溫度對不同葉齡長雄野生稻及其后代化感作用的影響
2016-10-25 03:00:14徐高峰申時才張付斗張玉華
中國水稻科學 2016年5期
關鍵詞:水稻
徐高峰 申時才 張付斗 張玉華
(云南省農業科學院 農業環境資源研究所, 昆明 650205; *通訊聯系人, E-mail: fdzh@vip.sina.com)
?
溫度對不同葉齡長雄野生稻及其后代化感作用的影響
徐高峰申時才張付斗*張玉華
(云南省農業科學院 農業環境資源研究所, 昆明 650205;*通訊聯系人, E-mail: fdzh@vip.sina.com)
XU Gaofeng, SHEN Shicai, ZHANG Fudou, et al. Allelopathic response to different temperature conditions of wild rice (Oryzalongistaminata) and its descendants. Chin J Rice Sci, 2016, 30(5): 559-566.
掌握溫度對長雄野生稻(Oryzalongistaminata)及其后代化感作用的影響, 對開發利用野生種質資源的化感抗草基因具有重要意義。采用直接樹脂吸收法收集不同葉齡長雄野生稻及其后代在各溫度條件下的根系分泌物,并以稗草作為受體,測定其化感作用。結果表明:1)溫度、葉齡和溫度×葉齡對水稻化感作用的影響與其自身化感潛力正相關,且對強化感和中化感潛力水稻(OL、F1、RL159 和RL169)的化感作用影響極顯著,而對弱化感水稻(RD23和RL219)的化感作用影響不顯著。2)供試條件下長雄野生稻及其后代根系分泌物對稗草的化感作用均為抑制作用,其中,低溫不利于水稻化感潛力的發揮。15℃時, 強和中化感潛力水稻在不同葉齡對稗草的苗高、根長和生物量的抑制作用最弱,化感綜合效應指數也顯著大于同等葉齡的高溫處理。3)高溫有利于水稻化感潛力的發揮,但并非總是隨溫度的升高而增強。在6葉期和8葉期,水稻材料OL、F1、RL159和RL169隨溫度的升高化感作用逐漸增強,但在2葉期和4葉期則表現為先增強后減弱,在25℃時對稗草化感作用最強。4)在15℃時,強和中等化感潛力水稻在2葉期化感作用最弱,而在其他溫度條件下,其化感作用隨葉齡的增加表現為先減弱后增強,在2葉期化感作用最強。研究結果顯示,溫度對不同葉齡的水稻化感作用具有調控作用,幼苗階段高溫有利于強和中化感潛力水稻化感作用的發揮。
長雄野生稻; 根系; 分泌物; 溫度; 葉齡; 化感作用
稻田雜草嚴重影響水稻的產量和品質,化學除草劑是現代農業控制稻田雜草的主要手段,但長期大量施用化學除草劑對生態環境以及食品安全的負面效應已越來越被關注。水稻化感作用是水稻通過釋放次生代謝物質影響臨近生物(如田間微生物和雜草等)生長發育的化學生態學現象[1-2]。利用水稻化感作用控制稻田雜草,將降低或取代現有農業生產對化學除草劑的依賴,保護環境和生物多樣性,是21世紀農業可持續發展關鍵技術之一[4-7]。然而,由于人類長期馴化培育,現代作物品種的化感特性大多已喪失或特征基因不再表達,即便是那些極少數保留化感特性的水稻品種, 其在產量、品質和農藝性狀等方面也已不能滿足目前生產種植的要求[8-10]。
作物野生類群中蘊涵著高產、抗逆等特異性狀基因,已成為現代水稻育種抗逆基因的一個重要來源[11]。利用野生稻化感抗雜草基因,培育高產、優質、抗雜草的化感水稻新品種具有重要的科學意義和應用價值[12-13]。長雄野生稻與亞洲栽培稻(O.sativa)具有相同的AA基因組,是向亞洲栽培稻轉移抗生物和非生物脅迫有利基因的重要基因庫[14]。張付斗等[15]和郭恰卿等[16]研究表明,長雄野生稻在2~3葉期對稗草的化感作用與國際公認的化感水稻PI312777差異不顯著,具有較強的化感潛力[15-16]。因此,研究利用長雄野生稻化感作用特性,改良現代栽培稻的抗草能力具有重要的科學意義和應用價值。
