類蘆對鉛鎘的吸收動力特性及亞細胞分布規律研究
2016-09-13 13:51:00羅潔文黃玫英殷丹陽吳鵬飛蔡麗平周垂帆福建農林大學林學院福州350002海峽兩岸紅壤區水土保持協同創新中心福州350002
農業環境科學學報 2016年8期
關鍵詞:植物
羅潔文,黃玫英,殷丹陽,吳鵬飛,蔡麗平,周垂帆(福建農林大學林學院,福州350002;海峽兩岸紅壤區水土保持協同創新中心,福州 350002)
類蘆對鉛鎘的吸收動力特性及亞細胞分布規律研究
羅潔文,黃玫英,殷丹陽,吳鵬飛,蔡麗平,周垂帆*
(福建農林大學林學院,福州350002;海峽兩岸紅壤區水土保持協同創新中心,福州 350002)
以南方水土保持植物類蘆為研究對象,采用營養液培養的方法,以Pb、Cd為目標污染物,設置不同的脅迫濃度和時間,脅迫后測定類蘆體內重金屬的含量和亞細胞分布。結果分析發現,類蘆體內重金屬含量與濃度和時間有顯著的關系,可利用Freundlich方程和Michaelis-Menten方程進行擬合,R2>0.95,相關性強。通過擬合發現,重金屬主要分布在根部,類蘆對Pb的吸收能力更強(最高含量達到4 687.87 mg·kg-1),但是對Cd的吸收速率增長更為明顯,即類蘆在吸收Cd方面有一定的潛力。就亞細胞分布而言,兩種重金屬均主要分布在細胞壁和可溶組分中,不同濃度和時間處理下,兩個組分中的重金屬含量共占總量的60%以上。由此說明,類蘆對Pb的吸收能力更強,同時,類蘆通過改變重金屬的亞細胞分布來降低重金屬的毒害作用,維持自身的穩態。
類蘆;Pb;Cd;吸收動力學;亞細胞分布
羅潔文,黃玫英,殷丹陽,等.類蘆對鉛鎘的吸收動力特性及亞細胞分布規律研究[J].農業環境科學學報,2016,35(8):1451-1457.
LUO Jie-wen,HUANG Mei-ying,YIN Dan-yang,et al.Uptake kinetic characteristics and subcellular distribution of Pb2+and Cd2+in Neyraudia reynaudiana [J].Journal of Agro-Environment Science,2016,35(8):1451-1457.
重金屬污染日趨嚴重,重金屬污染的治理逐漸引起了人們的重視。其中,Pb、Cd是植物體內的非必需元素,這兩種重金屬毒性極強,Pb或Cd脅迫至一定的濃度則可以使植物出現中毒癥狀,如葉片失綠、生長遲緩、光合作用被抑制、擾亂植物對礦質營養元素的吸收等[1-2]。植物的防御主要通過外排作用避免重金屬進入細胞,或是通過細胞壁對重金屬進行沉淀、區隔化,同時通過一些配位基團、有機酸、氨基酸以及植物螯合物與重金屬作用,在其進入細胞前減輕其毒害,不同的植物對Pb、Cd的響應過程和耐性機制不同[3-4]。植物修復是近年來新興的綠色修復技術,其中一些生物量大、生長速度快、對污染物吸收能力較強但還達不到超富集植物標準的耐性植物受到了廣泛的關注。
類蘆(Neyraudia reynaudiana)是中國南方優良的水土保持防風固土的綠化植物,不僅可在重金屬礦廢棄地上較好生長,且保持有較高的生物量,在重金屬廢棄地的惡劣環境下表現出較強的適應性,其耐干旱、貧瘠、酸、重金屬等,根系發達(可長達300 cm),莖分蘗力強,生長快,生物量大(高可達300 cm左右,直徑約1 cm),能適應各種惡劣的環境,這些特性使類蘆成為能在鉛鋅礦上生長的幾種植物之一[5]。礦山廢棄地研究[6]發現其地上部分Pb含量最高可達773 mg·kg-1,接近Pb超積累植物的臨界含量標準(1000 mg·kg-1),且其生物富集系數0.19~1.69,從轉運系數來看,類蘆的轉運系數為3.1~21.2。