錳離子濃度及其轉運通道對水稻幼苗鎘吸收轉運特性的影響
2016-09-13 13:50:58楊益新李文華趙艷玲劉仲齊農業部環境保護科研監測所生態毒理與環境修復研究中心天津3009湖南省湘陰縣農業局湖南湘陰44600東北農業大學資源與環境學院哈爾濱50030
農業環境科學學報 2016年8期
關鍵詞:水稻
徐 莜,楊益新,李文華,陳 蕊,趙艷玲,唐 琦,3,劉仲齊*(.農業部環境保護科研監測所生態毒理與環境修復研究中心,天津 3009;.湖南省湘陰縣農業局,湖南 湘陰 44600;3.東北農業大學資源與環境學院,哈爾濱 50030)
污染生態
錳離子濃度及其轉運通道對水稻幼苗鎘吸收轉運特性的影響
徐莜1,楊益新2,李文華1,陳蕊1,趙艷玲1,唐琦1,3,劉仲齊1*
(1.農業部環境保護科研監測所生態毒理與環境修復研究中心,天津 300191;2.湖南省湘陰縣農業局,湖南 湘陰 414600;3.東北農業大學資源與環境學院,哈爾濱 150030)
為探明錳(Mn)及其轉運通道對水稻鎘(Cd)吸收轉運特性的影響,以水稻幼苗為材料進行水培試驗,對幼苗根系和地上部Cd、Mn的積累特征及其與P-型ATP酶和非選擇性陽離子通道(NSCCs)活性的關系進行研究。結果表明,0.8~1.6 mmol·L-1Mn能顯著降低水稻根系和地上部的Cd含量,并能提升水稻細胞壁中Cd的分配比率和降低胞液中Cd的分配比率。在含4.5 μmol·L-1Cd 和1.6 mmol·L-1Mn的營養液中,NSCCs對水稻根系和地上部Cd含量的貢獻率分別為34.9%和17.0%,P-型ATP酶的貢獻率分別為24.7%和7.6%;NSCCs對水稻根系和地上部Mn的貢獻率分別為24.0%和10.3%,明顯低于其對Cd的貢獻率,而P-型ATP酶對根系和地上部Mn的貢獻率分別為69.5%和20.3%,顯著高于其對Cd的貢獻率。綜上認為,Cd主要通過離子通道進入水稻根系細胞,Mn主要通過載體蛋白進入水稻根系細胞。Mn可能通過競爭離子通道和載體蛋白來抑制水稻幼苗對Cd的吸收和轉運。
鎘;錳;水稻;離子通道;載體蛋白
徐莜,楊益新,李文華,等.錳離子濃度及其轉運通道對水稻幼苗鎘吸收轉運特性的影響[J].農業環境科學學報,2016,35(8):1429-1435.
XU You,YANG Yi-xin,LI Wen-hua,et al.Effects of manganese concentrations and transporters on uptake and translocation of cadmium in rice seedlings[J]. Journal of Agro-Environment Science,2016,35(8):1429-1435.
近些年我國工農業發展迅猛,伴隨著工業“三廢”的大量排放,其中的部分Cd通過各種途徑進入農田土壤,致使全國土壤Cd點位超標率高達7%,農田Cd污染問題日益顯露和突出[1]。水稻是我國第一大糧食作物,農田土壤中大量Cd可被水稻根系富集,部分轉移至籽粒中,從而進入食物鏈積累于人體[2],對人體肝、腎、骨骼等組織造成嚴重的損害[3]。
Mn是人類與植物所必需的微量營養元素。在人體中,Mn主要參與脂肪代謝與造血過程,與鈣、磷代謝有關[4]。在水稻中,Mn參與光合作用中的放氧階段,負責水的光解過程,并且Mn與眾多酶促反應有關,是許多酶的活化劑[5]。此外,水稻還是一種高耐Mn作物。一些水稻品種即使葉片中Mn含量高達5000 mg· kg-1時,也不會出現Mn毒癥狀[6]。有研究表明,Mn2+具有緩解Cd對水稻的生長抑制作用,提高Mn2+濃度可顯著降低Cd在水稻根系中的含量[7]。
