shù)量和活性有關(guān);石英砂表面生成無定形鐵白云石、亮白色菱柱狀碳酸鹽和長(zhǎng)石類礦物的中間產(chǎn)物。
甲烷氧化菌;甲烷厭氧氧化;碳酸鹽礦物
甲烷是一種重要的溫室氣體,從生態(tài)學(xué)的角度分析,自然界中廣泛存在產(chǎn)甲烷菌,有機(jī)物被不斷地降解釋放甲烷[1],甲烷在大氣中的停留時(shí)間大約為 7.9 a[2-3],對(duì)全球氣候變暖的影響是二氧化碳的25倍[1-3]。而厭氧的海洋沉積物作為地球上最大的甲烷儲(chǔ)層,其中的甲烷或溶解在沉積物孔隙水中并向海底滲漏,或在適合的溫度壓力和地質(zhì)條件下形成天然氣水合物[4],海洋沉積物的甲烷生成率為(85~300)×1012g/a[5-7],若甲烷釋放到大氣中,則將嚴(yán)重加劇溫室效應(yīng)。但實(shí)際監(jiān)測(cè)結(jié)果顯示,產(chǎn)于海洋沉積物的甲烷對(duì)總甲烷氣體釋放量的貢獻(xiàn)較小(~2%)[7-8];同時(shí)聯(lián)合國(guó)環(huán)境保護(hù)署2010年的研究報(bào)告也顯示,占全球總面積71%的海洋對(duì)大氣圈自然排放甲烷總量的貢獻(xiàn)僅為3.5%[9],該控制甲烷氣體釋放的現(xiàn)象是由于海洋沉積物產(chǎn)生的大量甲烷向海底泄漏(或運(yùn)移)過程中被沉積物中厭氧生物所消耗,發(fā)生甲烷厭氧氧化反應(yīng)(anaerobic oxidation of methane, AOM)[5],并轉(zhuǎn)化為碳酸鹽[9]導(dǎo)致的。大量海洋地質(zhì)調(diào)查獲得的同位素分餾特征等地球化學(xué)數(shù)據(jù)表明[5,10]:甲烷厭氧氧化(AOM)反應(yīng)普遍存在,通常發(fā)生在冷泉區(qū)、泥火山和天然氣水合物儲(chǔ)層上方等富甲烷流體滲漏逃逸的地方。在AOM 區(qū)中,海底甲烷分別在甲烷氧化細(xì)菌和硫酸鹽還原細(xì)菌2種微生物的協(xié)同作用下[11-13],發(fā)生如下反應(yīng):CH4+SO42-→HCO3-+HS-+H2O,其中的甲烷作為電子供體,硫酸鹽作為電子受體,生成的重碳酸根可與地層中的陽(yáng)離子反應(yīng)生成碳酸鹽礦物[14-15]。甲烷厭氧氧化(AOM)消耗海洋沉積物產(chǎn)生的甲烷,并可將其轉(zhuǎn)化為碳酸鹽巖礦物保存下來,故研究甲烷厭氧氧化機(jī)理及其誘導(dǎo)下的礦物生成機(jī)制對(duì)了解其在全球碳循環(huán)和全球氣候變化的作用以及由甲烷氣泄漏引起的環(huán)境效應(yīng)的研究中均有著重要的意義。近年來,有關(guān)AOM的研究越來越多,但實(shí)驗(yàn)室內(nèi)關(guān)于AOM的研究多是用于確定其生物機(jī)理[16]、甲烷厭氧氧化速率[17-18]或者其同位素分餾特征[19]。如KNITTEL等[16]對(duì)甲烷厭氧氧化反應(yīng)的速率、環(huán)境影響因素以及ANME的作用機(jī)制進(jìn)行了介紹;NAUHAUS等[17-18]對(duì)甲烷厭氧氧化反應(yīng)的速率進(jìn)行了研究,結(jié)果顯示在不同的甲烷分壓情況下,AOM的反應(yīng)速率可差幾個(gè)數(shù)量級(jí)。梁洪野等[20-21]也進(jìn)行了甲烷氧化菌的分離鑒定相關(guān)工作,其初衷多是進(jìn)行淡水環(huán)境下甲烷氧化菌快速、高密度培養(yǎng)[20]以便應(yīng)用在工業(yè)生物催化中或者降解污水中的三氯甲烷、三氯乙烯等有毒鹵代烴和多環(huán)芳烴[21-22];近年來,國(guó)內(nèi)也開展了部分關(guān)于海洋環(huán)境中微生物方面的研究[23],但仍然處于起步階段。