除此之外,研究[7]發現在添加EDTA螯合劑的情況下,類蘆地上部分對Pb的富集量可高達11 008 mg·kg-1,是對照的16.5倍。正是類蘆具備的這些特點,使其成為適合南方Pb、Cd重金屬污染的修復植物。近年來,國內外學者對類蘆開展的研究大多集中在水土流失治理、根系固土作用以及其對干旱和養分脅迫的響應等方面[8-9]。但有關類蘆對重金屬耐性的機理并不清楚,這成為限制類蘆在重金屬污染修復應用的主要技術瓶頸。鑒于此,本文選取南方水土保持先鋒植物類蘆為試驗材料,采用營養液培養的方法,研究不同Pb、Cd處理濃度、處理時間下類蘆對Pb、Cd的吸收動力學特性,并對Pb、Cd在各亞細胞組分的分布進行分析。本研究旨在探明類蘆對Pb、Cd的吸收動力學規律,揭示Pb、Cd在類蘆細胞內的分布格局,為闡明類蘆對Pb、Cd污染的耐性和富集機理研究提供基礎數據。
1 材料與方法
1.1試驗材料
類蘆種子于2015年4月購自云南省昆明市某種子公司。同年8月底種植于福建農林大學溫室大棚,10月底在水培實驗前,選擇生長均一的類蘆幼株轉移到培養箱中,移植在泡沫板上并放置于500 mL超純水中培養3~4 d,然后依次用1/2和完全Hoagland培養液在塑料盆中培養3~4 d。培養期間晝/夜溫度為25℃/22℃,光照時間16 h。
1.2試驗方法
1.2.1Pb2+、Cd2+吸收濃度動力學
取成長到一定程度大小一致的類蘆苗,依次用自來水和去離子水沖洗干凈,用含2 mmol·L-1Mes-Tris和0.5 mmol·L-1CaCl2的預處理液(用HCl和NaOH調節pH值為5.8)預培養12 h后,放入含有不同濃度Pb2+/Cd2+[以Pb(NO3)2/CdCl2的形式加入]的2 mmol·L-1Mes-Tris、0.5 mmol·L-1CaCl2(pH5.5)吸收液中培養48 h,濃度分別為0.5、1、5、10、25、50、100、200 μmol·L-1(為避免產生磷酸鉛沉淀,營養液中KH2PO4的濃度設置為0.25 mmol·L-1),每個處理設置3次重復,每個處理重復4棵類蘆。類蘆苗處理48 h后取出,依次用自來水和去離子水沖洗根部,放入含有2 mmol·L-1Mes-Tris和5 mmol·L-1CaCl2(pH5.5)的洗脫液中冰水浴30 min。洗脫后的根用去離子水沖洗干凈,吸水紙擦干,分為根部和地上部分,置于烘箱105℃殺青30 min,然后75℃烘干72 h,稱重,測定Pb2+/Cd2+含量。選取Pb2+/Cd2+的濃度分別為0、5、25、200 μmol·L-1處理的鮮樣作亞細胞組分分析,每個處理3次重復。
1.2.2Pb2+、Cd2+吸收時間動力學
基于預試驗的結果,在Pb2+/Cd2+濃度為25 μmol· L-1時能夠正常生長,選取Pb2+/Cd2+濃度為25 μmol·L-1進行時間動力學試驗。將類蘆預培養12 h后,置于Pb2+/Cd2+濃度為25 μmol·L-1的吸收液中(含有2 mmol· L-1Mes-Tris、0.5 mmol·L-1CaCl2,pH5.5)培養3、6、12、24、48 h;試驗設置4次重復,每個處理重復6棵類蘆。吸收培養后,洗脫,洗凈,擦干,烘干,稱重,測定Pb2+/Cd2+含量;并取鮮樣作亞細胞組分分析,每個處理3次重復。
1.3樣品Pb2+、Cd2+總含量的測定
把地上部分和地下部分烘干至恒量,將干燥的類蘆根或類蘆葉子粉碎后準確稱取0.2 g(精確到0.000 1)于50 mL三角瓶中,加混酸(HNO3∶HClO4=5∶1)10 mL,在電爐上加熱至冒白煙,用1%的稀硝酸定容至25 mL容量瓶中,然后用原子吸收法測定Pb2+/Cd2+含量。1.4樣品亞細胞組分Pb2+、Cd2+含量的測定