土壤中的Cd和Mn主要以離子形態進入水稻根系[8],一部分Cd2+與Mn2+吸附于根細胞壁或進入根細胞內,另一部分則通過共質體與質外體兩種運輸途徑,先橫向運輸至根系維管束組織,然后通過木質部縱向運輸至地上部組織[9-10]。水稻根系內外皮層間有一條木質化與栓質化加厚的凱式帶阻止根外溶液進入維管束,金屬離子需要依靠根細胞膜上的轉運通道進行跨膜運輸,才得以進入細胞內[11]。P-型ATP酶是一類位于質膜上利用ATP水解產生的能量使離子進行跨膜運輸的載體蛋白,負責主動轉運環境中微量的金屬元素,可同時參與Mn2+、Cd2+、Fe2+、Zn2+等多種陽離子的吸收轉運過程[12]。Laurent等[13]發現在擬南芥中,位于細胞膜上的P-型ATP酶家族成員AtHMA4可轉運Zn2+、Cd2+等多種陽離子,其序列N段具有富含半胱氨酸和組氨酸的重金屬結合域(MBDs),多種陽離子需競爭MBDs的結合位點,才得以通過AtHMA4進入胞內。釩酸鈉(Na3VO4)是一類能有效抑制植物細胞P-型ATP酶活性的抑制劑,用Na3VO4抑制P-型ATP酶的活性能顯著降低蘆蒿中Cd和Pb的積累量[14]。
水稻細胞膜上還存在一些能同時轉運多種離子的通道蛋白,它們主要是通過化學門控或電壓門控機制對離子進行被動運輸。如K+通道和非選擇性陽離子通道(NSCCs)等通道蛋白也能進行Cd2+的跨膜運輸[15-17]。NSCCs是泛指一類對于多種陽離子具有很低辨別力的離子通道集合體[18],Gd3+、La3+、奎寧、焦磷酸二乙酯等可顯著抑制NSCCs的活性[19]。
本研究以水稻幼苗為材料,對Mn2+濃度與水稻幼苗Cd、Mn積累及亞細胞分布特征的關系進行了研究,并應用P-型ATP酶抑制劑與NSCCs抑制劑對載體蛋白和離子通道在水稻Cd、Mn吸收轉運過程中的貢獻率進行分析,以期為應用Mn調控水稻Cd的吸收與積累提供理論依據。
1 材料與方法
1.1供試材料與培養條件
以早熟秈稻(Oryza sativa L.)品種T優705為供試材料,在人工氣候室內進行試驗。挑選均一飽滿的水稻種子,在裝有100 mL 5%NaClO溶液的器皿中浸泡20 min,用超純水反復清洗其上殘留的NaClO,洗凈后均勻地播撒于育苗盤上,在去離子水中進行第一階段培養。待水稻幼苗長至兩葉一心期,將其轉移至盛有1/10 Hoagland營養液的8 L培養箱中,進行第二階段培養。待水稻幼苗長至三葉一心期,挑選長勢一致的幼苗,在去離子水中緩苗1 d后,放置于以1/10 Hoagland營養液為基礎的不同處理液中進行5 d處理(由于1/10 Hoagland營養液自身含有約1 μmol·L-1MnCl2,處理液中實際Mn2+濃度為外源施加Mn2+濃度加營養液中自身Mn2+濃度)。水培實驗的整個過程均在人工氣候室內完成。人工氣候室條件參數:晝夜時間為16 h/8 h,晝夜溫度為25℃/20℃,白天光照105 μmol·m-2·s-1,相對濕度為60%[20]。
1.2培養與處理方法
設定處理液鎘(CdCl2)濃度為4.5 μmol·L-1,3個錳(MnSO4)水平設定為:0(Mn0)、0.8(Mn1)、1.6 mmol· L-1(Mn2)。GdCl3(NSCCs抑制劑)和Na3VO4(P-型ATP酶抑制劑)濃度均設定為0.1 mmol·L-1。處理液用HNO3和KOH調節pH至5.5~6.0。每個處理重復3次,每個重復中有15株水稻幼苗。
