谷類食品中克羅諾桿菌的分離與鑒定
2016-09-12 00:47:56李遠宏張逸飛張慶成鄒小倩裴尚飛趙珉生
食品工業科技 2016年15期
李遠宏,張逸飛,張慶成,徐 灣,鄒小倩,裴尚飛,趙珉生,姜 華
(1.徐州醫科大學公共衛生學院,江蘇徐州 221004;2.空軍勤務學院航空軍需系,江蘇徐州 221000)
?
谷類食品中克羅諾桿菌的分離與鑒定
李遠宏1,張逸飛1,張慶成2,徐灣1,鄒小倩1,裴尚飛1,趙珉生1,姜華1
(1.徐州醫科大學公共衛生學院,江蘇徐州 221004;2.空軍勤務學院航空軍需系,江蘇徐州 221000)
為了解谷類食品中克羅諾桿菌(Cronobacterspp.)的污染情況。本研究采集了100份谷類食品樣品,采用國家標準GB 4789.40-2010分離克羅諾桿菌,并利用基于ITS和OmpA特異基因片段的PCR鑒定方法、16S rDNA基因序列分析等方法對可疑克羅諾桿菌進行了初步的鑒定。結果發現谷類食品中克羅諾桿菌的污染率為21.0%,并分離出21株可疑克羅諾桿菌,其中17株(占總數的81.0%)被初步鑒定為阪崎克羅諾桿菌(C.sakazakii),3株(占總數的14.2%)為都柏林克羅諾桿菌(C.dublinensis),1株(占總數的4.8%)可能為尤尼沃斯克羅諾桿菌(C.universalis)或蘇黎世克羅諾桿菌(C.turicensis)。研究結果表明市售谷類食品中存在食源性克羅諾桿菌的污染,應該加強谷類食品中克羅諾桿菌的流行病學監測。
克羅諾桿菌,谷類食品,分離,鑒定
克羅諾桿菌(Cronobacterspp.),原阪崎腸桿菌(Enterobactersakazakii),是近年來嬰幼兒配方奶粉中引起廣泛關注的一種食源性條件致病菌。它是一種周生鞭毛、能運動、無芽孢的革蘭陰性細菌,在一定條件下可引起人和動物致病[1]。近期的分類學研究結果表明該菌包括7個種:阪崎克羅諾桿菌(C.sakazakii),丙種二酸鹽陽性克羅諾桿菌(C.malonaticus),蘇黎世克羅諾桿菌(C.turicensis),都柏林克羅諾桿菌(C.dublinensis),穆汀斯克羅諾桿菌(C.muytjensis),康迪蒙提克羅諾桿菌(C.condimenti)和尤尼沃斯克羅諾桿菌(C.universalis)[2]。克羅諾桿菌主要通過嬰幼兒配方奶粉感染新生兒及嬰幼兒,特別是早產兒、低出生體重的嬰幼兒,引起腦膜炎、菌血癥和壞死性小腸結腸炎等疾病,死亡率高達20%~50%[3-4]。此外,研究表明該菌也能導致老年人和免疫力低下的人群致病[5]。
很多報道證實克羅諾桿菌主要的傳染源和傳播媒介是嬰兒配方粉,然而在奶酪、肉制品、谷物制品、香腸、香辛料、蔬菜等各種食品中也分離出了克羅諾桿菌[6-9]。此外,克羅諾桿菌還廣泛存在于食品生產企業的各種加工設備及生產環境中[10]。目前,不同學者對克羅諾桿菌的真正宿主、傳播途徑和致病機制還沒達成統一的認識。一些學者認為嬰幼兒配方奶粉及其加工環境、面粉及其生產環境可能是克羅諾桿菌的天然儲藏場所,在克羅諾桿菌的傳播過程中起重要作用[11-12]。
董曉暉等[2]對非奶粉類食品中克羅諾桿菌的污染情況做定量檢測,并從冷凍面條中分離出了克羅諾桿菌,檢出的克羅諾桿菌數量為0.62 MPN/g。張翼等[13]對嬰幼兒食品中克羅諾桿菌分離株的生物被膜形成能力進行了研究,結果表明從嬰兒米粉和嬰兒面條等樣品中分離的克羅諾桿菌均具有一定生物膜成膜能力。污染了克羅諾桿菌的谷類食品將可能導致新生兒及嬰幼兒致病,但目前國內外尚未有較為全面的關于谷類食品中克羅諾桿菌污染情況的研究資料。進一步了解各類食品中克羅諾桿菌的分布特征和污染狀況可為預防該菌引起的疾病的爆發、流行及追蹤污染源提供理論依據,具有重要的公共衛生學意義。本文主要對常見谷類食品中克羅諾桿菌的污染狀況進行了檢測,并利用基于ITS(Internal Transcribed Spacer,內轉錄間隔區)和OmpA(Outer membrane protein A,外膜蛋白A)基因片段的特異性PCR鑒定和16S rDNA基因序列分析等方法對可疑克羅諾桿菌進行了初步的鑒定。
1 材料與方法
1.1材料與儀器