采用差速離心法分離不同的細胞組分,參考周小勇等[10]的方法,具體步驟為:準確稱取鮮樣0.2 g,加入20 mL提取液[0.25 mol·L-1蔗糖+50 mmol·L-1Tris-HCl緩沖液(pH 7.5)+1 mmol·L-1二硫赤鮮糖醇],液氮研磨勻漿,勻漿液在冷凍離心機300 r·min-1下離心1 min,沉淀為細胞壁組分(F1);上清液在2000 r·min-1下離心15 min,沉淀為細胞核和葉綠體組分(F2);上清液在10 000 r·min-1下離心20 min,沉淀為線粒體組分(F3);上清液為含核糖體的可溶組分(F4),全部操作在4℃下進行。各組分采用1.3節中的方法進行消解,定容至10 mL后測定Pb、Cd含量。
1.5數據處理
數據為3次重復樣品測定結果的平均值(含標準差)。運用Excel 2007和SPSS 18進行數據統計分析,采用單因素方差分析(One-way ANOVA)和多重比較,用SNK法對處理間數據進行差異顯著性分析,以P<0.05表示處理間差異顯著,并用Origin 9.2作圖。
2 結果與分析
2.1類蘆對Pb2+、Cd2+的吸收濃度動力學
在Pb、Cd處理下,隨著重金屬濃度的增加,類蘆葉片和根對Pb、Cd的吸收都呈明顯的增長趨勢。但是類蘆對Pb、Cd的吸收在200 μmol·L-1下,經過48 h的處理后仍未達到飽和。類蘆對Pb2+、Cd2+吸收動力學曲線呈線性、非飽和曲線,用Freundlich方程對類蘆葉片吸收Pb、Cd曲線進行擬合,修改后的方程為[11]:
Q=KfXn
式中:Q為吸收量,mg·kg-1;X為溶液中Pb2+、Cd2+的濃度,μmol·L-1;Kf為與吸收量有關的常數;n為吸收曲線斜率。Kf和n都由實驗測定分析得出。
由圖1可知,類蘆葉片吸收Pb2+、Cd2+可以采用Freundlich方程擬合,相關系數高。擬合得到的方程為:
Pb處理:Q=34.34X0.23
Cd處理:Q=2.91X0.44
擬合方程在坐標上的截距為Kf值,對于重金屬的吸收情況而言,類蘆吸收Pb的Kf值大于Cd的Kf值,說明類蘆在重金屬吸收中具有選擇性,相同濃度脅迫下,類蘆能更多地富集重金屬Pb。另外,n為曲線的斜率,在48 h的脅迫時間內,類蘆吸收Cd的n值大于Pb的n值,說明類蘆吸收Cd的速率增長得更快。
類蘆根系對Pb2+、Cd2+的吸收曲線(其中,Pb只對濃度0~100 μmol·L-1進行擬合)可利用Michaelis-Menten方程進行擬合[12]:
VC=Vmax×X/(Km+X)
式中:VC是吸收速率,mol·g-1·h-1;X是溶液中Pb2+、Cd2+的濃度,μmol·L-1;Vmax是最大吸收速率,mol·g-1· h-1;Km是方程的速率常數,μmol·L-1,Km值代表了根系對離子的親和力,該值越小,親和力越高。根吸收力α=Vmax/Km,表示根系對離子的實際吸收能力,α值越大,表明根系對離子的吸收能力越強。
根據方程的擬合效果(圖2)及參數(表1),相關系數R2>0.95,說明該方程能很好地擬合吸收曲線。類蘆根系對Pb2+、Cd2+的親和值Km相近,而最大吸收速率Vmax有較大的差異,類蘆吸收Cd的α值明顯高于Pb的α值,說明類蘆根系對Cd吸收能力更強。但是此吸收規律僅對于Pb濃度為0~100 μmol·L-1而言,當Pb濃度達到200 μmol·L-1,根系大量富集Pb,其含量達到4 687.86 mg·kg-1,說明類蘆對Pb有較高的耐受和富集能力。
2.2類蘆對Pb2+、Cd2+的吸收時間動力學
吸收液中Pb2+、Cd2+濃度為25 μmol·L-1時,類蘆對Pb2+、Cd2+的積累量都隨時間的增加而增加,并且都能用修改的Freundlich方程進行擬合,相關系數R2>0.95,擬合性好,見圖3。