植物化感作用強度與其本身所產生特征物質的種類和濃度有關[17]。水稻化感是多基因控制的數量遺傳性狀,環境條件對水稻化感特征物質的產生、釋放、生物活性以及在環境中的行為產生廣泛而復雜的影響[13]。如胡飛等[18]研究顯示,水、肥、光等環境因子均會影響水稻化感作用的發揮。孔垂華等[17]研究表明化感水稻在不同環境條件下所產生與釋放的化感特征物質在種類與濃度方面存在差異。張付斗等[15]研究表明,不同水肥條件對長雄野生稻化感潛力的發揮具有顯著影響,其化感作用可被外界環境所調控。
溫度是影響植物生長、發育以及空間分布的主要生態因子,也對植物化感潛力的發揮具有重要影響。掌握溫度對不同葉齡長雄野生稻(Oryzalongistaminata)及其后代化感潛力的影響,對充分發揮水稻的化感潛力,促進野生種質資源化感抗草基因的開發利用具有重要的理論與實際意義。為此, 本研究以長雄野生稻、亞洲栽培稻“RD23”及其F1(RD23×O.longistaminata)和3份化感潛力不同后代作為試驗材料, 探索溫度對不同葉齡水稻材料化感作用的影響,以加深人們對溫度對化感作用的理解,為化感抗稗草水稻新品種的選育及其田間應用奠定基礎。
1 材料與方法
1.1供試材料
通過雜交方法將長雄野生稻與亞洲栽培稻RD23雜交所培育的F2群體內具有化感抗雜草特性,即攜帶主效QTL植株用RD23回交,每次回交用已定位標記進行分子標記輔助選擇,目標植株回交至BC2F1,自交至BC2F3,完成了近等基因系材料的培育。供試水稻材料長雄野生稻、亞洲栽培稻RD23 (泰國優質秈稻)及其F1(RD23 ×O.longistaminata)和3份具有不同化感潛力的近等基因系材料(分別為強化感潛力水稻材料RL159、中化感潛力水稻材料RL169和弱化感潛力水稻材料RL219),由云南省農業科學院糧食作物研究所提供。各試材在2~3葉期葉片水提液對稗草的化感指數以及在不同生育期的形態特征見文獻[19]。供試稗草(Echinochloacrusgalli) 種子經實驗室多年純化,發芽率>98%。
1.2試驗方法
1.2.1各葉齡水稻試材在不同溫度條件下的根系分泌物收集
參照孔垂華等直接樹脂吸收法[20],收集不同溫度條件下各葉齡供試材料的根系分泌物。具體如下:首先,挑選籽粒飽滿的供試水稻材料種子,將其催芽后分別播于秧盤中。在水稻的2、4、6和8葉齡期,分別挑選健壯均勻的供試秧苗移入盛有Yoshida營養液和AG 50W-X8樹脂的塑料盒(25 cm×20 cm×10 cm)中,每盒栽種3株,每處理栽種4盒。后分別置于溫度為15℃、20℃、25℃和30℃,每天光照14 h的人工氣候箱中恒溫培養10 d,期間每天補充一定量相同濃度的營養液。培養結束后從塑料盒中取出樹脂,仔細清除樹脂中的水稻根組織,然后將樹脂和溶液倒入5 cm×20 cm的玻璃柱中,待柱中的溶液流盡后,先用蒸餾水洗柱,再用甲醇淋洗,甲醇淋洗液低溫減壓除去溶劑,得根系分泌物,置于4℃的冰箱中密封無菌保存備用。
1.2.2根系分泌物化感作用的生物測定
試驗采用培養皿濾紙法,每處理重復4次。具體如下:在直徑9 cm的培養皿中墊定性濾紙,后每皿撒入經挑選的稗草種子50 粒。將前期收集的各處理根系分泌物分別用蒸餾水定容至50 mL,搖勻后每培養皿移入10 mL根系分泌物,空白對照加入10 mL無菌水。密封后在光照強度10 000 lx,溫度(28±0.5) ℃,每天光照12 h的人工氣候箱中恒溫培養,7 d后取出稗草并用吸水紙吸干植株上粘附的水分, 調查稗草的株高、根長和生物量。
1.3數據分析
供試材料根系分泌物對稗草的化感效應指數(RI)采用Williamson等[21]的方法: RI=1C/T(當T≥C時)或RI=T/C1(當T