谷類食品2014年9月至2015年6月,從湖南、湖北、江蘇、上海等地共采集谷類食品樣品100份,包括玉米碴13份,大米9份,紅豆8份,燕麥片和綠豆7份,黃豆、薏米、小黃米、蕎麥粉、小麥粉、糯米和黑米各6份;高粱米和黑豆各5份,西米4份(表1)。樣品采集完畢后立即送回實驗室檢測。
克羅諾桿菌標準菌株阪崎克羅諾桿菌C.sakazakiiCICC 21560和穆汀斯克羅諾桿菌C.muytjensiiCICC 21563由南京農業大學食品科技學院酶工程實驗室饋贈;緩沖蛋白胨水(BPW)、改良月桂基硫酸鹽胰蛋白胨肉湯(mLST)、萬古霉素、胰蛋白胨大豆瓊脂(TSA)北京陸橋技術有限責任公司;克羅諾桿菌顯色培養基HiCromeTMCronobacterspp. agar(配方:酪蛋白15.0 g/L,大豆蛋白胨5.0 g/L,顯色混合物10.17 g/L,氯化鈉5.0 g/L,瓊脂15.0 g/L)美國sigma公司;細菌基因組DNA提取試劑盒美國Omega公司;2× Taq Master Mix南京諾唯贊生物科技有限公司;引物由上海生工生物工程有限公司合成。
DNP9162電熱恒溫培養箱上海精宏;SW-CJ-2FD超凈工作臺蘇州安泰;Veriti 96 PCR儀美國ABI公司;5418型小型高速離心機德國Eppendorf公司;Gel Doc XR凝膠成像儀美國BIO-RAD公司。
1.2實驗方法
1.2.1分離方法按照國家標準GB 4789.40-2010《食品衛生微生物學檢驗阪崎腸桿菌檢驗》中的方法對100份谷類食品中的克羅諾桿菌進行分離鑒定[14]。
1.2.2基因組DNA提取將純化后的可疑分離株接種于LB培養基中,置于搖床中于37 ℃,180 r/min過夜培養,取過夜培養液1~2 mL,參照美國Omega公司的細菌基因組DNA提取試劑盒的操作說明書提取細菌總DNA。
1.2.3PCR擴增ITS基因特異性片段按照參考文獻[15]的方法采用PCR方法擴增克羅諾桿菌ITS基因特異性片段。正向引物SG-F:5′-GGGTTGTCTG CGAAAGCGAA-3′;反向引物SG-R:5′-GTCTTC GTGCTGCGAGTTTG-3′。PCR反應體系:2×Taq Master Mix 25 μL,引物SG-F和SG-R各2 μL,DNA模板1 μL,加水補足至50 μL。PCR反應條件:95 ℃預變性3 min;95 ℃變性30 s,57 ℃退火1 min,72 ℃延伸1 min,35個循環;72 ℃延伸7 min。反應結束后,取PCR產物經2%瓊脂糖凝膠電泳,在凝膠成像系統下觀察并記錄實驗結果。
1.2.4PCR擴增OmpA基因特異性片段按照參考文獻[16]的方法采用PCR方法擴增克羅諾桿菌OmpA基因特異性片段。正向引物ESSF:5′-GGATTTAACCGTGAACTTTTCC-3′;反向引物ESSR:5′-CGCCAGCGATGTTAGAAGA-3′。PCR反應體系:2×Taq Master Mix 25 μL,引物ESSF和ESSR各2 μL,DNA模板1 μL,加水補足至50 μL。PCR反應條件:95 ℃預變性5 min;95 ℃變性30 s,55 ℃退火45 s,72 ℃延伸50 s,35個循環;72 ℃延伸7 min。反應結束后,取PCR產物經1%瓊脂糖凝膠電泳,在凝膠成像系統下觀察并記錄實驗結果。
1.2.516S rDNA基因擴增及測序以細菌總DNA為模板,用細菌16S rDNA通用引物進行PCR擴增[17]。正向引物fD1:5′-AGAGTTTGATCCTGG CTCAG-3′和反向引物rP1:5′-ACGGTTACCTTGTT ACGACTT-3′。PCR反應體系:2×Taq Master Mix 25 μL,引物(10 μmol/L)各2 μL,DNA模板1 μL,加無菌雙蒸水至50 μL。PCR反應條件:95 ℃預變性2 min;95 ℃變性45 s,55 ℃退火55 s,72 ℃延伸2 min,35個循環;72 ℃延伸7 min。PCR產物經1%瓊脂糖凝膠電泳檢測后送南京金思瑞生物科技有限公司測序。將測序獲得的16S rDNA基因序列提交至GenBank數據庫(http://www.ncbi.nlm.nih.gov/),用Blastn進行序列同源性比對。采用Clustalx 2.0軟件將16S rDNA基因序列進行多重比較后,利用MEGA 4.0軟件采用鄰近法構建系統發育樹。