而在實(shí)驗(yàn)室條件下有關(guān)AOM的礦物生成機(jī)制以及其與生物活動(dòng)間關(guān)系的報(bào)道較少,關(guān)于生物活動(dòng)與礦物沉積間機(jī)理仍然不清楚。CHEN等[24]觀察到AOM過程中的細(xì)胞微生物團(tuán)被硅酸鹽包裹,并提出了AOM生物團(tuán)的礦物生成機(jī)制。因此,研究海底甲烷滲漏背景下的甲烷厭氧氧化誘導(dǎo)的礦物形成機(jī)制十分必要。本文作者采用實(shí)驗(yàn)室內(nèi)微生物培育技術(shù)、水化學(xué)分析以及礦物表面形態(tài)觀察和成分分析等方法,獲得甲烷-微生物實(shí)驗(yàn)的微生物、水化學(xué)、礦物組分結(jié)果,探討AOM 過程中微生物生長(zhǎng)以及水化學(xué)變化、礦物形成的機(jī)理。
1 實(shí)驗(yàn)
1.1 樣品采集
甲烷氧化菌廣泛分布于湖泊、沼澤、海底中,而海底底泥樣品不易取得,故為了便于實(shí)驗(yàn),本次采集長(zhǎng)春某湖底淤泥樣品培養(yǎng)甲烷氧化菌。
1.2 甲烷氧化菌培養(yǎng)
分別稱取淤泥樣品5 g,置于已滅菌并加入液體培養(yǎng)基(表 1)100 mL的錐形瓶中,用錫箔紙封口。用100 mL無菌注射器充入甲烷氣體(甲烷氣體購(gòu)于大連氣體廠,純度為99.99%),放置在30 ℃、轉(zhuǎn)速為110 r/min的震蕩培養(yǎng)箱中,培養(yǎng)6 d直到培養(yǎng)液混濁,得到微生物的母液,期間每隔1 d充1次甲烷氣體。為了使微生物更好地適應(yīng)富含甲烷的環(huán)境,逐漸增加充入的甲烷氣體量。作為空白對(duì)照的錐形瓶中,僅加入液體培養(yǎng)液和甲烷氣體進(jìn)行培養(yǎng),不添加淤泥樣品。

表1 培養(yǎng)基的成分(質(zhì)量濃度)Table 1 Composition of culture medium g/L
為了得到純度更高的微生物菌液,取20 mL已培養(yǎng)6 d的培養(yǎng)液上清液接種于新鮮培養(yǎng)基中,相同條件下培養(yǎng)6 d至培養(yǎng)液混濁,重復(fù)培養(yǎng)多次,培養(yǎng)期間逐漸加大充入的甲烷氣體量,獲得純度較高的微生物菌液。
通過母液的培養(yǎng)、傳代以及純化等步驟,得到了能夠利用甲烷進(jìn)行生長(zhǎng)的菌液。空白樣培養(yǎng)液清澈透明,說明培養(yǎng)出的菌量很少,可認(rèn)為幾乎沒有,其他的培養(yǎng)液混濁,說明培養(yǎng)的微生物為甲烷氧化菌。
1.3 菌種鑒定
為了進(jìn)一步確定所培養(yǎng)的微生物為甲烷氧化菌,本次實(shí)驗(yàn)進(jìn)行了菌種的分子生物學(xué)鑒定。采用平板涂布法(表 1)對(duì)甲烷氧化菌進(jìn)行純種分離,待菌落生長(zhǎng)后挑取單菌落采用劃線平板法分離純化,獲得優(yōu)勢(shì)菌株,并郵寄至上海生工進(jìn)行分子生物學(xué)鑒定。甲烷氧化菌的 DNA提取使用 UNIQ-10柱式細(xì)菌基因組DNA抽提試劑盒(上海生工),提取流程按照DNA提取盒的說明進(jìn)行。用細(xì)菌 16S rDNA通用引物27F(5’-AGTTTGATCMTGGCTCAG-3’)和 1492R(5’-GGTTACCTTGTTACGACTT-3’)擴(kuò)增16S rDNA片段。PCR體系選用25 μL反應(yīng)體系:基因組DNA(20~50 mg/L)0.5 μL;5×PCR 緩沖液 2.5 μL;dNTP(各
2.5 mmol/L) 1 μL;F(10 μmol/L) 0.5 μL;R(10 μmol/L)0.5 μL;ddH2O補(bǔ)足體積。在PCR儀上進(jìn)行擴(kuò)增,條件為:98 ℃預(yù)變性3 min;98 ℃時(shí)進(jìn)行25 s,55 ℃時(shí)進(jìn)行25 s,72 ℃時(shí)進(jìn)行1 min,30個(gè)循環(huán);72 ℃修復(fù)延伸10 min;4 ℃保存。