所有數據采用DPS 9.01軟件進行統計分析。對測定指標進行二因素方差分析(固定模型)(Two-way ANOVA),分別計算溫度、葉齡以及二因子(溫度×葉齡)互作效應的F值,Duncan多重檢驗。并采用單因素方差分析(One-Way ANOVA)對不同溫度和葉齡條件下供試材料的化感綜合效應進行檢驗, Duncan新復極差法進行多重比較, 分析不同處理間的差異。
2 結果與分析
2.1水稻在不同葉齡和溫度條件下的根系分泌物對稗草生長的影響
研究結果顯示,各葉齡供試水稻材料在不同溫度條件下的根系分泌物對稗草的苗長、根長和生物量的影響均為抑制作用,同等條件下抑制強度為根長>生物量>苗長。在15℃時,相同葉齡供試材料的根系分泌物對稗草的苗長、根長和生物量的抑制作用最弱。而后隨著溫度的上升,在2葉期和4葉期,除RD23和RL219外,其他各處理根系分泌物對稗草生長的抑制作用表現為先增強后減弱,在25℃時抑制作用達到最強;而在6葉期和8葉期,各處理根系分泌物對稗草生長的抑制作用則表現為逐漸增強,在30℃時抑制作用最強。研究結果也表明,在15℃下,除RD23和RL219外。其他不同葉齡水稻根系分泌物對稗草苗長、根長和生物量的抑制作用隨著葉齡的增加逐漸增強,8葉期達到最大;而在其他溫度條件下(20℃、25℃和30℃),各處理根系分泌物對稗草苗長、根長和生物量的抑制作用,表現為先減弱后增強,在2葉期抑制作用最強,6葉期抑制作用最弱。另外,研究也顯示,強化感潛力水稻材料,如OL、F1和RL159,其根系分泌物對稗草苗長、根長和生物量的影響受溫度和葉齡影響明顯大于中化感潛力和弱化感潛力水稻材料;而弱化感潛力材料,如RD23和RL219,隨著溫度和葉齡的改變,其根系分泌物對稗草的抑制作用雖也有波動,但波動較弱(表1、表2和表3)。
2.2水稻在不同葉齡和溫度條件下的根系分泌物對稗草化感綜合效應指數分析
由表4可知,溫度對不同葉齡水稻材料根系分泌物的化感作用具有顯著影響。隨著溫度的上升,在2葉和4葉期,供試強化感和中等化感水稻材料(OL、F1、RL159 和RL169),在25℃時,其根系分泌物的化感綜合效應指數(RISE)最小,顯示在該條件下供試水稻的化感作用最強;而在6葉期和8葉期,其根系分泌物的化感綜合效益指數在30℃時RISE最小,表明供試水稻材料在該條件下化感作用最強。研究結果也表明試驗水稻材料在不同葉齡應對溫度變化的化感作用存在差異。在20℃、25℃和30℃時,水稻材料OL、F1、RL159和RL169對稗草的化感作用隨葉齡的增加表現為先減弱后增強,其中在2葉期對稗草的化感作用最強,6葉期時的化感作用最弱;而在15℃時,上述材料的根系分泌物在2葉期化感作用均表現最弱,顯示低溫不利于水稻化感潛力的發揮。另外研究也表明,上述材料除長雄野生稻在30℃其2葉期的根系分泌物化感綜合效益指數RISE與其8葉期差異不顯著外,其他水稻材料隨著葉齡的增加,其根系分泌物的化感綜合效益指數(RISE)均顯著增大。
2.3溫度、葉齡以及二者互作對試驗水稻材料化感作用的影響分析
方差分析表明,溫度、葉齡及其互作效應對強化感和中等化感潛力水稻材料OL、F1、RL159 和RL169化感作用的影響均達極顯著水平,而且單因素效應均大于溫度和葉齡的二因素互作效應。溫度對RD23根系分泌物化感作用的影響均達極顯著水平,但葉齡以及溫度與葉齡二者互作對其化感作用的影響均不顯著。溫度和葉齡對RL219根系分泌物化感作用的影響均達極顯著水平,但二者互作對其根系分泌物化感作用的影響則不顯著。溫度對水稻材料OL、F1、RD23和RL219化感作用的影響大于葉齡,而葉齡對RL159 和RL169化感作用的影響則大于溫度(表5)。
表1不同溫度和葉齡條件下的水稻材料根系分泌物對稗草苗長的影響
Table 1. Effects of the root exudates of Oryza longistaminata on seedling height of barnyard grass at different temperature and leaf ages.