由圖3可知,在吸收時間動力學試驗中(3~48 h),類蘆對Pb2+、Cd2+的吸收量基本保持平穩增長,根部隨時間增長大量富集Pb、Cd,處理48 h后,根部吸收的Pb、Cd分別是葉片的3.2倍和13.72倍。而由表2可知,類蘆吸收Pb的Kf值大于Cd的Kf值,說明在3~48 h內,類蘆能更多地富集重金屬Pb。但是在曲線斜率n上,類蘆吸收Cd的n值均大于Pb的n值,說明相對而言,類蘆吸收Cd的速率增長得更快、更明顯。
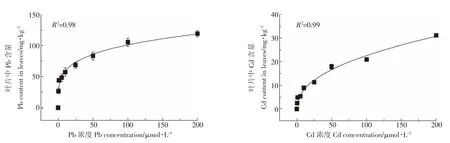
圖1 類蘆葉片對Pb2+、Cd2+的吸收速率與溶液濃度的關系Figure 1 Concentration-dependent Pb2+,Cd2+influx kinetics in leaves of Neyraudia reynaudiana
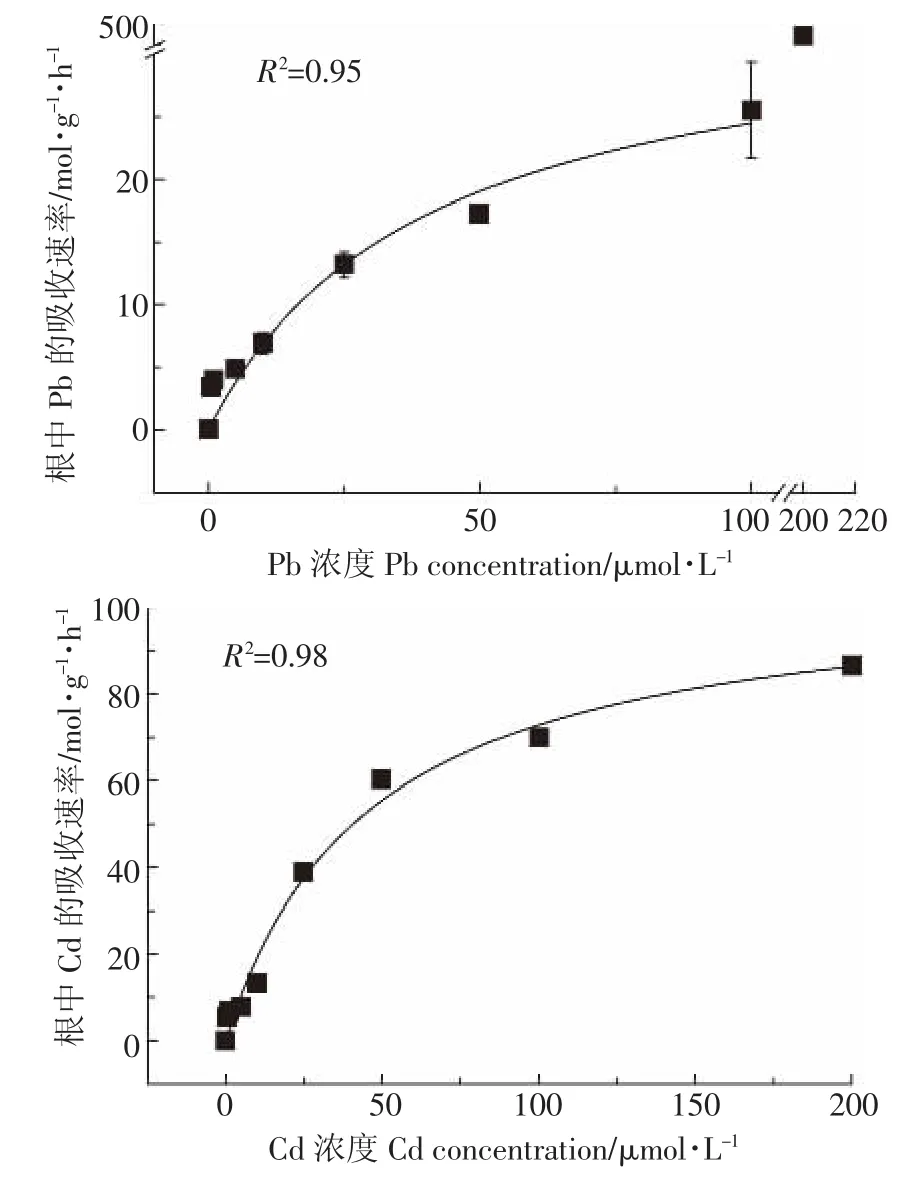
圖2 類蘆根系對Pb2+、Cd2+的吸收速率與溶液濃度的關系Figure 2 Concentration-dependent Pb2+,Cd2+influx kinetics in roots of Neyraudia reynaudiana

表1 Michaelis-Menten方程擬合參數Table 1 Fitting parameters for Michaelis-Menten equation
2.3Pb2+、Cd2+在類蘆體內的亞細胞分布
通過差速離心分離法,將類蘆葉片和根細胞分別分離出四部分,并對Pb2+、Cd2+在類蘆體內的分布進行測定,結果顯示,沒有Pb、Cd脅迫時,植物體內只有少量的Cd分布于細胞壁(可能是培育的土壤受到了輕微Cd污染),并且沒有檢測出Pb。從總量來說,根中的重金屬含量基本高于葉片中的含量,Pb在植物體內的含量高于Cd的含量,在不同Pb2+、Cd2+濃度脅迫時(表3),Pb、Cd在細胞壁組分(F1)和含核糖的可溶組分(F4)的分布最高,占61.98%~100%,其次為細胞核和葉綠體組分(F2),僅有少量分布在線粒體組分(F3),這說明細胞壁和含核糖體可溶組分是Pb、Cd在類蘆體內的主要分布位點,只有少量分布于細胞核和葉綠體以及線粒體等細胞器中,說明Pb、Cd的脅迫使細胞壁對重金屬的滯留作用顯著增強。同是類蘆葉片,Cd主要分布在F1中,在低濃度脅迫時,該組分中的Cd含量達到74.76%,但隨著Cd濃度的增加,部分金屬轉移到F4中,而Pb在類蘆體內的比例并沒有明顯的變化。根系狀況與葉片有所不同,高濃度Pb脅迫下,Pb在細胞壁中大量富集,Pb濃度為200 μmol· L-1時,在此組分的含量達到最高,為1 096.49 mg·kg-1,所占比例為90.43%,同時減少了Pb在其他組分中的分配比例,說明細胞壁為促進植物進行正常生理活動發揮了作用。而Cd在類蘆根部的分布,隨著Cd濃度的增加,其在F1組分中的比例下降,在200 μmol·L-1時降至44.63%,而F4的比例上升,說明部分Cd從F1組分轉移到F4組分中。

表2 Freundlich方程的擬合Table 2 Fitting parameters for Freundlich equation
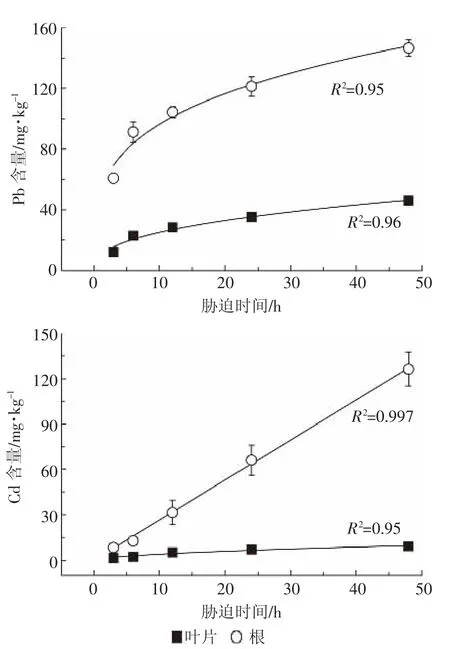
圖3 類蘆對Pb2+、Cd2+的吸收速率與時間的關系Figure 3 Time-dependent Pb2+,Cd2+influx kinetics in roots and leaves of Neyraudia reynaudiana