1.3樣品采集與處理
處理好的水稻幼苗先用5 mmol·L-1的CaCl2浸泡漂洗20 min,以去除根系表面吸附的Cd2+,再用去離子水將根系沖洗干凈。用剪刀將一部分水稻幼苗的根部與地上部分開,裝入信封,放入烘箱105℃殺青15 min,再置于75℃下烘干至恒重,用于全Cd含量的測定。另一部分直接用于亞細胞測定。
1.4Cd、Mn含量測定
Cd、Mn含量測定主要參照潘瑤等[21]的方法。用剪刀將烘至恒重后的樣品剪碎,稱取根系0.1 g、地上部0.25 g,將稱好后的樣品放入消煮管中,加入7 mLHNO3浸泡過夜。次日將消煮管放入消解儀ED54上,在110℃下消煮1.5 h。待消解液冷卻后,加入1 mL H2O2,再在150℃下消煮2.5h,最后取下管蓋,在180℃下趕酸至體積為0.5mL左右。消解完成后用去離子水將趕酸后的消解液轉移至25 mL容量瓶中并定容,用原子吸收光譜儀(AASZEEnit700)測定樣品Cd含量。
Cd的亞細胞分布方法主要依照Wang等[22]的方法。稱取新鮮的水稻幼苗根系或葉片0.5 g,置于研體中,加入20 mL預冷的提取緩沖溶液(250 mmol·L-1蔗糖,1 mmol·L-1二硫赤蘚醇,50 mmol·L-1Tris-HCl,pH7.4),將樣品充分研磨至勻漿,倒入50 mL離心管中。將裝有勻漿液的離心管放入4℃離心機中,3000 r·min-1離心15 min,沉淀即為細胞壁組分(F1)。將上清液轉入新50 mL離心管,15 000 r·min-1離心30 min,沉淀部分為細胞器組分(F2),上清液部分為胞液組分(F3)。將所得各組分轉移至消解管中后放入烘箱,待烘干水分后加酸消解,測定Cd和Mn含量。
1.5數據統計及分析
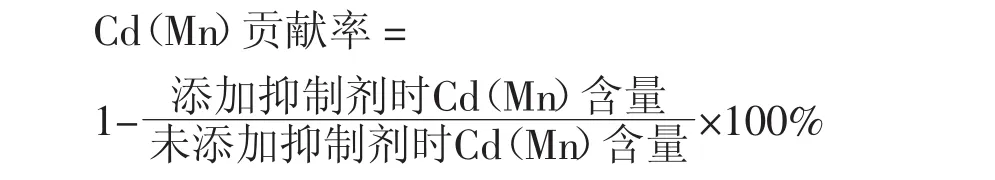
采用Excel與SPSS17.0軟件進行數據處理及顯著性分析,并用Origin8.6軟件作圖。
2 結果與分析
2.1Mn2+濃度和抑制劑對水稻幼苗Cd和Mn積累量的影響
Mn2+濃度對水稻根系和地上部Cd積累量有顯著影響。施加0.8 mmol·L-1和1.6 mmol·L-1外源Mn可使水稻根系中Cd的積累量分別下降18.4%和31.4%(圖1a),使地上部Cd的積累量分別下降25.7%和50.1%(圖1b)。NSCCs與P-型ATP酶抑制劑對水稻根系Cd積累量的影響比1.6 mmol·L-1外源Mn還明顯。0.1 mmol·L-1Gd3+和0.1 mmol·L-1VO3-4能使水稻根系的Cd積累量分別下降35.0%和24.7%,使地上部的Cd含量分別下降17.1%和7.6%。由此說明,增加根際環境Mn2+濃度和添加轉運通道抑制劑均有利于降低水稻幼苗根系與地上部對Cd的吸收與積累。
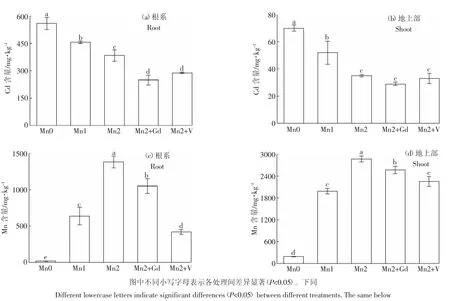
圖1 Mn和抑制劑對水稻幼苗Cd及Mn積累量的影響Figure 1 Effects of Mn and inhibitors on Cd concentrations and Mn concentrations of rice seedlings