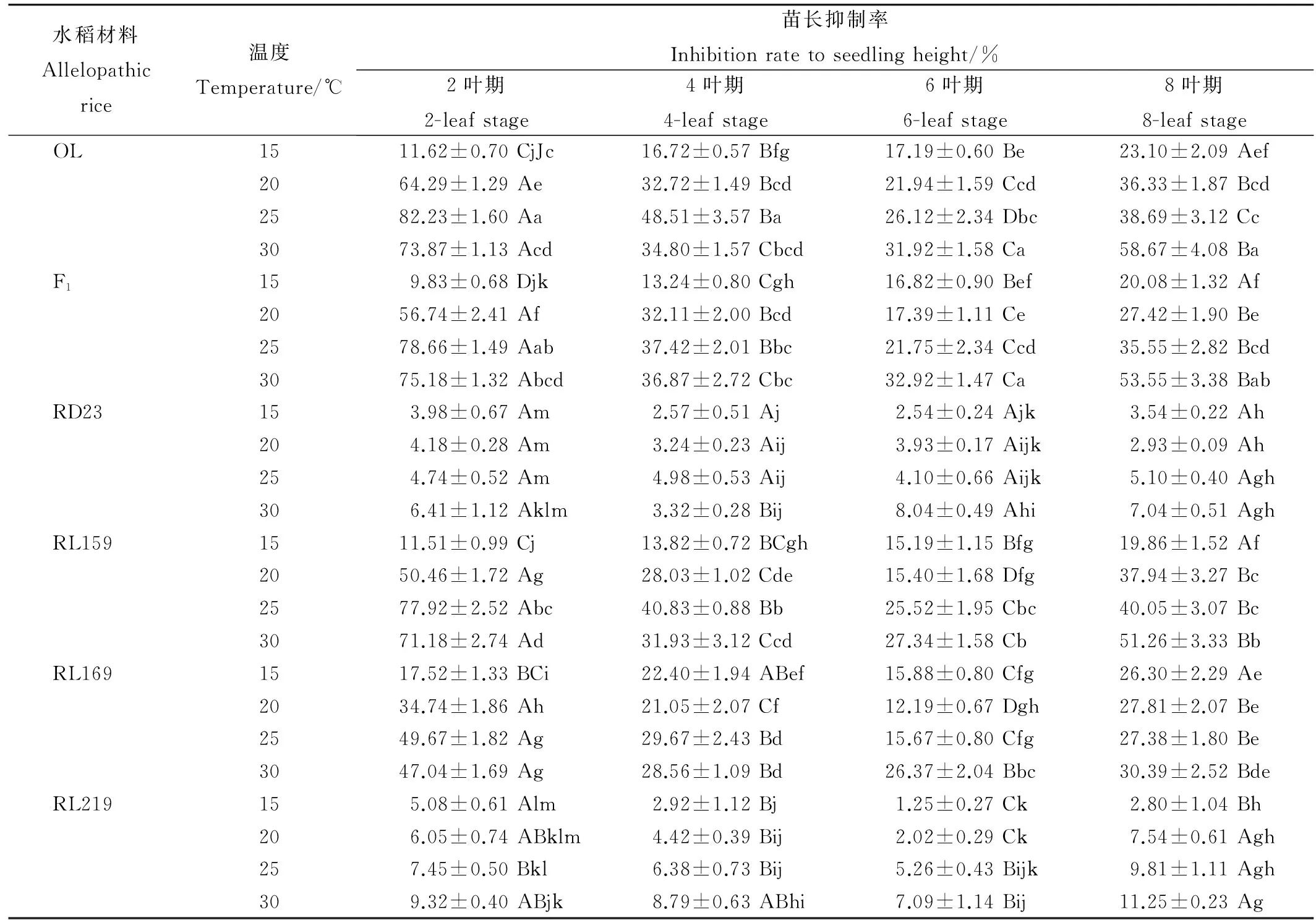
水稻材料Allelopathicrice溫度Temperature/℃苗長抑制率Inhibitionratetoseedlingheight/%2葉期2-leafstage4葉期4-leafstage6葉期6-leafstage8葉期8-leafstageOL1511.62±0.70CjJc16.72±0.57Bfg17.19±0.60Be23.10±2.09Aef2064.29±1.29Ae32.72±1.49Bcd21.94±1.59Ccd36.33±1.87Bcd2582.23±1.60Aa48.51±3.57Ba26.12±2.34Dbc38.69±3.12Cc3073.87±1.13Acd34.80±1.57Cbcd31.92±1.58Ca58.67±4.08BaF1159.83±0.68Djk13.24±0.80Cgh16.82±0.90Bef20.08±1.32Af2056.74±2.41Af32.11±2.00Bcd17.39±1.11Ce27.42±1.90Be2578.66±1.49Aab37.42±2.01Bbc21.75±2.34Ccd35.55±2.82Bcd3075.18±1.32Abcd36.87±2.72Cbc32.92±1.47Ca53.55±3.38BabRD23153.98±0.67Am2.57±0.51Aj2.54±0.24Ajk3.54±0.22Ah204.18±0.28Am3.24±0.23Aij3.93±0.17Aijk2.93±0.09Ah254.74±0.52Am4.98±0.53Aij4.10±0.66Aijk5.10±0.40Agh306.41±1.12Aklm3.32±0.28Bij8.04±0.49Ahi7.04±0.51AghRL1591511.51±0.99Cj13.82±0.72BCgh15.19±1.15Bfg19.86±1.52Af2050.46±1.72Ag28.03±1.02Cde15.40±1.68Dfg37.94±3.27Bc2577.92±2.52Abc40.83±0.88Bb25.52±1.95Cbc40.05±3.07Bc3071.18±2.74Ad31.93±3.12Ccd27.34±1.58Cb51.26±3.33BbRL1691517.52±1.33BCi22.40±1.94ABef15.88±0.80Cfg26.30±2.29Ae2034.74±1.86Ah21.05±2.07Cf12.19±0.67Dgh27.81±2.07Be2549.67±1.82Ag29.67±2.43Bd15.67±0.80Cfg27.38±1.80Be3047.04±1.69Ag28.56±1.09Bd26.37±2.04Bbc30.39±2.52BdeRL219155.08±0.61Alm2.92±1.12Bj1.25±0.27Ck2.80±1.04Bh206.05±0.74ABklm4.42±0.39Bij2.02±0.29Ck7.54±0.61Agh257.45±0.50Bkl6.38±0.73Bij5.26±0.43Bijk9.81±1.11Agh309.32±0.40ABjk8.79±0.63ABhi7.09±1.14Bij11.25±0.23Ag
表中數據為平均值±標準誤。不同大寫字母表示同一行中數據在5%水平上差異顯著;不同小寫字母表示同一列數據在5%水平上差異顯著。表2、表3和表4同。
The values in the table were average±standard error. Different capital letters in the same column mean significant different at 5% level, and different small letters in the same line mean significant different at 5% level according to Duncan’s test. The same as in Table 2, 3 and 4.