根據(jù)獲得的16S rDNA序列,采用BLAST對(duì)菌種的DNA序列進(jìn)行同源性對(duì)比,獲知其菌種屬性。
1.4 微生物-水-巖石-甲烷相互作用實(shí)驗(yàn)
1.4.1 實(shí)驗(yàn)裝置
實(shí)驗(yàn)在自行研制的多功能水合物模擬實(shí)驗(yàn)系統(tǒng)(圖1)中進(jìn)行,該系統(tǒng)專為微生物-水-巖石-甲烷相互作用及其次生礦物生成實(shí)驗(yàn)設(shè)計(jì)。系統(tǒng)由供氣、微生物注入、反應(yīng)釜、控溫、數(shù)據(jù)采集等子系統(tǒng)構(gòu)成,反應(yīng)釜的結(jié)構(gòu)采用快開式以方便拆卸,反應(yīng)釜內(nèi)徑為80 mm,高為 200 mm,有效體積為 1 000 mL,耐壓值為25 MPa。釜體安裝熱電阻溫度傳感器和壓力傳感器。溫度傳感器(Pt100)探針伸入到釜體液面(溶液)中20 mm,精度為±0.1 ℃。壓力傳感器最大量程為 40 MPa,精度為±0.25%,并配有壓力表以方便觀察,實(shí)驗(yàn)過程中的溫度和壓力變化由數(shù)據(jù)采集系統(tǒng)自動(dòng)記錄。實(shí)驗(yàn)時(shí)反應(yīng)釜內(nèi)分為飽水的砂、水、氣三層,由微生物注入系統(tǒng)向低溫高壓反應(yīng)釜中注入甲烷氧化菌,實(shí)驗(yàn)過程中取水樣測(cè)試HCO3-等的變化以探究甲烷氧化機(jī)理。

圖1 高壓反應(yīng)釜實(shí)驗(yàn)裝置示意圖Fig.1 Schematic diagram of experimental device of high pressure reactor
1.4.2 實(shí)驗(yàn)條件
根據(jù)GUAN等[25]的研究,甲烷厭氧氧化可發(fā)生在水合物儲(chǔ)層上方、泥火山等甲烷滲漏區(qū),其在水深約470 m到大于3 000 m的海底均有發(fā)現(xiàn),其相應(yīng)的溫度在 0~8 ℃之間,溫度相對(duì)較低;相應(yīng)的壓力在4.0~30.0 MPa間變化。因此,在現(xiàn)有的條件下,本文開展了 2組微生物-水-巖石-甲烷相互作用實(shí)驗(yàn),在固定實(shí)驗(yàn)溫度為1.8 ℃條件下,分別進(jìn)行了壓力為4.5和7.0 MPa的2組實(shí)驗(yàn)。
1.4.3 實(shí)驗(yàn)步驟及測(cè)試分析
在將培養(yǎng)好的菌液注入反應(yīng)釜前,選擇生長(zhǎng)好的甲烷氧化菌進(jìn)行擴(kuò)增、濃縮后用紫外分光光度計(jì)(UV2100,尤尼科(上海)儀器有限公司)調(diào)節(jié)甲烷氧化菌菌液的光密度(OD)為1(OD為被檢測(cè)物吸收掉的光密度,能夠定性表征甲烷氧化菌菌液的濃度,本次實(shí)驗(yàn)在600 nm下測(cè)定)。將OD為1的甲烷氧化菌菌液50 mL注入到微生物容器(圖1)中,通過平流泵將微生物容器中的微生物菌液注入到反應(yīng)釜中。注入后,取液體樣,分別進(jìn)行HCO3-質(zhì)量濃度的測(cè)定和甲烷氧化菌的平板計(jì)數(shù)。HCO3-質(zhì)量濃度的測(cè)定采用酸堿中和滴定法測(cè)試,單位為mg/L;而甲烷氧化菌的平板計(jì)數(shù)采用涂布平板法,單位為個(gè)。在實(shí)驗(yàn)結(jié)束后,將巖石樣品烘干并進(jìn)行掃描電鏡分析,巖石片的表面形態(tài)觀察在吉林大學(xué)古生物中心實(shí)驗(yàn)室中進(jìn)行,利用配置的JSM-6700F型掃描電鏡(scanning electron microscope,SEM, JEOL Corporation, Tokyo, Japan)和INCAX-SIGHT型能譜儀(energy-dispersive X-ray spectrometer, EDS,Oxford, UK)進(jìn)行表面形態(tài)觀察和礦物成分分析。