2 結果與分析
2.1克羅諾桿菌污染情況
將在克羅諾桿菌顯色培養基上生長呈藍綠色,且在TSA平板上于25 ℃培養48 h呈黃色的菌落初步確定為可疑克羅諾桿菌分離株。通過顯色平板及TSA平板共檢出24份谷類食品樣本污染了克羅諾桿菌,檢出率為24.0%(表1)。對可疑分離株進行了16S rDNA測序鑒定,結果最終證實21份谷類食品樣品污染了克羅諾桿菌,檢出率為21.0%(表1)。本研究從綠豆、紅豆、玉米、黃豆、蕎麥粉、小黃米、黑豆、小麥粉、糯米、燕麥、黑米和薏米等樣品中分離出了克羅諾桿菌,其中污染率較高的樣品有蕎麥粉(50%)、小麥粉(50%)和綠豆(42.9%)(表1)。

表1 不同谷類食品中克羅諾桿菌的污染率Table 1 Prevalence of Cronobacter isolates screened from various cereal samples
注:a:顯色平板法;b:16S rDNA測序結果;c:ND,未檢測(Not detected)。
2.2基于ITS基因的PCR驗證結果
經瓊脂糖凝膠電泳檢測,結果顯示24株可疑克羅諾桿菌分離株PCR檢測全部為陽性,在250 bp附近出現目的條帶,和預期片段大小(282 bp)相吻合,而陰性對照(雙蒸水替代基因組DNA)未擴增出目的片段(圖1)。
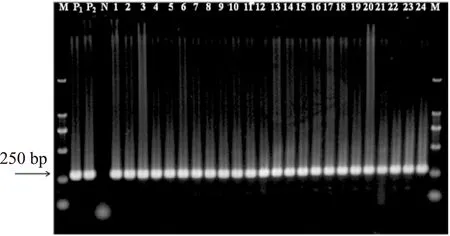
圖1 基于ITS基因的克羅諾桿菌分離株PCR檢測結果Fig.1 Detection of Cronobacter spp. isolated from cereal samples by PCR based on ITS gene注:Lane M:DL2000 DNA ladder;lane P1:陽性對照(C. sakazakii CICC 21560);lane P2:陽性對照(C. muytjensii CICC 21563);lane N:陰性對照;lanes 1~24:可疑克羅諾桿菌分離株。
2.3基于OmpA基因的PCR驗證結果
從瓊脂糖凝膠電泳圖譜可知:經PCR擴增后,有21株分離株能擴增出片段大小約為469 bp的目標基因片段(圖2),表明這些分離株可能為克羅諾桿菌。然而分離株N1(17)、分離株N2(19)和分離株N3(21)未擴增出目的片段,表明這些分離株可能不是克羅諾桿菌,分析可能的原因為基于ITS基因的PCR檢測方法產生了假陽性結果或基于OmpA基因的PCR檢測方法產生了假陰性結果。為了進一步驗證本研究獲得的分離株是否為克羅諾桿菌,本研究擴增了上述可疑分離株的16S rDNA基因片段,并對其進行測序驗證和BLAST在線比對分析。

圖2 基于OmpA基因的克羅諾桿菌 分離株PCR檢測結果Fig.2 Detection of Cronobacter spp. isolated from cereal samples by PCR based on OmpA gene注:Lane M:DL2000 DNA ladder;lane P1:陽性對照(C. sakazakii CICC 21560);lane P2:陽性對照(C. muytjensii CICC 21563);lane N:陰性對照;lanes 1~24:可疑克羅諾桿菌分離株。
2.4分離株16S rDNA測序結果
利用BLAST在線比對分析(http://blast.ncbi.nlm.nih.gov/Blast.cgi)分離株16S rDNA基因序列,結果顯示21株可疑分離株均為Cronobacterspp.,其中大部分分離株(17株)的16S rDNA基因序列與C.sakazakii的相似性較高,序列相似性達99%以上;3株分離株的16S rDNA基因序列與C.dublinensis的相似性最高,序列相似性達99%以上;1株分離株的16S rDNA基因序列與C.universalis和C.turicensis的相似性高達99%以上(表2)。由于克羅諾桿菌屬內不同種間菌株的16S rDNA基因序列相似性很高,通常無法通過16S rDNA基因序列比對的方法將其鑒定到種的水平,因而將本研究獲得的上述21株分離株初步鑒定為Cronobacterspp.。此外,BLAST比對分析結果表明分離株N1和N2為克雷伯菌屬(Klebsillaspp.),N3 為泛菌屬(Pantoeaspp.)(表2)。該結果與基于OmpA基因的PCR驗證結果一致,進一步證實了分離株N1、N2和N3不是克羅諾桿菌,同時也表明基于ITS基因的特異性PCR驗證方法產生了假陽性結果。