在Pb、Cd不同時間處理下(表4),類蘆吸收Pb的總量仍高于吸收Cd的量,植物體內F1與F4組分中Pb、Cd的含量隨著時間的增長而增加,Pb、Cd在這兩個組分中所占比例最高,占總量的59.88%~100%。在48 h處理時間內,Cd在F2、F3組分含量極低,儀器未能檢測出其含量,而Pb在F2和F3中的含量在類蘆不同部位變化趨勢不一。同在類蘆葉片,從Pb、Cd含量所占的百分比來看,Pb在F1組分中的比例下降而F4組分比例增加,而Cd則相反,隨著處理時間的增加,F1組分中從100%降至80.56%,F4組分從0%增加至19.44%。Cd在根系中的分布比例狀況與葉片中的相似,且含量與時間有顯著的關系(P<0.05),而Pb在根F1組分中的比例隨時間的增長先升后降,F4的比例呈上升趨勢,F2、F3組分所占比例呈下降趨勢。上述結果表明,隨著脅迫時間的增加,類蘆根細胞壁能將大部分Pb、Cd吸附在細胞壁上,并且類蘆對Pb、Cd的吸收具有一定的動態變化。
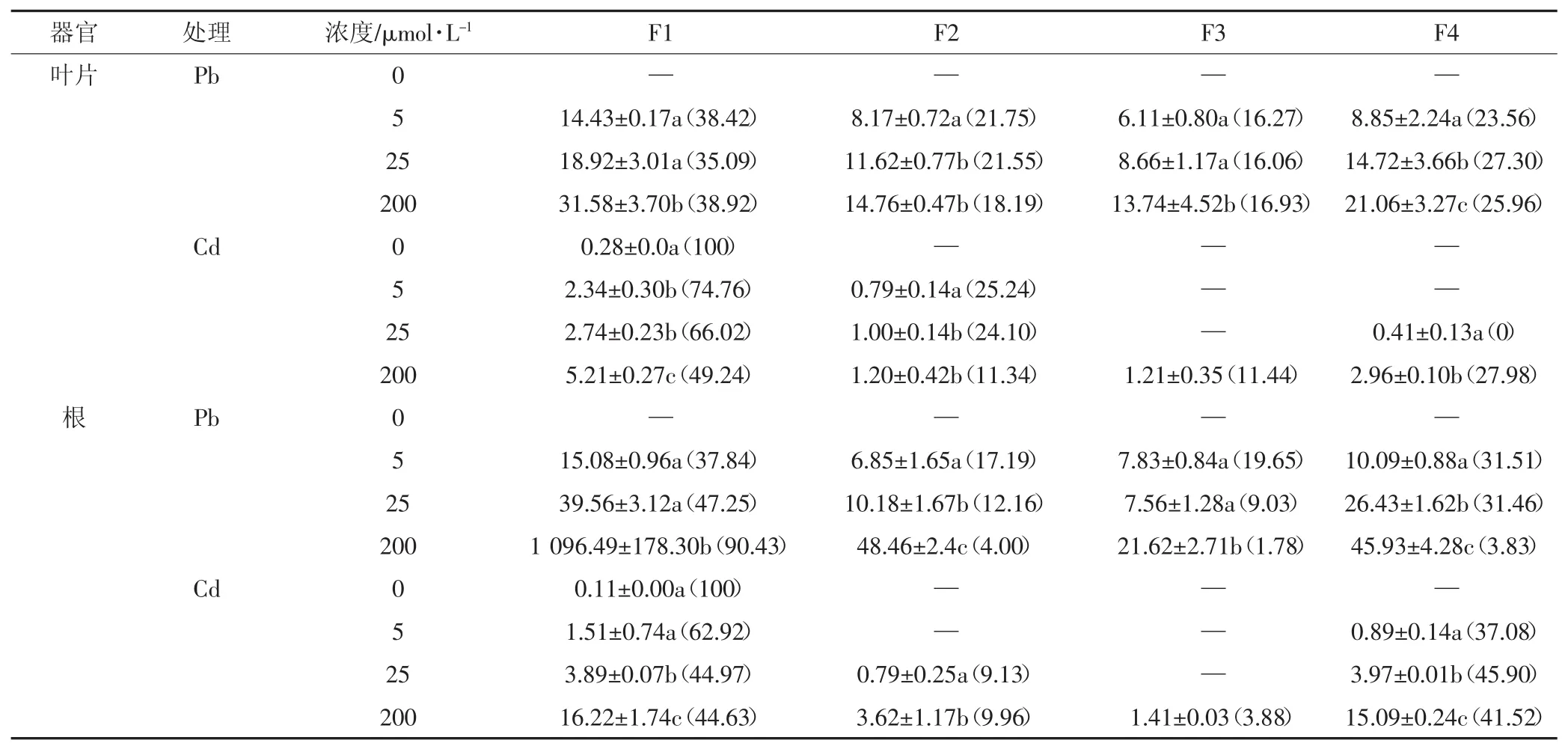
表3 不同濃度處理下Pb2+、Cd2+在類蘆體內的亞細胞分布(mg·kg-1)Table 3 Subcellular distribution of Pb2+and Cd2+in Neyraudia reynaudiana under different concentrations(mg·kg-1)
3 討論
3.1類蘆對Pb2+、Cd2+的吸收規律
一般而言,植物通過根際吸收重金屬元素,并將其輸送和富集到地上部分,在本研究中,類蘆體內重金屬含量與濃度和時間有顯著的相關性,類蘆根系吸收溶液中的重金屬,并轉移到葉片中。經過分析后發現,可利用Freundlich方程和Michaelis-Menten方程對吸收量和吸收速率進行擬合。從擬合方程對比來看,類蘆對Pb有更強的敏感性,吸附量更高,但對Cd的吸收速率更為顯著,類蘆吸收Cd有一定的潛力。類蘆根中可以積累較大量的Pb2+、Cd2+,并且這兩種重金屬濃度在根中均高于葉片中的濃度,這跟以往關于植物吸收重金屬的研究一致[13],可能是重金屬從根到地上部分遠距離轉運而導致的。當溶液中重金屬濃度達到200 μmol·L-1時,Cd在類蘆葉片中的含量為31.10 mg·kg-1,根中含量達到465.43 mg·kg-1,約為葉片含量的15倍;而Pb在類蘆體內的分布差異更為顯著,其在葉片中含量為118.65 mg·kg-1時,在根中大量富集,達到4 687.87 mg·kg-1,是葉片中含量的39.5倍。由此可見,水培條件下根系是類蘆主要富集Pb、Cd的部分,而類蘆對Pb2+具有較高的選擇性,其中可能是有一些通道對Pb有特定的吸附能力,還有待進一步的研究。
3.2類蘆對Pb2+、Cd2+的耐性機制
植物根系吸收離子主要是通過細胞外空間(質外體)或經由胞質(通過共質體),在本試驗中,細胞壁結合部分(質外體途徑)會干擾Pb2+、Cd2+流進細胞質的量,原則上,細胞壁吸附的重金屬不能被認為是真正的吸收,因此,利用洗脫液去除細胞壁松散的Pb2+、Cd2+后再作進一步分析[12]。細胞壁是重金屬離子進入植物體的第一道屏障,它與重金屬的結合作用是植物耐重金屬的原因之一,大量有害重金屬沉積在植物細胞壁,能阻止重金屬對細胞內溶物的傷害。由于重金屬離子被固定在細胞壁上,不能進入細胞質影響細胞內的代謝活動,使植物對重金屬表現出耐性[3,14]。有學者分別對礦山生態型的東南景天和少根紫萍進行研究,同樣發現重金屬鎘或鈾都主要分布在細胞壁上[15-16]。在本試驗中,洗脫后類蘆根細胞壁仍有較高的量,說明類蘆根細胞壁對Pb2+、Cd2+的吸附能力較強,同時,進入植物葉片和根部的Pb、Cd主要分布在細胞壁上且一直保持較高的比例,僅有少量分布在細胞核和線粒體等組分,可能是因為根細胞壁中固定了大量Pb、Cd,減少了重金屬的流通量,減弱了Pb、Cd向地上部分遷移的能力。這些說明了可能是由于凱氏帶的存在,離子在進入中柱之前,必須首先進入共質體,而凱氏帶則能有效減少重金屬離子進入的量[17]。