2 實(shí)驗(yàn)結(jié)果與分析
2.1 微生物鑒定結(jié)果
由菌落微生物提取的總DNA在凝膠成像系統(tǒng)下拍照結(jié)果可知:各菌落均得到1條較為清晰的電泳條帶,說明其基因組DNA提取有效。對(duì)以上DNA進(jìn)行PCR擴(kuò)增,結(jié)果如圖2所示。與Marker比較可知,擴(kuò)增后各菌落微生物的基因片段約為1 400 bp。

圖2 各菌落16S rDNA PCR 擴(kuò)增產(chǎn)物瓊脂糖電泳Fig.2 Agarose gel electrophoresis of PCR amplified 16S rDNA for different samples
利用菌落的DNA序列采用BLAST進(jìn)行同源性序列對(duì)比:圖中S1408菌屬于假單胞菌屬,與假單胞菌fluorescens strain LMG 14576 16S同源性為100%; S1409菌屬于假單胞菌屬,與假單胞菌putida H8234菌株序列同源性為99%;S1410菌屬于不動(dòng)桿菌屬,與不動(dòng)桿菌屬calcoaceticus strain YLZZ-1 16S同源性為 99%;S1411菌屬假單胞菌屬,與假單胞菌fluorescens strain LMG 14576 16S同源性為99%;由此判定所培養(yǎng)的微生物為假單胞菌屬的甲烷氧化菌。
2.2 微生物-水-巖石-甲烷相互作用結(jié)果
微生物注入后實(shí)驗(yàn)過程中HCO3-質(zhì)量濃度隨時(shí)間的變化如圖3(a)和4(a)所示。由圖3(a)和4(a)可以看出:甲烷氧化菌菌液注入后,其t=0時(shí)刻的HCO3-質(zhì)量濃度為注入菌液中含有的 HCO3-被稀釋的結(jié)果,HCO3-質(zhì)量濃度初始時(shí)刻背景值分別為23 mg/L和10 mg/L,而微生物注入后的微生物數(shù)量背景值分別為1.45×106和1.70×106個(gè);甲烷氧化菌在飽和甲烷的反應(yīng)釜中生長(zhǎng),代謝產(chǎn)生HCO3-,HCO3-質(zhì)量濃度增加,4.5 MPa(實(shí)驗(yàn) 1)時(shí),HCO3-質(zhì)量濃度最高約 47 mg/L,7.0 MPa(實(shí)驗(yàn)2)時(shí)最高能達(dá)到49 mg/L;隨后HCO3-質(zhì)量濃度降低,接近但高于其初始質(zhì)量濃度。
從圖3(a)可以看出:HCO3-質(zhì)量濃度隨時(shí)間呈現(xiàn)先增加(0~90 min)后減小(90~150 min)的趨勢(shì)。反應(yīng)液中 HCO3-質(zhì)量濃度的變化是微生物-甲烷相互作用的結(jié)果,注入的甲烷氧化菌利用甲烷單加氧酶氧化反應(yīng)釜內(nèi)的甲烷氣,發(fā)生的反應(yīng)式為:CH4+3H2O→HCO3-+4H2+H+,產(chǎn)生 HCO3-,這個(gè)反應(yīng)是由微生物引起的反向甲烷生成反應(yīng)[26-28],從熱力學(xué)角度來說是可以實(shí)現(xiàn)的[27]。微生物注入后,微生物需適應(yīng)反應(yīng)釜中的新環(huán)境,原菌液中營(yíng)養(yǎng)物質(zhì)充分,部分微生物活性較高;且微生物在適應(yīng)新環(huán)境過程中不斷地代謝產(chǎn)生HCO3-,HCO3-質(zhì)量濃度增加,在90 min時(shí)HCO3-質(zhì)量濃度達(dá)到最大值47.65 mg/L。