表2 克羅諾桿菌分離株形態學和分子生物學鑒定結果Table 2 Morphology and molecular identification of Cronobacter isolates screened from cereal samples
注:+,陽性;-,陰性。
2.5分離株系統發育分析
為了進一步分析克羅諾桿菌分離株的變異及系統進化關系,利用Mega 4.0軟件以16S rDNA基因序列為基礎構建系統發育樹。分析結果表明,21株分離菌株與克羅諾桿菌標準菌株(C.sakazakiiATCC 29544、C.malonaticusLMG 23826、C.turicensisLMG 23827、C.dublinensissubsp.lactaridiLMG 23823、C.dublinensissubsp.lactaridiLMG 23824、C.dublinensissubsp.lactaridiLMG 23825、C.universalisNCTC 9529和C.muytjensiiATCC 51329)位于同一簇群,親緣關系較近,而與其他腸桿菌科細菌的親緣關系較遠(圖4),進一步推斷本研究分離的上述21株可疑菌株為Cronobacterspp.。此外,分離株N1、N2和N3與克羅諾桿菌不在同一簇群,親緣關系較遠,進一步說明分離株N1、N2和N3不是克羅諾桿菌(圖3)。
3 結論與討論
采用準確可靠的檢測方法是實現對食品中克羅諾桿菌的檢測和監測的關鍵。常見的克羅諾桿菌分離鑒定方法主要有基于顯色平板的微生物培養法、基于特異性基因片段進行PCR鑒定方法等,但是這2種檢測方法往往會導致假陽性或假陰性結果[18]。因此,有必要結合16S rDNA基因序列分析方法、rpoB基因序列分析方法、脈沖場凝膠電泳(PFGE)和多位點序列分型(MLST)等方法對分離株進行進一步的驗證[19-20]。本研究采用GB 4789.40-2010分離了谷類食品中的克羅諾桿菌,并利用了基于ITS基因和OmpA基因的特異性PCR以及16S rDNA基因序列分析的方法對分離株進行了初步的鑒定,最終從100份谷類食品樣品中分離出21株克羅諾桿菌,其中17株被初步鑒定為C.sakazakii,3株為C.dublinensis,1株為C.universalis或C.turicensis。國內外關于食品中克羅諾桿菌污染狀況的調查結果表明食品中污染的克羅諾桿菌主要為C.sakazakii,與本研究的結果一致[21-22]。
不同種的克羅諾桿菌在致病性、耐藥性和耐干燥特性等方面具有較大差異。將分離株精確到種的水平對于追溯污染源、疾病的診斷和控制具有重要的意義。由于克羅諾桿菌不同種間菌株尤其是C.sakazakii和C.malonaticus之間的親緣關系較近,16S rDNA基因序列分析技術無法將其準確鑒定到種,可通過PCR-限制酶片段長度多態性(PCR-RFLP)、PFGE和MLST等基因分型方法予以解決[22-24]。
[1]Vojkovska H,Karpiskova R,Orieskova M,et al. Characterization ofCronobacterspp. isolated from food of plant origin and environmental samples collected from farms and from supermarkets in the Czech Republic[J]. International Journal of Food Microbiology,2016,217:130-136.