3 討論
植物化感作用是復雜的化學生態學現象,受遺傳、環境、苗齡等多種因素共同影響,具有較強化感潛力的植物田間并不都能表現出抑草效應[10,23]。環境條件對水稻化感作用的影響, 是選育與栽培化感抗雜草水稻品種的重要基礎[24]。不同作物品種或品系產生化感作用因環境條件的不同,其作用強度也有所差別。本研究結果表明,溫度、葉齡及其二因子(溫度×苗齡)互作對水稻化感作用的影響復雜,具有強化感或中等化感潛力的水稻材料OL、F1、RL159 和RL169,其化感作用受溫度、葉齡及其二因子(溫度×苗齡)互作的影響顯著大于弱化感材料(RD23);且單因素效應均大于二因素互作效應;而弱化感材料RD23的化感綜合效應指數(RISE)則隨溫度和葉齡的改變無顯著變化。如研究顯示,葉齡為2葉期的長雄野生稻,在25℃時其綜合化感效應指數為-0.91,具有較強的化感抑草效應;而在同等葉齡條件下,在15℃時其綜合化感效應指數僅為-0.14,化感作用顯著降低。長雄野生稻是本項目組前期篩選得到的對稗草具有較強抑制作用的化感水稻材料[19]。因此,本研究不僅對充分發揮化感水稻的化感潛力提供指導,也進一步驗證了為何強化感潛力的水稻品種田間并不都表現出化感效應。
表2不同溫度和葉齡條件下的水稻材料根系分泌物對稗草根長的影響
Table 2. Effects of the root exudates of Oryza longistaminata on root length of barnyard grass at different temperature and leaf ages.

水稻材料Allelopathicrice溫度Temperature/℃根長抑制率Inhibitionratetorootlength/%2葉期2-leafstage4葉期4-leafstage6葉期6-leafstage8葉期8-leafstageOL1514.92±0.80Cij23.30±1.32Bh27.86±1.53Bf32.98±2.08Ae2067.79±2.27Ad54.71±2.41Cde32.87±2.00Dde60.92±0.88Bc2595.57±0.60Aa61.87±2.90Dabc43.29±1.44Cab80.64±2.45Bb3091.29±1.43Aab57.14±1.93Ccde45.94±1.51Da88.84±0.71BaF11512.82±0.84Bj15.71±0.56Bij21.28±1.67Ag25.18±2.01Af2064.29±1.89Ad59.90±2.92Bbcd27.03±1.01Cf59.27±2.24Bc2592.07±0.56Aab66.27±2.11Ba41.55±1.40Cb61.50±3.44Bc3089.64±2.16Ab64.97±2.74Ca43.14±2.46Dab83.94±2.20BbRD23156.11±0.50Ak3.22±0.16Cl4.03±0.39BCkl5.03±0.77ABi204.64±1.05Ak5.40±0.45Akl5.42±0.66Akl4.42±0.21Ai254.07±1.16Bk7.75±0.31Akl5.85±0.58ABkl6.85±0.80Ai303.68±0.73Bk6.91±0.68ABkl8.94±0.38Ajk7.94±0.71AiRL1591527.27±1.70Cg32.76±1.42Bg36.01±2.12Bcd47.06±1.67Ad2057.37±2.74Ae31.87±2.91Cg17.51±1.16Dgh43.14±1.33Bd2588.60±2.32Ab46.42±3.33Bf29.02±1.83Cef45.54±1.42Bd3080.93±0.72Ac36.31±2.11Cg31.08±1.80Def58.28±1.66BcRL1691541.53±1.42Cf53.08±0.69Be37.62±1.01Cc62.33±0.87Ac2039.50±1.46Af23.93±2.27Ch13.86±0.63Dhi31.62±1.59Be2556.48±2.52Ae33.73±1.20Bg17.82±0.71Cgh31.13±0.85Be3053.48±1.98Ae32.47±1.85Bg29.98±1.83Bef34.55±2.50BeRL2191512.04±1.64Ag5.23±0.58BCkl3.21±0.41Cl6.98±0.80Bi2014.35±2.01ABij10.69±0.98BCjk7.51±0.79Cjkl15.36±1.13Ah2517.66±3.36Ahi14.24±0.92ABij11.32±0.82Bij17.21±1.11ABgh3022.09±1.45Ah17.42±0.90Bi16.81±0.72Bh20.97±1.04Afg
表3不同溫度和葉齡條件下的水稻材料根系分泌物對稗草生物量的影響
Table 3. Effects of the root exudates of Oryza longistaminata on biomass of barnyard grass at different temperature and leaf ages.