水稻幼苗中Mn的含量隨處理液中Mn2+濃度的增加而大幅升高。施加0.8 mmol·L-1外源Mn使水稻根系Mn含量增加33倍(圖1c),使地上部Mn含量增加近9倍(圖1d);施加1.6 mmol·L-1Mn使水稻根系Mn含量增加72倍,地上部Mn含量增加14倍。0.1 mmol·L-1Gd3+能使水稻根系與地上部Mn含量分別下降24.0%和10.3%,0.1 mmol·L-1VO3-4能使水稻根系與地上部Mn含量分別下降69.5%和20.3%。這說明水稻對Mn具有很高的富集能力,其地上部的Mn含量顯著高于根系。
2.2Mn2+對水稻幼苗內Cd亞細胞分布的影響
水稻Cd主要分布于細胞壁(F1)與胞液(F3)兩部分中,細胞器(F2)中Cd含量很少。施加外源Mn可顯著降低水稻幼苗亞細胞各組分Cd含量,其中對F3組分的抑制效果最為顯著(圖2)。施加0.8 mmol·L-1和1.6 mmol·L-1外源Mn使水稻根系F1組分中Cd含量分別下降8.3%和20.0%,地上部F1中分別下降13.6%和 41.3%,根系 F2中分別下降 21.8%和58.5%,地上部F2中分別下降27.7%和49.8%,根系F3中分別下降23.6%和39.4%,地上部F3組分中分別下降41.1%和61.3%。
Mn不僅有助于降低水稻幼苗根系和地上部亞細胞組分中Cd的積累量,而且改變了各組分中Cd的分配比率。施加0.8~1.6 mmol·L-1外源Mn可使根系F1中Cd的分配比率提高4.7%~7.9%,使地上部F1 中Cd的分配比率提高8.5%~9.1%;與此同時,根系F3中Cd的分配比率下降了3.9%~4.7%,地上部F3 中Cd的分配比率下降了8.3%~9.2%。這說明Mn有助于提升Cd在細胞壁組分的積累,并減少Cd在胞液中的積累。
2.3NSCCs和P-型ATP酶對水稻幼苗Cd、Mn積累量的貢獻率
NSCCs和P-型ATP酶對水稻幼苗根系Cd和Mn積累量的貢獻率均大于對地上部積累量的貢獻率,且相互間存在顯著性差異(圖3)。NSCCs對水稻根系與地上部Cd貢獻率分別為34.9%和17.0%,均大于P-型ATP酶的24.7%和7.6%,說明Cd進入根系細胞以及向地上部分轉運的過程主要依靠離子通道來完成。NSCCs對水稻根系與地上部Mn貢獻率分別為24.0%和10.3%,而P-型ATP酶對水稻根系Mn積累量的貢獻率高達69.5%,是NSCCs貢獻率的2.9倍;P-型ATP酶對水稻地上部Mn積累量的貢獻率為20.3%,是NSCCs貢獻率的2.0倍。這說明Mn進入根系細胞和向地上部轉運的過程主要依靠載體蛋白,P-型ATP酶對水稻根系Mn的積累發揮了決定性作用。
3 討論
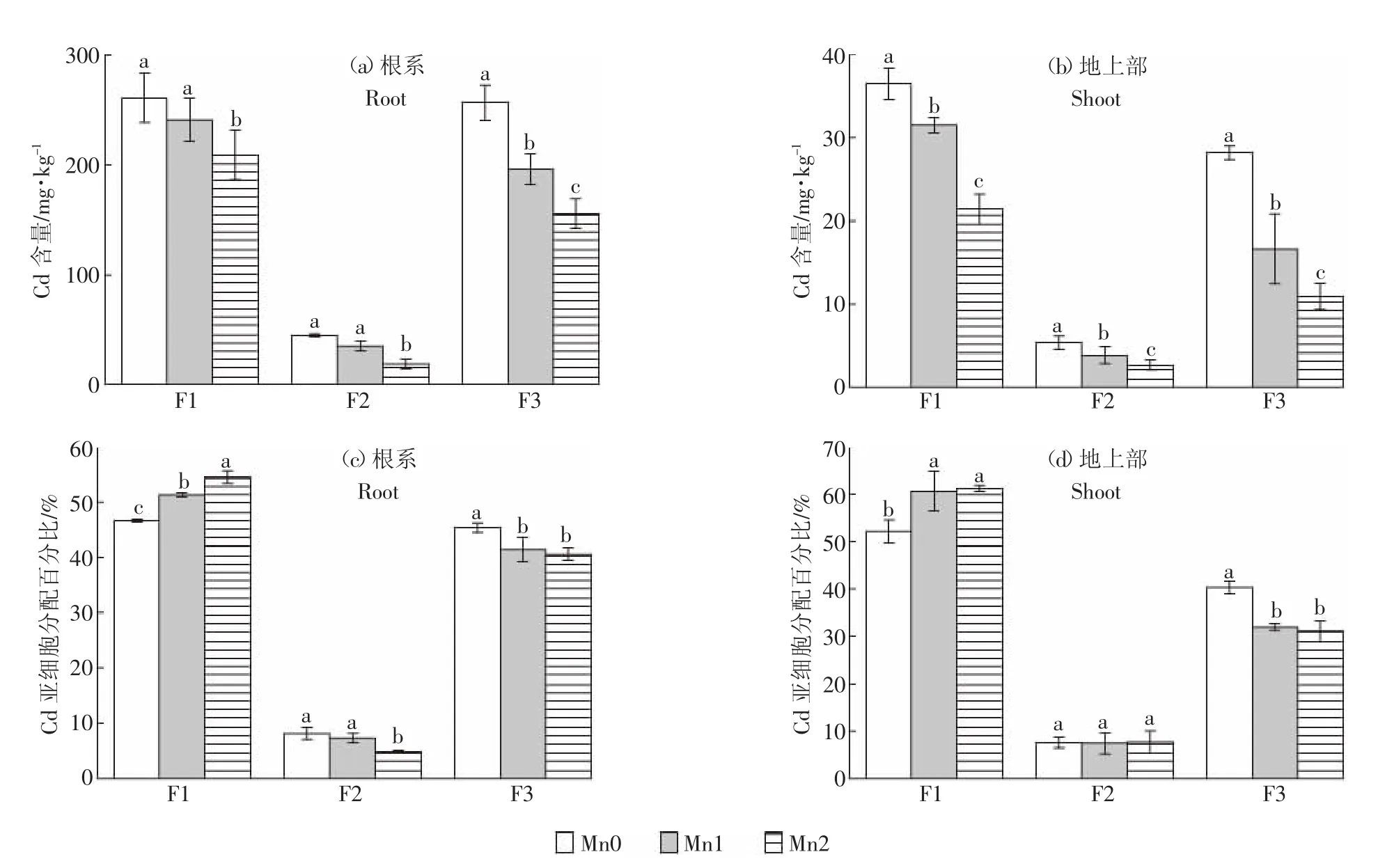
圖2 Cd水稻幼苗內的根系與地上部亞細胞分布及其亞細胞分布百分比Figure 2 Amount and percentages of Cd subcellular distribution in roots and shoots of rice seedlings
Mn是植物與人體必需的微量元素之一,應用其治理水稻Cd污染具有安全無污染的特點。本研究發現0.8~1.6 mmol·L-1外源Mn能顯著降低水稻根系與地上部Cd含量的18.4%~31.4%和25.7%~50.1%(圖1)。這與楊國治[23]發現Mn能抑制水稻對Cd吸收的規律一致,說明增加根際環境中的Mn能夠有效抑制Cd的吸收積累。
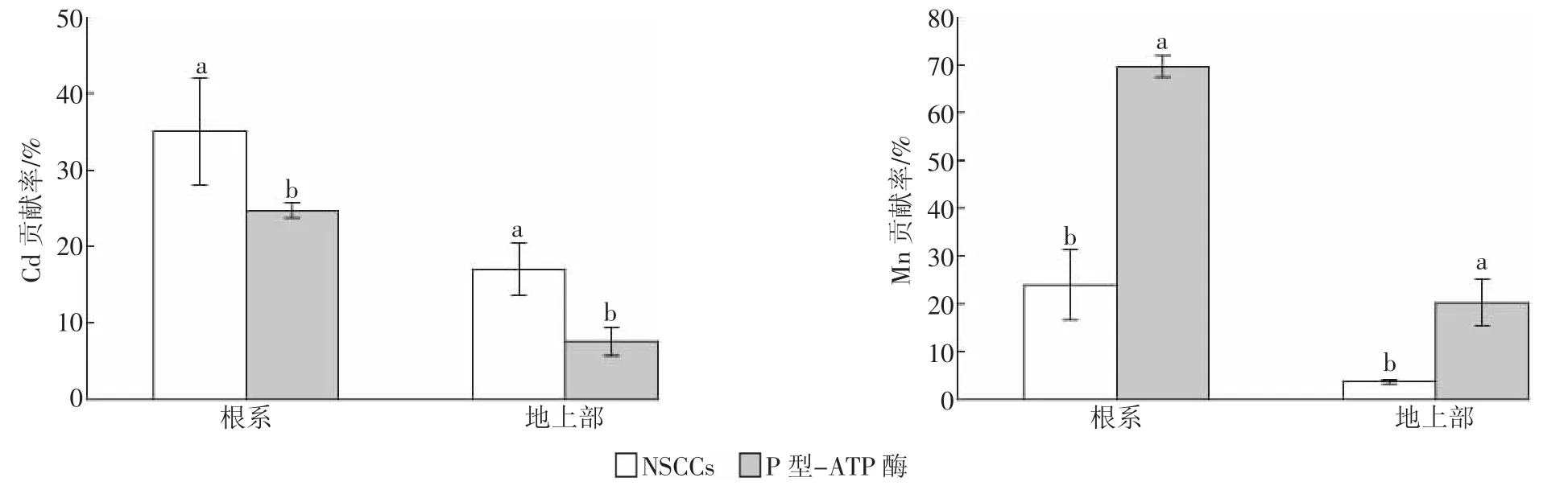