而在適應(yīng)新環(huán)境的過程中,部分微生物不能適應(yīng)新環(huán)境而導(dǎo)致生物活性受到抑制或死亡,微生物數(shù)量降低,90 min時(shí)微生物的數(shù)量最小達(dá)到9.30×105個(gè)(圖3(b))。微生物在適應(yīng)了新環(huán)境后,利用原有菌液中的營(yíng)養(yǎng)物質(zhì)生長(zhǎng),微生物數(shù)量增加到 2.75×106個(gè)(90~150 min)。隨后由于HCO3-與溶液中的某些陽(yáng)離子結(jié)合生成沉淀消耗HCO3-,消耗量大于微生物作用下甲烷厭氧氧化反應(yīng)生成的量,因此,HCO3-質(zhì)量濃度在逐漸減小。由于實(shí)驗(yàn) 1時(shí)間較短,150 min時(shí)的 HCO3-質(zhì)量濃度為29.32 mg/L,比初始時(shí)刻的23.46 mg/L高出5.86 mg/L。因此,在實(shí)驗(yàn)2時(shí)加長(zhǎng)了微生物與甲烷相互作用的時(shí)間,以期獲得效果更好的HCO3-質(zhì)量濃度變化數(shù)據(jù)。

圖3 4.5 MPa時(shí)HCO3-質(zhì)量濃度和微生物數(shù)量隨時(shí)間的變化Fig.3 Changes of mass concentration of HCO3-and microbial quantity over time at 4.5 MPa
從圖4(a)可以看出:HCO3-質(zhì)量濃度隨時(shí)間也是先增加(0~90 min)后減小(90~1 170 min)的趨勢(shì),在90 min時(shí)HCO3-質(zhì)量濃度達(dá)到最大值48.45 mg/L。同實(shí)驗(yàn) 1,此時(shí)的微生物最活躍,對(duì)應(yīng)圖4(b)中微生物數(shù)量最少。微生物在適應(yīng)了環(huán)境的過程中,利用原有菌液中的營(yíng)養(yǎng)物質(zhì)生長(zhǎng),微生物數(shù)量增加到2.85×106個(gè)(150~210 min間)。210 min后,微生物繼續(xù)利用反應(yīng)釜內(nèi)的營(yíng)養(yǎng)物質(zhì)生長(zhǎng),微生物的數(shù)量超過3.00×106個(gè),無法計(jì)數(shù),但是能看出微生物數(shù)量增加。在1 170 min時(shí),檢測(cè)溶液中HCO3-質(zhì)量濃度降為4.39 mg/L,這是HCO3-與溶液中的某些陽(yáng)離子結(jié)合生成沉淀所造成的。

圖4 7.0 MPa時(shí)HCO3-質(zhì)量濃度和微生物數(shù)量隨時(shí)間的變化Fig.4 Changes of mass concentration of HCO3-and microbial quantity over time at 7.0 MPa
2次不同壓力下的實(shí)驗(yàn)顯示:微生物在不同壓力下的HCO3-質(zhì)量濃度均呈先增加后減小的趨勢(shì),達(dá)到HCO3-質(zhì)量濃度峰值點(diǎn)所需的時(shí)間基本一致。HCO3-質(zhì)量濃度最高的時(shí)候是微生物個(gè)數(shù)最低的時(shí)候,HCO3-質(zhì)量濃度的增大和減小能夠表征微生物的生長(zhǎng)及其活性衰減情況。在整個(gè)甲烷厭氧氧化反應(yīng)發(fā)生的過程中,微生物不斷的適應(yīng)新環(huán)境,從初始時(shí)刻到實(shí)驗(yàn)結(jié)束時(shí),微生物的形態(tài)發(fā)生了改變(圖5),微生物菌落的形態(tài)由開始的直徑3 mm左右的圓形菌轉(zhuǎn)變?yōu)楹笃诘? mm左右的圓形菌居多。其中的甲烷氧化菌為甲烷的氧化提供動(dòng)力,促使甲烷厭氧氧化反應(yīng)的發(fā)生。