水稻材料Allelopathicrice溫度Temperature/℃生物量抑制率Inhibitionratetobiomass/%2葉期2-leafstage4葉期4-leafstage6葉期6-leafstage8葉期8-leafstageOL1514.20±0.79Dkl19.81±0.88Ch23.14±1.01Bf30.00±1.14Ah2072.64±2.52Ad45.51±1.36Cd32.88±1.23Dcd53.49±1.34Bd2594.24±1.58Aa59.33±1.71Ca39.49±1.44Db66.26±1.59Bc3090.01±1.12Aab49.19±1.38Bc43.09±1.64Ca88.50±2.22AaF11512.12±0.61Clm14.33±0.81Ci19.57±0.72Bg24.21±0.88Ai2066.57±1.23Ae47.89±1.39Bcd26.65±1.52Ce47.68±1.60Be2590.49±3.15Aab55.73±2.58Bb35.11±2.00Cc53.88±1.76Bd3089.83±2.43Aab54.48±1.41Bb42.10±2.14Cab82.50±2.29AbRD23155.40±0.26An2.86±0.14Cm3.38±0.07Cl4.59±0.24Bm204.85±0.21An4.50±0.31Aklm5.61±0.14Akl4.04±0.19Am254.67±0.37An6.84±0.23Akl7.30±0.25Ajk6.64±0.31Alm305.50±0.21An5.48±0.22Aklm9.40±0.37Aj8.99±0.44AlRL1591520.74±0.37Bj23.06±0.58Bg26.29±1.13Be35.80±0.98Afg2059.31±1.05Af31.18±0.85Cf19.74±0.78Dg44.59±1.06Be2588.26±2.96Ab46.90±1.23Bcd29.58±0.99Cde47.52±1.37Be3082.90±2.56Ac36.51±0.83Ce32.33±1.12Dcd65.72±1.33BcRL1691531.59±1.06Ci37.36±1.06Be27.47±1.32De47.41±1.46Ae2040.83±0.92Ah23.41±0.96Cg15.63±1.08Dhi32.69±0.86Bgh2556.26±1.14Afg34.08±1.27Bef18.58±1.34Cgh32.48±1.46Bgh3054.79±0.74Ag32.65±0.94Cf31.19±1.08Cd38.97±1.26BfRL219159.16±0.60Amn4.03±0.29Clm2.29±0.14Cl5.23±0.25Bm2011.22±0.42Alm7.87±0.33Bjk5.72±0.21Bkl12.60±0.37Ak2513.31±0.48Akl11.08±0.39ABij9.20±0.44Bj15.00±0.33Ak3017.12±0.64ABjk14.02±0.69Ci13.23±0.82BCi19.33±0.65Aj
表4溫度對不同葉齡長雄野生稻及其后代化感綜合效應指數的影響
Table 4. Effects of leaf stage and temperature on allelopathy synthetic inhibitory effect of Oryza longistaminata and its descendants.