[2]董曉輝,李程思,吳清平,等.食品污染克羅諾桿菌(阪崎腸桿菌)的分離鑒定[J].微生物學報,2013,53(5):429-436.
[3]Caubilla-Barron J,Hurrell E,Townsend S,et al. Genotypic and phenotypic analysis ofEnterobactersakazakiistrains from an outbreak resulting in fatalities in a neonatal intensive care unit in France[J]. Journal of Clinical Microbiology,2007,45(12):3979-3985.
[4]Lai KK.Enterobactersakazakiiinfections among neonates,infants,children,and adults:Case reports and a review of the literature[J]. Medicine,2001,80(2):113-122.
[5]龔燕,顧學章. 阪崎腸桿菌肺炎 64 例臨床及藥敏分析[J]. 上海醫學,2005,28(12):1047-1048.
[6]Li Y,Chen Q,Zhao J,et al. Isolation,identification and antimicrobial resistance ofCronobacterspp. isolated from various foods in China[J]. Food Control,2014,37:109-114.
[7]陳萬義,任婧,吳正鈞,等. 生鮮蔬菜中阪崎克羅諾桿菌的分離與鑒定[J].食品科技,2014,39(1):304-308.
[8]Jung K,Park H. Prevalence and thermal stability ofEnterobactersakazakiifrom unprocessed ready-to-eat agricultural products and powdered infant formulas[J]. Food Science and Biotechnology,2006,15:152-157.
[9]Iversen C,Lane M,Forsythe J. The growth profile thermotolerance and biofilm formation ofEnterobactersakazakiigrown in infant formula milk[J]. Letters in Applied Microbiology,2004,38:378-382.
[10]Shaker R,Osaili T,Al-Omary W,et al. Isolation ofEnterobactersakazakiiand otherEnterobactersp from food and food production environments[J]. Food Control,2007,18:1241-1245.
[11]Jacobs C,Braun P,Hammer P. Reservoir and routes of transmission ofEnterobactersakazakii(Cronobacterspp.)in a milk powder-producing plant[J]. Journal of Dairy Science,2011,94(8):3801-3810.
[12]Lou X,Si G,Yu H,et al. Possible reservoir and routes of transmission ofCronobacter(Enterobactersakazakii)via wheat flour[J]. Food Control,2014,43:258-262.
[13]張翼,陳雅蘅,周幗萍,等. 克羅諾桿菌的生物膜檢測和藥敏性分析[J].食品科學,2015,36(21):129-134
[14]GB 4789.40-2010.食品微生物學檢驗 阪崎腸桿菌檢驗[S].北京:中國標準出版社,2010.
[15]Liu Y,Gao Q,Zhang X,et al. PCR and oligonucleotide array for detection ofEnterobactersakazakiiin infant formula[J]. Mol Cell Probe,2006,20:11-17.
[16]Nair M,Vekitanarayanan S. Cloning and sequencing of the
ompA gene ofEnterobactersakazakiiand development of an ompA-targeted PCR for rapid detection ofEnterobactersakazakiiin infant formula[J].Applied and Environmental Microbiology,2006,72:2539-2546.