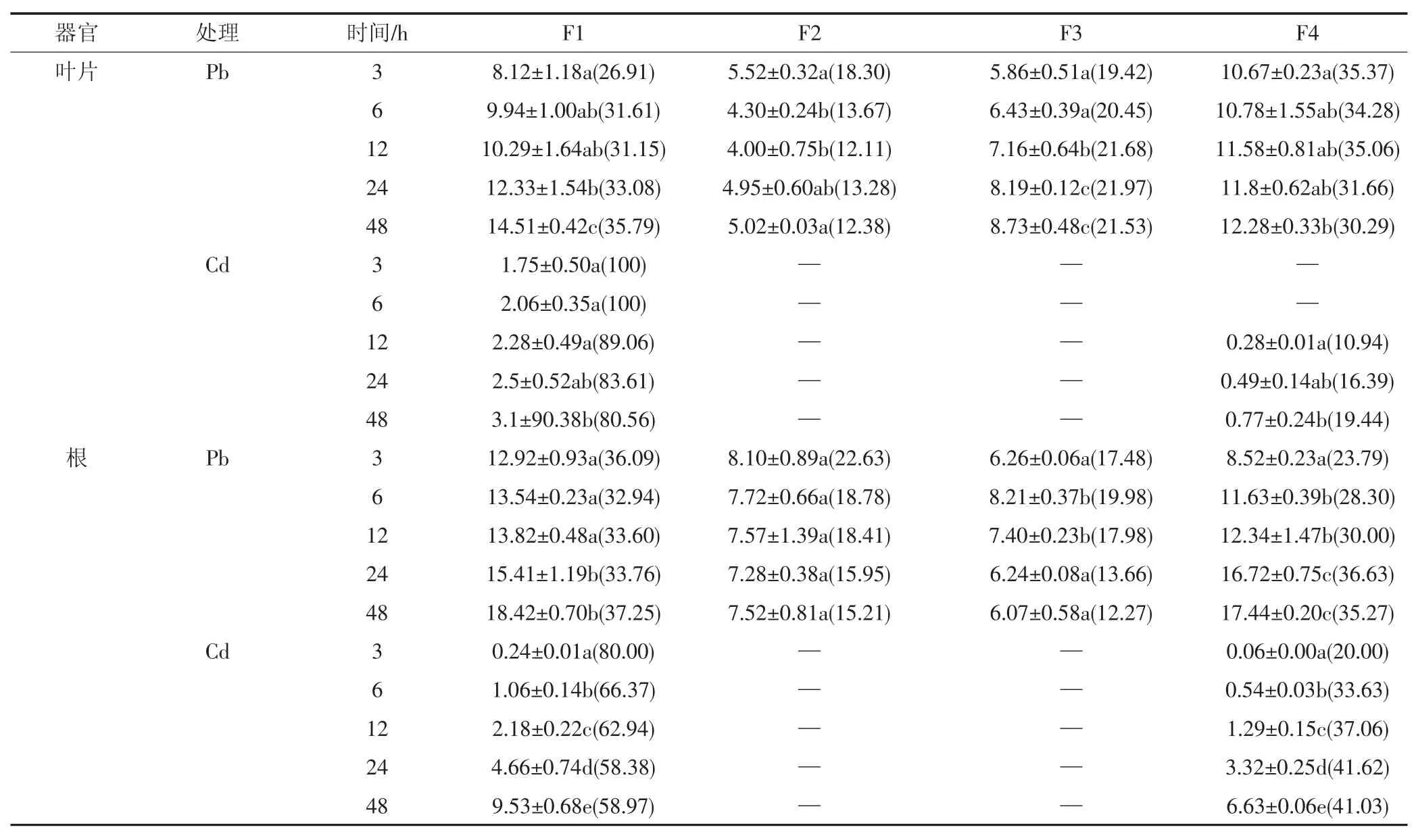
表4 不同時間處理下Pb2+、Cd2+在類蘆體內的亞細胞分布(mg·kg-1)Table 4 Subcellular distribution of Pb2+and Cd2+in Neyraudia reynaudiana under different stress times(mg·kg-1)
Pb、Cd進入細胞內會進一步分布到各種細胞器中,在本試驗中,F4組分在總量中占有相當的比重,F4組分為含核糖的可溶性組分,主要由液泡和細胞質兩部分組成。隨著Pb、Cd濃度的增加和處理時間的延長,細胞壁沉淀達到飽和及細胞質膜斷裂后,多余的Pb、Cd便進入原生質體內,進入原生質中的重金屬可通過向液泡輸送來降低其在原生質體中的濃度,使植物表現出耐性。液泡是植物細胞代謝副產品及廢物囤積場所,其含有的多種蛋白質、有機酸、有機堿等物質都能與重金屬結合,使重金屬離子在細胞內被區隔化,從而明顯降低了細胞質中游離重金屬離子的濃度[18]。因此,類蘆在受到重金屬脅迫時,部分Pb、Cd向可溶組分轉移,可能是類蘆通過改變Pb、Cd在亞細胞組分中的分布以增強植物對重金屬的耐受性。
類蘆具有的對Pb、Cd富集能力、耐受性及生物量大等特點,或能使其成為植物修復的潛能植物。而目前,在植物修復方面,超積累植物與非超積累植物最大的區別在于其向地上部轉運重金屬的能力,對生物量大而富集量一般的耐性植物而言,EDTA、DTPA、有機肥等螯合劑,尤其是胡敏酸等天然螯合劑得到了廣泛的使用,這些螯合劑能促使植物將大量的重金屬轉運至地上部分[19]。但是何種螯合劑能促進類蘆對重金屬的吸收,螯合劑在類蘆體內的螯合機制,以及是否對環境有潛在的風險等仍需進一步的討論和深入的研究。
4 結論
(1)類蘆對Pb、Cd吸收隨著濃度和時間的增加而增加,吸收的Pb、Cd主要分布在植物的根部,在相同濃度和時間處理下,類蘆體內的Pb明顯比Cd含量高,但是對Cd的吸收有一定的潛力。類蘆對Pb、Cd的吸收規律能夠利用Freundlich方程和Michaelis-Menten方程進行擬合。
(2)細胞壁是Pb、Cd在類蘆細胞內主要的分布位點,尤其是Pb在植物根中最高能達到90.43%,其次是可溶組分,細胞核、葉綠體組分和線粒體組分含量較低。重金屬在類蘆體內的動態分布使得類蘆對Pb、Cd有一定的耐性,維持了植物自身的穩態。
[1]Xu S S,Lin S Z,Lai Z X.Cadmium impairs iron homeostasis in Arabidopsisthaliana by increasing the polysaccharide contents and the ironbinding capacity of root cell walls[J].Plant and Soil,2015,392(1):71-85.
[2]Lamhamdi M,Galiou O E,Bakrim A,et al.Effect of lead stress on mineral content and growth of wheat(Triticum aestivum)and spinach (Spinacia oleracea)seedlings[J].Saudi Journal of Biological Sciences,2013,20(1):29-36.