圖3 NSCCs與P型-ATP酶對水稻Cd和Mn積累量的貢獻率Figure 3 Percentages of NSCCs and P-ATPase contributions to Cd and Mn accumulation in rice seedlings
在水稻根際Mn2+隨水勢向根系遷移的過程中,水稻根系可將地上部吸收的O2通過疏導組織輸送到根系并釋放到環境中,使根際的Mn2+發生氧化,形成高價態Mn氧化物膠膜附著于根表,這部分膠膜對阻控Cd具有一定作用[24-26]。此外,水稻細胞壁中含有多種能與Cd產生螯合作用的糖類與蛋白質,有助于降低Cd的移動性,部分Mn2+進入根系質外體途徑后被輸送的O2氧化,形成的高價態Mn氧化物附著于細胞壁,增強了細胞壁對Cd的固定[27]。在本研究中,加入外源Mn后細胞壁組分中Cd含量顯著降低,但細胞壁組分中鎘的分配比率顯著升高(圖2)。這很可能是由于Mn2+濃度的提升增加了根表Mn膠膜厚度與細胞壁固持作用所致。
Cd2+與Mn2+均為二價陽離子,表現出相似的理化性質。因此,植物吸收轉運Cd2+與Mn2+常常涉及相同的運輸蛋白,它們對相關離子通道和載體蛋白的結合存在競爭關系[28]。本研究發現,加入外源Mn后胞液組分中Cd含量銳減,并且胞液組分中Cd的分配比率顯著降低,表明在水稻細胞中Mn2+和Cd2+對相關轉運蛋白的結合存在顯著的拮抗作用。胞液是維持植物細胞生命活動的重要場所,也是對Cd毒害最為敏感的部分之一,其組分中Cd含量的降低會減緩Cd對細胞代謝造成的毒害。
水稻中絕大部分Mn位于葉綠體,參與光合作用過程,地上部是水稻進行光合作用的主要場所[10],水稻中含有大量的載體蛋白,例如OsNRAMP5[28]、Os-NRAMP3[29]、OsYSL2[30]、OsMTP8.1[31]等,可將根系中的Mn2+轉運至維管束后運輸至地上部。非選擇性陽離子通道和金屬耐性蛋白(Metal tolerance protens,MTPs)等通道蛋白普遍存在于植物原生質、液泡膜和其他內膜系統中,對陽離子的選擇性很低,既可以轉運Mn2+和Ca2+、Mg2+、Zn2+等植物必需的營養元素,同時也參與Cd2+進入水稻體內的過程[32-34]。載體蛋白和通道蛋白的共同作用,保證了植物各種代謝活動對Mn的需求。而Cd作為植物的非必需元素,至今未在水稻中發現專屬于Cd的載體蛋白。Cd更多是通過對離子專一性較低的一些離子通道進入水稻胞液。例如,轉運Mn和Ca的載體蛋白及離子通道幾乎都能轉運Cd[34]。本研究發現,NSCCs對Cd積累量的貢獻率顯著大于P-型ATP酶,而P-型ATP酶對水稻根系和地上部Mn積累量的貢獻率顯著大于對Cd的貢獻率(圖3)。這說明Cd主要通過非選擇性陽離子通道進入水稻根系細胞,而Mn主要通過載體蛋白進入根系細胞和向地上部轉運。因此,在Mn和Cd共存的條件下,Mn2+通過優先結合細胞膜上的載體蛋白和通道蛋白對Cd2+產生拮抗作用,從而抑制Cd2+的跨膜運輸,減少根系細胞胞液中的Cd含量和向地上部的轉運量,緩解Cd2+的生理毒害作用。
4 結論
(1)增加根際Mn2+濃度能顯著升高水稻中Mn含量和降低Cd含量,1.6 mmol·L-1Mn能使水稻根系與地上部的Cd積累量分別下降31.4%和50.1%。
(2)水稻幼苗細胞壁與胞液是富集Cd的主要組分,細胞器組分中Cd含量很少。施加外源Mn有助于提高水稻細胞壁中Cd的分配比率和降低細胞液中Cd的分配比率。