[17]Weisburg G,Barns M,Pelletier A,et al. 16S ribosomal DNA amplification for phylogenetic study[J]. Journal of Bacteriology,1991,173(2):697-703.
[18]Jaradat Z,Ababneh Q,Saadoun I,et al. Isolation ofCronobacterspp.(formerlyEnterobactersakazakii)from infant food,herbs and environmental samples and the subsequent identification and confirmation of the isolates using biochemical,chromogenic assays,PCR and 16S rRNA sequencing[J]. BMC Microbiology,2009,9:225.
[19]Li Y,Cao L,Zhao J,et al. Use of rpoB gene sequence analysis for phylogenetic identification ofCronobacterspecies[J]. Journal of Microbiological Methods,2012,88:316-318.
[20]Joseph S,Sonbol H,Hariri S,et al. Diversity of theCronobactergenus as revealed by multilocus sequence typing[J]. Journal of Clinical Microbiology,2012,50(9):3031-3039.
[21]Lee D,Ryu W,Chang I,et al. Identification and classification ofCronobacterspp. isolated from powdered food in Korea[J]. Journal of Microbiology and Biotechnology,2010,20(4):757-762.
[22]Strydom A,Cameron M,Witthuhn R. PCR-RFLP analysis of the rpoB gene to distinguish the five species ofCronobacter[J]. Food Microbiology,2011,28:1472-1477.
[23]Molloy C,Cagney C,O’Brien S,et al. Surveillance and characterisation by pulsed-field gel electrophoresis ofCronobacterspp. in farming and domestic environments,food production animals and retail foods[J]. International Journal of Food Microbiology,2009,136(2):198-203.
[24]Cui J,Du X,Wei R,et al. Multilocus sequence typing analysis ofCronobacterspp. isolated from China[J]. Arch Microbiology,2015,197:665-672.
Isolation and identification ofCronobacterspp. from cereal foods
LI Yuan-hong1,ZHANG Yi-fei1,ZHANG Qing-cheng2,XU Wan1,ZOU Xiao-qian1,PEI Shang-fei1,ZHAO Min-sheng1,JIANG Hua1
(1.School of Public Health,Xuzhou Medical University,Xuzhou 221004,China;2.Department of Aviation Quartermaster,Air Force Logistics College,Xuzhou 221000,China)
To investigate the presence ofCronobacterspp. in cereal foods,a total of 100 cereal food samples were analyzed based on the national standard method of GB 4789.40-2010. The presumptive isolates was further identified by biochemical identification,PCR amplification of specific gene of ITS(internal transcribed spacer sequences)and OmpA,and sequence analysis of 16S rDNA. Twenty-one samples(21.0%)were positive forCronobacterspp.,and 21 isolates ofCronobacterwas collected in total. Most of theCronobacterisolates(81.0%)was identified asC.sakazakii,followed byC.dublinensis(14.2%),andC.universalisorC.turicensis(4.8%). The results indicated that commercially available cereal foods were possible reservoir ofCronobacterspp.,and therefore the epidemiological surveillance ofCronobacterspp. should be strengthened in cereal foods.
Cronobacterspp.;cereal foods;isolation;identification
2016-02-01
李遠宏(1984-),男,博士,講師,研究方向:食品微生物與生物技術,E-mail:lhy@xzmc.edu.cn。
國家自然科學基金青年科學基金項目(31401595);國家級大學生創新創業訓練計劃(20140313012);江蘇省高等學校大學生創新創業訓練計劃(201410313012Z)。
TS201.3
A
1002-0306(2016)15-0154-06
10.13386/j.issn1002-0306.2016.15.022
猜你喜歡
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
當代陜西(2019年7期)2019-04-25 00:22:18
領導決策信息(2018年26期)2018-10-12 02:18:26
海峽科技與產業(2016年3期)2016-05-17 04:32:12
Coco薇(2016年2期)2016-03-22 02:42:52
Coco薇(2015年1期)2015-08-13 02:47:34
小雪花·成長指南(2015年4期)2015-05-19 14:47:56
都市麗人(2015年5期)2015-03-20 13:33:49