實(shí)驗(yàn)結(jié)束后,在石英砂表面發(fā)現(xiàn)了含 Ca,F(xiàn)e和Mg的碳酸鹽礦物。典型的碳酸礦物照片如圖6(a)和6(b)所示,其中圖6(a)所示為團(tuán)狀的無定形礦物,含Mg,Ca,F(xiàn)e,Na,Si,C和O等組分(圖6(c)),推斷為鐵白云石。而實(shí)驗(yàn)所用石英砂主要含石英和長(zhǎng)石類礦物,因此掃描電鏡結(jié)果所含的Al和Si為背景礦物所含組分。而圖6(b)所示為亮白色菱柱狀礦物,含Mg,Ca,F(xiàn)e,Na,Si,C和O等組分(圖6(d)),推斷為碳酸鹽礦物和長(zhǎng)石類礦物的中間產(chǎn)物。
基于現(xiàn)有的實(shí)驗(yàn)結(jié)果,結(jié)合CHEN等[24]關(guān)于甲烷厭氧氧化的研究成果,將甲烷厭氧氧化中微生物的活動(dòng)以及礦物沉積間的關(guān)系表示如圖7所示。由于甲烷氧化菌的代謝作用,反應(yīng)釜中的甲烷被微生物利用,發(fā)生甲烷厭氧氧化反應(yīng),生成了HCO3-,導(dǎo)致溶液堿度增加,HCO3-與溶液中的陽(yáng)離子結(jié)合,發(fā)生反應(yīng):HCO3-+Ca2+(Mg2+, Fe2+)→CaCO3(MgCO3, FeCO3)+H+,生成碳酸鹽礦物。
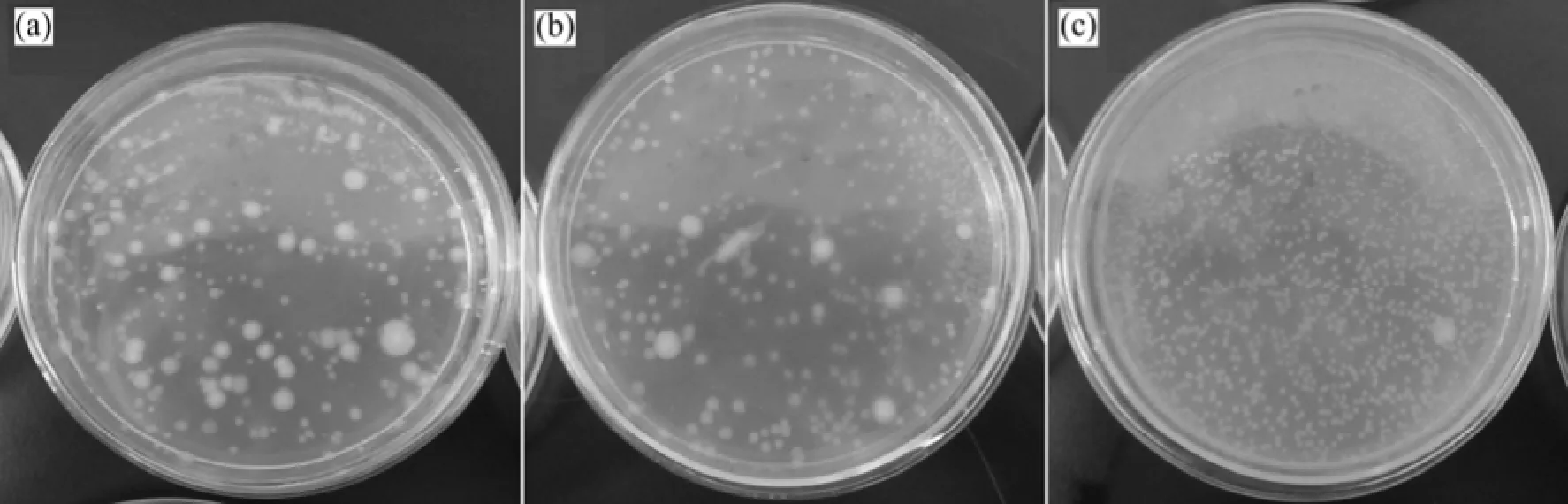
圖5 平板計(jì)數(shù)照片F(xiàn)ig.5 Photos of plate count

圖6 掃描電鏡觀察結(jié)果Fig.6 Results from SEM observation

圖7 甲烷氧化菌與碳酸鹽礦物沉淀的關(guān)系[24]Fig.7 Relationship between microbial metabolism and precipitation of carbonate[24]
3 結(jié)論
1) 本室內(nèi)實(shí)驗(yàn)培養(yǎng)的甲烷氧化菌為假單胞菌屬,可利用甲烷作為碳源生長(zhǎng)。
2) 微生物注入反應(yīng)釜后,甲烷被消耗,發(fā)生了甲烷厭氧氧化反應(yīng)CH4+3H2O→HCO3-+4H2+H+,實(shí)驗(yàn)獲得的HCO3-質(zhì)量濃度變化與微生物數(shù)量和活性有關(guān)。HCO3-的這種變化趨勢(shì)間接地表征微生物的生長(zhǎng)和活性衰減情況。
3) 甲烷氧化菌注入低溫、高壓條件的反應(yīng)器后,需要一定時(shí)間適應(yīng)新的環(huán)境,適應(yīng)環(huán)境后,微生物數(shù)量增加;但后期因營(yíng)養(yǎng)物質(zhì)得不到補(bǔ)充,甲烷氧化菌生物活性衰減。
4) 在甲烷氧化細(xì)菌的作用下,石英砂表面生成了無定形鐵白云石以及亮白色菱柱狀碳酸鹽和長(zhǎng)石類礦物的中間產(chǎn)物,印證了甲烷厭氧氧化反應(yīng)的發(fā)生。
[1] 韓冰. 甲烷氧化菌的微生態(tài)解析及其應(yīng)用基礎(chǔ)研究[D]. 北京:清華大學(xué)化學(xué)工程系, 2008: 3-23. HAN Bing. Molecular ecology study on methanotrophs and fundermantal research on the applications of methanotrophs[D]. Beijing: Tsinghua University. Department of Chemical Engineering, 2008: 3-23.