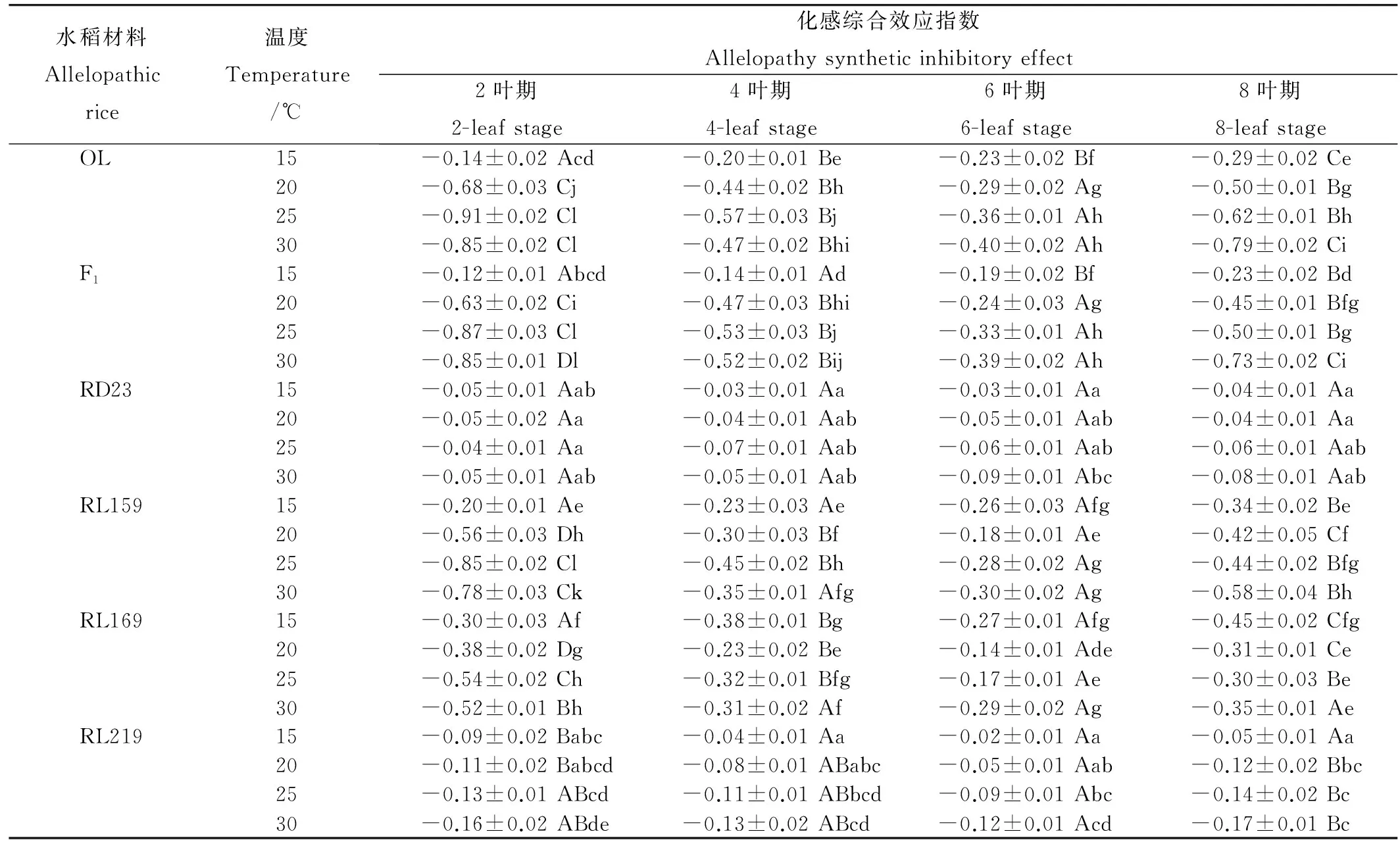
水稻材料Allelopathicrice溫度Temperature/℃化感綜合效應指數Allelopathysyntheticinhibitoryeffect2葉期2-leafstage4葉期4-leafstage6葉期6-leafstage8葉期8-leafstageOL15-0.14±0.02Acd-0.20±0.01Be-0.23±0.02Bf-0.29±0.02Ce20-0.68±0.03Cj-0.44±0.02Bh-0.29±0.02Ag-0.50±0.01Bg25-0.91±0.02Cl-0.57±0.03Bj-0.36±0.01Ah-0.62±0.01Bh30-0.85±0.02Cl-0.47±0.02Bhi-0.40±0.02Ah-0.79±0.02CiF115-0.12±0.01Abcd-0.14±0.01Ad-0.19±0.02Bf-0.23±0.02Bd20-0.63±0.02Ci-0.47±0.03Bhi-0.24±0.03Ag-0.45±0.01Bfg25-0.87±0.03Cl-0.53±0.03Bj-0.33±0.01Ah-0.50±0.01Bg30-0.85±0.01Dl-0.52±0.02Bij-0.39±0.02Ah-0.73±0.02CiRD2315-0.05±0.01Aab-0.03±0.01Aa-0.03±0.01Aa-0.04±0.01Aa20-0.05±0.02Aa-0.04±0.01Aab-0.05±0.01Aab-0.04±0.01Aa25-0.04±0.01Aa-0.07±0.01Aab-0.06±0.01Aab-0.06±0.01Aab30-0.05±0.01Aab-0.05±0.01Aab-0.09±0.01Abc-0.08±0.01AabRL15915-0.20±0.01Ae-0.23±0.03Ae-0.26±0.03Afg-0.34±0.02Be20-0.56±0.03Dh-0.30±0.03Bf-0.18±0.01Ae-0.42±0.05Cf25-0.85±0.02Cl-0.45±0.02Bh-0.28±0.02Ag-0.44±0.02Bfg30-0.78±0.03Ck-0.35±0.01Afg-0.30±0.02Ag-0.58±0.04BhRL16915-0.30±0.03Af-0.38±0.01Bg-0.27±0.01Afg-0.45±0.02Cfg20-0.38±0.02Dg-0.23±0.02Be-0.14±0.01Ade-0.31±0.01Ce25-0.54±0.02Ch-0.32±0.01Bfg-0.17±0.01Ae-0.30±0.03Be30-0.52±0.01Bh-0.31±0.02Af-0.29±0.02Ag-0.35±0.01AeRL21915-0.09±0.02Babc-0.04±0.01Aa-0.02±0.01Aa-0.05±0.01Aa20-0.11±0.02Babcd-0.08±0.01ABabc-0.05±0.01Aab-0.12±0.02Bbc25-0.13±0.01ABcd-0.11±0.01ABbcd-0.09±0.01Abc-0.14±0.02Bc30-0.16±0.02ABde-0.13±0.02ABcd-0.12±0.01Acd-0.17±0.01Bc
表5溫度對不同葉齡長雄野生稻及其后代化感作用影響的方差分析
Table 5. Variance analysis of interaction effects of leaf stage and temperature on allelopathy of Oryza longistaminata and their descendants.