[3]黃化剛,李廷軒,楊肖娥,等.植物對Pb脅迫的耐性及其解毒機制研究進展[J].應用生態學報,2009,20(3):696-704. HUANG Hua-gang,LI Ting-xuan,YANG Xiao-e,et al.Research advances in plant lead tolerance and detoxification mechanism[J].Journal of Applied Ecology,2009,20(3):696-704.
[4]荊紅梅,鄭海雷,趙中秋,等.植物對鎘脅迫響應的研究進展[J].生態學報,2001,21(12):2125-2130. JING Hong-mei,ZHENG Hai-lei,ZHAO Zhong-qiu,et al.Progresses of plants response to cadmium[J].Acta Ecologica Sinica,2001,21(12):2125-2130.
[5]王友生,侯曉龍,吳鵬飛,等.長汀稀土礦廢棄地土壤重金屬污染特征及其評價[J].安全與環境學報,2014,14(4):259-262. WANG You-sheng,HOU Xiao-long,WU Peng-fei,et al.Characteristics and evaluation of heavy metal pollution in the waste land of Changting rare earth mine[J].Journal of Safety and Environment,2014,14(4):259-262.
[6]戴文嬌,寧平,劉曉海,等.類蘆對鉛的耐性及富集能力探討[J].環境工程學報,2008,2(7):1004-1008. DAI Wen-jiao,NING Ping,LIU Xiao-hai,et al.Tolerance and accumulation of Neyraudia reynaudiana for Pb[J].Chinese Journal of Environmental Engineering,2008,2(7):1004-1008.
[7]戴文嬌.類蘆對鉛的耐性及吸收富集研究[D].昆明:昆明理工大學,2008. DAI Wen-jiao.Lead tolerance and accumulation in Neyraudia reynaudiana(Kunth)Keng[D].Kunming:Kunming University of Science and Technology,2008.
[8]蔡麗平,吳鵬飛,侯曉龍,等.類蘆根系對不同強度干旱脅迫的形態學響應[J].中國農學通報,2012,28(28):44-48. CAILi-ping,WUPeng-fei,HOUXiao-long,et al.Morphological response to different drought stress in the roots of Neyraudia reynaudiana[J].Chinese Agricultural Science Bulletin,2012,28(28):44-48.
[9]袁中友,郭彥彪,李強,等.有機無機肥配施對生態重建先鋒植物類蘆生長的影響[J].水土保持學報,2014,25(5):302-308. YUAN Zhong-you,GUO Yan-biao,LI Qiang,et al.Effects of organic-inorganic fertilizer application on pioneer plant Neyraudia reynaudiana on ecological restoration[J].Journal of Soil and Water Convervation,2014,25(5):302-308.