[2] 陳忠, 顏文, 陳木宏, 等. 海底天然氣水合物分解與甲烷歸宿研究進(jìn)展[J]. 地球科學(xué)進(jìn)展, 2006, 21(4): 394-400. CHEN Zhong, YAN Wen, CHEN Muhong, et al. Advances in gas hydrate dissociation and fate of methane in marine sediment[J]. Advances in Earth Science, 2006, 21(4): 394-400.
[3] LELIEVELD J O S, CRUTZEN P J, DENTENER F J. Changing concentration, lifetime and climate forcing of atmospheric methane[J]. Tellus B, 1998, 50(2): 128-150.
[4] DALE A W, VAN CAPPELLEN P, AGUILERA D R, et al. Methane efflux from marine sediments in passive and active margins: estimations from bioenergetic reaction-transport simulations[J]. Earth and Planetary Science Letters, 2008, 265(3):329-344.
[5] LIU L, WU N. Simulation of advective methane flux and AOM in Shenhu area, the northern South China Sea[J]. Environmental Earth Sciences, 2014, 71(2): 697-707.
[6] KNITTEL K, BOETIUS A. Anaerobic oxidation of methane:progress with an unknown process[J]. Annual Review of Microbiology, 2009, 63: 311-334.
[7] REEBURGH W S. Oceanic methane biogeochemistry[J]. Chemical Reviews, 2007, 107(2): 486-513.
[8] REEBURGH W S. Global methane biogeochemistry[J]. Treatise on Geochemistry, 2003, 4: 65-89.
[9] ANDERSON B, BARTLETT K, FROLKING S, et al. Methane and nitrous oxide emissions from natural sources[R]. Washington: United States Environmental Protection Agency,Office of Atmospheric Programs, 2010: 1-194.
[10] REEBURGH W S. Methane consumption in Cariaco Trench waters and sediments[J]. Earth and Planetary Science Letters,1976, 28(3): 337-344.
[11] BOETIUS A, RAVENSCHLAG K, SCHUBERT C J, et al. A marine microbial consortium apparently mediating anaerobic oxidation of methane[J]. Nature, 2000, 407(6804): 623-626.
[12] HINRICHS K U, HAYES J M, SYLVA S P, et al. Methaneconsuming archaebacteria in marine sediments[J]. Nature, 1999,398(6730): 802-805.
[13] ORPHAN V J, HOUSE C H, HINRICHS K U, et al. Methane-consuming archaea revealed by directly coupled isotopic and phylogenetic analysis[J]. Science, 2001, 293(5529):484-487.
[14] KOCHERLA M. Authigenic gypsum in gas-hydrate associated sediments from the east coast of India (bay of bengal)[J]. Acta Geologica Sinica, 2013, 87(3): 749-760.
[15] SUN Z L, WEI H L, ZHANG X H, et al. A unique Fe-rich carbonate chimney associated with cold seeps in the Northern Okinawa Trough, East China Sea[J]. Deep Sea Research Part Ⅰ:Oceanographic Research Papers, 2015, 95: 37-53.
[16] KNITTEL K, BOETIUS A. Anaerobic oxidation of methane:progress with an unknown process[J]. Annual review of microbiology, 2009, 63: 311-334.
[17] NAUHAUS K, BOETIUS A, KRüGER M, et al. In vitro demonstration of anaerobic oxidation of methane coupled to sulphate reduction in sediment from a marine gas hydrate area[J]. Environmental Microbiology, 2002, 4(5): 296-305.
[18] ZHANG Y, HENRIET J P, BURSENS J, et al. Stimulation of in vitro anaerobic oxidation of methane rate in a continuous high-pressure bioreactor[J]. Bioresource Technology, 2010, 101(9): 3132-3138.
[19] DEUSNER C, HOLLER T, ARNOLD G L, et al. Sulfur and oxygen isotope fractionation during sulfate reduction coupled to anaerobic oxidation of methane is dependent on methane concentration[J]. Earth and Planetary Science Letters, 2014, 399:61-73.
[20] 梁洪野. 甲烷氧化菌的高密度培養(yǎng)[D]. 哈爾濱: 哈爾濱商業(yè)大學(xué)食品工程學(xué)院, 2011: 1-6. LIANG Hongye. High cell density cultivation of methanotrophic bacteria[D]. Harbin: Harbin University of Commerce. College of Food Engineering, 2011: 1-6.
[21] 王曉麗, 于建國(guó). 一個(gè)甲烷氧化菌株的分離, 鑒定及其特性研究[J]. 微生物學(xué)通報(bào), 2008, 35(6): 934-938. WANG Xiaoli, YU Jianguo. Isolation, identification and characterization of a methanotrophic strain[J]. Microbiology,2008, 35(6): 934-938.