(3)Cd主要通過非選擇性陽離子通道進入水稻根系細胞,而Mn主要通過載體蛋白進入水稻根系細胞。Mn通過競爭離子通道和載體蛋白來抑制水稻幼苗對Cd的吸收和轉運。
[1]Liu Z Q.Research advance on the mechanism of cadmium transport in rice[J].Meteorological and Environmental Research,2014,5(5):48-52.
[2]宋建,金鳳媚,薛俊,等.鎘脅迫對植物生長及生理生態效應的研究進展[J].天津農業科學,2014,20(12):19-22. SONG Jian,JIN Feng-mei,XUE Jun,et al.Advances of cadmium stress on plants growth and physiological and ecological effects[J].Tianjin Agricultural Sciences,2014,20(12):19-22.
[3]Méndez-Armenta M,Ríos C.Cadmium neurotoxicity[J].Environmental Toxicology and Pharmacology,2007,23(3):350-358.
[4]牛蕓民,楊天林.若干重要微量金屬元素的生物化學功能及其與人體健康的關系[J].微量元素與健康研究,2014,31(2):78-80. NIU Yun-min,YANG Tian-lin.A number of important biochemical function of trace metal elements and its relationship to human health[J]. Studies of Trace Elements and Health,2014,31(2):78-80.
[5]Williams L E,Pittman J K.Dissecting pathways involved in manganese homeostasis and stress in higher plant cells[J].Plant Cell Monographs,2010,17:95-117.
[6]Vlamis J,Williams D E.Iron and manganese relations in rice and barley[J]. Plant and Soil,1964,20(2):221-231.
[7]覃都,陳銘學,周蓉,等.錳-鎘互作對水稻生長和植株鎘、錳含量的影響[J].中國水稻科學,2010,24(2):189-195. QIN Du,CHEN Ming-xue,ZHOU Rong,et al.Effects of interaction between manganese and cadmium on plant growth and contents of cadmium and manganese in rice[J].Chinese Journal Rice Science,2010,24 (2):189-195.
[8]Marschner H.Mineral nutrition of higher plants[J].Mineral Nutrition of Higher Plants,1995,46(1):681-861.