[10]周小勇,仇榮亮,胡鵬杰,等.鎘和鉛對長柔毛委陵菜體內鋅的亞細胞分布和化學形態的影響[J].環境科學,2008,29(7):2028-2036. ZHOUXiao-yong,QIURong-liang,HUPeng-jie,et al.Effects of cadmium and lead on subcellular distribution and chemical form of zinc in potentilla griffithii var.velutina[J].Environmental Science,2008,29(7):2028-2036.
[11]安增莉,侯艷偉,蔡超水,等.水稻秸稈生物炭對Pb(Ⅱ)的吸附特性[J].環境化學,2011,30(11):1851-1857. AN Zeng-li,HOU Yan-wei,CAI Chao-shui,et al.Adsorption characteristics of Pb(Ⅱ)from rice straw biological carbon[J].Environmental Chemistry,2011,30(11):1851-1857.
[12]陳彩虹,劉治昆,陳光才,等.蘇柳172和垂柳對Cu2+的吸收特性及有機酸影響[J].生態學報,2011,31(18):5255-5263. CHEN Cai-hong,LIU Zhi-kun,CHEN Guang-cai,et al.Uptake kinetic characteristics of Cu2+by Salix jiangsuensis CL J-172 and Salix babylonica Linn and the influence of organic acids[J].Acta Ecologica Sinica,2011,31(18):5255-5263.
[13]Zhao L,Li T,Yu H,et al.Changes in chemical forms,subcellular distribution,and thiol compounds involved in Pb accumulation and detoxification in Athyrium wardii(Hook.)[J].Environmental Science and Pollution Research,2015,22(16):12676-12688.
[14]Meyer C L,Juraniec M,Huguet S,et al.Intraspecific variability of cadmium tolerance and accumulation,and cadmium-induced cell wall modifications in the metal hyperaccumulator Arabidopsis halleri[J].Journal of Experimental Botany,2015,66(11):3215-3227.
[15]Nie X,Dong F,Liu N,et al.Subcellular distribution of uranium in the roots of Spirodela punctata and surface interactions[J].Applied Surface Science,2015,347(30):122-130.
[16]Ni T H,Wei Y Z.Subcellular distribution of cadmium in mining ecotype Sedum alfredii[J].Journal of Integrative Plant Biology,2003,45(8):925-928.
[17]泰茲.植物生理學[M].五版.北京:科學出版社,2009. Taiz L.Plant physiology[M].Fifth edition.Beijing:Science Press,2009.
[18]徐君,賈榮,施國新,等.鎘在水花生葉片中的亞細胞分布及其毒理學[J].應用生態學報,2012,23(4):1070-1076. XU Jun,JIA Rong,SHI Guo-xin,et al.Subcellular distribution and phytotoxicity of cadmium in Alternanthera philoxeroides leaves[J].Journal of Applied Ecology,2012,23(4):1070-1076.
[19]劉婷婷,彭程,王夢,等.海州香薷根細胞壁對銅的吸附固定機制研究[J].環境科學學報,2014,34(2):514-523. LIU Ting-ting,PENG Cheng,WANG Meng,et al.Mechanism of fixation and adsorption of copper on root cell wall of Elsholtzias plendens [J].Acta Scientiae Circumstantiae,2014,34(2):514-523.
Uptake kinetic characteristics and subcellular distribution of Pb2+and Cd2+in Neyraudia reynaudiana
LUO Jie-wen,HUANG Mei-ying,YIN Dan-yang,WU Peng-fei,CAI Li-ping,ZHOU Chui-fan*
(College of Forestry,Fujian Agriculture and Forestry University,Fuzhou 350002,China;Co-innovation Center for Soil and Water Conservation in Red Soil Region of the Cross-straits,Fuzhou 350002,China)
Neyraudia reynaudiana is a soil-conservating plant and can grow well in heavy metal polluted soils.Here a solution culture experiment was conducted to examine uptake and subcellular distrubution of Pb and Cd in the plant under different metal levels and stress times.Results showed that the uptake of Pb and Cd by N.reynaudiana increased with heavy metal concentrations and stress time.Freundlich and Michaelis-Menten equations fit the data very well,R2>0.95.It was also revealed that heavy metals were mainly distributed in the roots.Neyraudia reynaudiana had greater absorption ability for Pb(the highest content reached to 4 687.87 mg·kg-1),but higher absorption rate for Cd.For subcellular distribution,Pb and Cd were mainly distributed in the cell walls and soluble constituents.Under different concentrations and stress times of Pb and Cd,the Pb/Cd content in cell wall and soluble constituent subcellular components accounted for over 60%of the total.Therefore,N.reynaudiana has stronger absorption ability of Pb,and it could change the subcellular distribution of heavy metals to reduce the toxicity of heavy metals for maintaining its steady state.This study provides theoretical support for screening phytoremediation materials.
Neyraudia reynaudiana;Pb;Cd;absorption kinetics;subcellular distribution
X503.23
A
1672-2043(2016)08-1451-07
10.11654/jaes.2016-0202
2016-02-19
國家科技支撐計劃(2014BAD15B02);農業高校產學合作科技重大項目(2013N5002);福建省高校杰出青年科研人才培育計劃(2015);福建省自然科學基金項目(2013J01073)
羅潔文(1991—),女,碩士研究生,主要從事生態修復研究。E-mail:ellenlll@163.com
周垂帆E-mail:zhouchuifan@163.com
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