[22] 梁戰(zhàn)備, 史奕, 岳進(jìn). 甲烷氧化菌研究進(jìn)展[J]. 生態(tài)學(xué)雜志,2004, 23(5): 198-205. LIANG Zhanbei, SHI Yi, YUE Jin. The research progress of Methane oxidizing bacteria[J]. Chinese Journal of Ecolog, 2004,23(5): 198-205.
[23] 史春瀟, 雷懷彥, 趙晶, 等. 南海北部九龍甲烷礁鄰區(qū)沉積物層中垂向細(xì)菌群落結(jié)構(gòu)特征研究[J]. 沉積學(xué)報(bào), 2014, 32(6):1072-1082. SHI Chunxiao, LEI Huaiyan, ZHAO Jing, et al. Vertical microbial community structure characteristics of sediment in gas hydrate potential area of Northern South China Sea Jiulong Methane Reef[J]. Acta Sedimentologica Sinica, 2014, 32(6):1072-1082.
[24] CHEN Y, LI Y L, ZHOU G T, et al. Biomineralization mediated by anaerobic methane-consuming cell consortia[J]. Scientific Reports, 2014, 4(4): 5696.
[25] GUAN H, SUN Y, ZHU X, et al. Factors controlling the types of microbial consortia in cold-seep environments: a molecular and isotopic investigation of authigenic carbonates from the South China Sea[J]. Chemical Geology, 2013, 354: 55-64.
[26] HANSEN L B, FINSTER K, FOSSING H, et al. Anaerobic methane oxidation in sulfate depleted sediments: effects of sulfate and molybdate additions[J]. Aquatic Microbial Ecology,1998, 14(2): 195-204.
[27] HEIJS S K, ALOISI G, BOULOUBASSI I, et al. Microbial community structure in three deep-sea carbonate crusts[J]. Microbial Ecology, 2006, 52(3): 451-462.
[28] PAPE T, BLUMENBERG M, SEIFERT R, et al. Marine methane biogeochemistry of the Black Sea: a review[C]// Links between Geological Processes, Microbial Activities & Evolution of Life. Netherlands: Springer, 2008: 281-311.
(編輯 楊幼平)
Experiment on anaerobic oxidation of methane and precipitation of carbonate mediated by microbes
LIU Xiao, XU Tianfu, WEI Mingcong, WEN Yujuan, JIN Guangrong, CAO yuqing
(Key Laboratory of Groundwater Resources and Environment, Ministry of Education, Jilin University,Changchun 130021, China)
The methane oxidizing bacteria was isolated from lake bottom sediment and the corresponding DNA sequencing was carried out. Experiment of interaction between methane oxidizing bacteria-water-rock was conducted in a self-developed reactor system, and variations of the number of microorganisms and the mass concentration of HCO3-were analyzed, respectively. Finally, rock slices were viewed with scanning electron microscope (SEM). The results show that the cultured methane oxidizing bacteria belongs to pseudomonas genus. HCO3-produced after bacteria injected into the reactor. Changes of mass concentration of HCO3-have certain correlation relationship with the number and microbial activity of microorganism. Intermediate minerals, including amorphous ankerite, bright columnar of carbonate and feldspars, are precipitated on the surface of quartz sandstone. Key words: methane oxidizing bacteria; anaerobic oxidation of methane; carbonates
P593
A
1672-7207(2016)05-1473-07
10.11817/j.issn.1672-7207.2016.05.003
2015-05-21;
2015-07-16
中國(guó)博士后面上基金資助項(xiàng)目(2015M571369);吉林大學(xué)研究生創(chuàng)新基金資助項(xiàng)目(2014104) (Project(2015M571369)supported by the Postdoctoral Science Foundation of China; Project(2014104) supported by the Graduate Innovation Fund of Jilin University)
許天福,博士,教授,從事海洋天然氣水合物試開采優(yōu)化和相關(guān)自生礦物形成、地?zé)崮荛_發(fā)與利用等研究;E-mail: Tianfu_Xu@jlu.edu.cn
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中學(xué)生數(shù)理化·中考版(2022年11期)2022-02-16 07:01:20
中學(xué)生數(shù)理化·八年級(jí)物理人教版(2021年12期)2021-12-31 03:23:08
中學(xué)生數(shù)理化·中考版(2020年10期)2020-11-27 01:59:48
小哥白尼(趣味科學(xué))(2019年6期)2019-10-10 01:01:50
中國(guó)生殖健康(2019年2期)2019-08-23 08:12:08
產(chǎn)品可靠性報(bào)告(2017年7期)2017-09-05 09:49:12
發(fā)明與創(chuàng)新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
汽車觀察(2016年3期)2016-02-28 13:16:26