[9]Uraguchi S,Mori S,Kuramata M,et al.Root-to-shoot Cd translocation via the xylem is the major process determining shoot and grain cadmium accumulation in rice[J].Journal of Experimental Botany,2009,60(9):2677-2688.
[10]Pittman J K.Managing the manganese:Molecular mechanisms of manganese transport and homeostasis[J].New Phytologist,2005,167(3):733-742.
[11]Kobayashi T,Nishizawa N K.Iron uptake,translocation,and regulation in higher plants[J].Annual Review of Plant Biology,2012,63(3):131-152.
[12]張玉秀,張媛雅,孫濤,等.植物重金屬轉運蛋白P1B-ATPase結構和功能研究進展[J].生物工程學報,2010,26(6):715-725. ZHANG Yu-xiu,ZHANG Yuan-ya,SUN Tao,et al.Structure and function of heavy metal transporter P1B-ATPase in plant:A review[J]. Chinese Journal of Biotechnology,2010,26(6):715-725.
[13]Laurent C,Lekeux G,Ukuwela A A,et al.Metal binding to the N-terminal cytoplasmic domain of the PIB ATPase HMA4 is required for metal transport in Arabidopsis[J].Plant Molecular Biology,2016.doi:10.1007/S11103-016-0429-2
[14]薛艷,王超,王沛芳,等.外源添加抑制劑對蘆蒿吸收Cd和Pb的影響[J].農業環境科學學報,2009,28(11):2255-2258. XUE Yan,WANG Chao,WANG Pei-fang,et al.Effect of additive inhibitor on the Cd and Pb uptake of Artemisia seleirgensis[J].Journal of Agro-Environment Science,2009,28(11):2255-2258.
[15]Zhou Y,Xia X M,Lingle C J.Cadmium-cysteine coordination in the BK inner pore region and its structural and functional implications[J]. Proceedings of the National Academy of Sciences,2015,112(16):5237-5242.
[16]王玉倩,汪曉麗,單玉華,等.環境因素對小麥根系非選擇性陽離子通道(NSCCs)轉運鉀的影響[J].植物營養與肥料學報,2010,16 (4):913-917. WANG Yu-qian,WANG Xiao-li,SHAN Yu-hua,et al.Effects of environmental factors on nonselective cation channels-mediated potassium uptake of wheat roots[J].Plant Nutrition and Fertilizer Science,2010,16(4):913-917.
[17]張參俊,尹潔,張長波,等.非選擇性陽離子通道對水稻幼苗鎘吸收轉運特性的影響[J].農業環境科學學報,2015,34(6):1028-1033. ZHANG Shen-jun,YIN Jie,ZHANG Chang-bo,et al.Effects of nonselective cation channels on accumulation and transfer of Cd in rice seedlings[J].Journal of Agro-Environment Science,2015,34(6):1028-1033.
[18]Zhang W H,Skerrett M,Walker N A,et al.Nonselective currents and channels in plasma membranes of protoplasts from coats of developing seeds of bean[J].Plant Physiology,2002,128(2):388-399.
[19]戴松香,陳少良.植物根細胞離子通道研究進展[J].北京林業大學學報,2005,27(3):98-103. DAI Song-xiang,CHEN Shao-liang.Research review on root ion channels of plants[J].Journal of Beijing Forestry University,2005,27(3):98-103.
[20]陸仲煙,劉仲齊,宋正國,等.大麥中鎘的亞細胞分布和化學形態及PCs合成的基因型差異[J].農業環境科學學報,2013,32(11):2125-2131. LU Zhong-yan,LIU Zhong-qi,SONG Zheng-guo,et al.Subcellular distribution and chemical forms of Cd and the synthesis of phytochelatins (PCs)in different barley genotypes[J].Journal of Agro-Environment Science,2013,32(11):2125-2131.
[21]潘瑤,尹潔,高子平,等.硫對水稻幼苗鎘積累特性及亞細胞分布特征的影響[J].農業資源與環境學報,2015,32(3):275-281. PAN Yao,YIN Jie,GAO Zi-ping,et al.Effects of sulfur on the accumulation and subcellular distribution of cadmium in rice seedlings[J]. Journal of Agricultural Resources and Environment,2015,32(3):275-281.
[22]Wang X,Liu Y G,Zeng G M,et al.Subcellular distribution and chemical forms of cadmiun in Bechmeria nivea(L.)Gaud[J].Environmental and Experimental Botany,2008,62(1):389-395.
[23]楊國治.錳能抑制水稻對鎘的吸收[J].土壤,1986,57(1):77-80. YANG Guo-zhi.Manganese can inhibit the uptake of cadmium in rice[J].Soil,1986,57(1):77-80.
[24]Wang M Y,Chen A K,Wong M H,et al.Cadmium accumulation in and tolerance of rice(Oryza sativa L.)varieties with different rates of radial oxygen loss[J].Environmental Pollution,2011,159(6):1730-1736.
[25]Liu W J,Zhu Y G,Smith F A.Effects of iron and manganese plaques on arsenic uptake by rice seedlings(Oryza sativa L.)grown in solution culture supplied with arsenate and arsenite[J].Plant and Soil,2005,277 (1/2):127-138.
[26]王丹,李鑫,王代長,等.硫素對水稻根系鐵錳膠膜形成及吸收鎘的影響[J].環境科學,2015,36(5):65-76. WANG Dan,LI Xin,WANG Dai-zhang,et al.Effects of nonselective cation channels on accumulation and transfer of Cd in rice seedlings[J]. Journal of Agro-Environment Science,2015,36(5):65-76.
[27]Fu X,Dou Chen Y,et al.Subcellular distribution and chemical forms of cadmium in Phytolacca Americana L.[J].Journal of Hazard Material,2011,186(1):103-170.
[28]Ishimaru Y,Takahashi R,Bashir K,et al.Characterizing the role of rice NRAMP5 in manganese,iron and cadmium transport[J].Scientific Reports,2012.doi:10.1038/Srep00286.
[29]Yamaji N,Sasaki A,Xia J X,et al.A node-based switch for preferential distribution of manganese in rice[J].Nature Communications,2013,4(9):2442-2442.
[30]Yasuhiro I,Hiroshi M,Khurram B,et al.Rice metal-nicotianamine transporter,OsYSL2,is required for the long-distance transport of iron and manganese[J].Plant Journal for Cell&Molecular Biology,2010,62 (3):379-390.
[31]Chen Z,Fujii Y,Yamaji N,et al.Mn tolerance in rice is mediated by MTP8.1,a member of the cation diffusion facilitator family[J].Journal of Experimental Botany,2013,64(14):4375-4387.
[32]Emmanuel D,Gruber B D,Pittman J K,et al.A role for the AtMTP11 gene of Arabidopsis in manganese transport and tolerance[J].Plant Journal for Cell&Molecular Biology,2007,51(2):198-210.
[33]劉勝浩,劉晨臨,黃曉航,等.植物細胞的非選擇性陽離子通道[J].植物生理學通訊,2006,42(3):523-528. LIU Sheng-hao,LIU Chen-lin,HUANG Xiao-hang,et al.Nonselective cation channels in plant cells[J].Plant Physiology Communications,2006,42(3):523-528.
[34]Pinto E,Isabel M P L V O.Ferreira.Cation transporters/channels in plants:Tools for nutrient biofortification[J].Journal of Plant Physiology,2015,179:64-82.
Effects of manganese concentrations and transporters on uptake and translocation of cadmium in rice seedlings
XU You1,YANG Yi-xin2,LI Wen-hua1,CHEN Rui1,ZHAO Yan-ling1,TANG Qi1,3,LIU Zhong-Qi1*
(1.Centre for Research in Ecotoxicology and Environmental Remediation,Agro-Environmental Protection Institute,Ministry of Agriculture,Tianjin 300191,China;2.Xiangyin Bureau of Agriculture,Xiangyin 414600,China;3.School of Resources and Environmental Sciences,Northeast Agricultural University,Harbin 150030,China)
Heavy metal uptake by plants is influenced by coexisting ions.In this study,the effects of manganese(Mn)and its transporters on uptake and translocation of cadmium(Cd)in rice seedlings,the accumulation characteristics of Cd and Mn as well as their relationships with nonselective cation channels(NSCCs)and P-ATPase were studied under hydroponic conditions.Experimental results showed that at 0.8 mmol·L-1to 1.6 mmol·L-1of solution Mn,Cd accumulations were reduced both in roots and shoots,but the percentages of Cd distribution in the cell wall was increased,while that in solube fraction was decreased.In the solution with 4.5 μmol Cd·L-1and 1.6 mmol Mn·L-1,the percentages of NSCCs contributions to the accumulation of Cd in roots and shoots were 34.9%and 17.0%,and those of P-ATPase were 24.7% and 7.6%,respectively.However,the percentages of NSCCs contributions to the accumulation of Mn in roots and shoots were 24.0%and 10.3%,those of P-ATPase to the accumulation of Mn in roots and shoots were 69.5%and 20.3%,respectively.These results indicate that Cd is mainly transported into root cells through nonselective cation channels,while Mn is primarily carried into root cells via positive transporters.Manganese may inhibit Cd uptake and translocation in rice seedlings by competing ionic channels and carrier proteins.
cadmium;manganese;rice;ionic channel;carrier protein
X171.5
A
1672-2043(2016)08-1429-07
10.11654/jaes.2016-0133
2016-01-26
2015年中國農科院科技創新工程項目(2015-cxgc-lzq);公益性行業(農業)科研專項(201403015)
徐莜(1992—),男,北京海淀人,碩士研究生,主要從事植物營養學研究。E-mail:xuyou406@126.com
劉仲齊E-mail:liuzhongqi508@163.com
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