水稻材料Allelopathicrice處理Treatment自由度df平方和SS均方MSF值FvalueP值PvalueOL溫度Temperature31.780.59371.870.0001葉齡Leafstage30.960.32201.080.0001溫度×葉齡Temperature×Leafstage90.600.0741.950.0001誤差Error480.080.00F1溫度Temperature31.880.63423.180.0001葉齡Leafstage30.730.24163.460.0001溫度×葉齡Temperature×Leafstage90.580.0643.490.0001誤差Error480.070.00RD23溫度Temperature30.010.007.910.0002葉齡Leafstage30.000.001.070.3710溫度×葉齡Temperature×Leafstage90.010.001.870.0797誤差Error480.020.00RL159溫度Temperature30.700.2388.390.0001葉齡Leafstage31.070.36135.930.0001溫度×葉齡Temperature×Leafstage90.600.0725.480.0001誤差Error480.130.00RL169溫度Temperature30.100.0321.500.0001葉齡Leafstage30.400.1384.700.0001溫度×葉齡Temperature×Leafstage90.230.0316.180.0001誤差Error480.080.00RL219溫度Temperature30.080.0341.500.0001葉齡Leafstage30.030.0115.010.0001溫度×葉齡Temperature×Leafstage90.000.000.710.699誤差Error480.030.00
溫度是植物生長必需的生態因子,也是水稻產量和品質的限制因子。另外,溫度也影響植物化感潛力的發揮,研究表明高溫通常能夠促進化感植物分泌酚酸,有利于其發揮化感潛力[25]。因此,研究溫度對水稻化感作用的影響,對選育與栽培化感抗雜草水稻品種,充分發揮化感水稻的化感潛力具有重要意義。本研究顯示,溫度對化感水稻化感潛力的發揮具有復雜的影響;不同苗齡、不同化感潛力的水稻其化感作用對溫度的響應存在明顯差異。有研究表明,低溫(15℃)不利于化感水稻材料化感潛力的發揮,隨著溫度的升高,強和中等化感潛力水稻的化感作用增強,但不同苗齡水稻化感潛力的變化存在差異。如在2葉期和4葉期,其化感作用隨溫度的升高表現為先增強后減弱,在25℃時對稗草化感作用最強,而在6葉期和8葉期,上述材料對稗草的化感作用均隨溫度的升高而增強。植物化感作用是植物應對環境壓力的一種適應性機制。Metlen等[26]認為,植物可能隨生長環境的變化而改變對化感物質的資源投入。水稻在不同生長階段對溫度的適應性存在差異。如2葉期和4葉期為水稻的幼苗期,其對溫度的耐受性較弱,低溫和高溫均不利于其生長,因此,其化感作用隨溫度的升高并非一直增強,而呈現出先增加后降低的趨勢。而在6葉期和8葉期,其對低溫和高溫的耐受性增強,化感作用也隨溫度的升高而一直增強。這或許是不同葉齡水稻其化感作用隨溫度波動而改變的原因之一。
水稻的化感作用受到遺傳和基因型與環境互作效應的共同影響[27]。在不同的環境條件下,水稻不同葉齡期化感作用的雜種優勢可分為由遺傳主效應決定的遺傳優勢和由基因型與環境互作引起的互作優勢[28]。在與雜草共生環境中, 雜交稻表現出比常規稻更強的資源競爭優勢,因此,雜種優勢利用是充分發揮水稻化感作用潛力的一條重要途徑。本研究結果表明,在20℃、25℃和30℃時,不同苗齡的OL、F1和F2代(RL159 和RL169)對稗草化感作用的強度為OL>F1>RL159 >RL169,且隨溫度變化的波動較為一致,表明在上述條件下供試材料對稗草的化感作用以遺傳優勢為主。而在15℃時,其對稗草化感作用的強度為RL169>RL159>OL和F1,顯示在低溫條件下,強和中等化感潛力的F1和F2代對稗草的化感作用表現為基因型與環境互作為主。研究表明在不同環境條件下互作優勢對水稻化感作用的影響較大,由環境效應引起的互作優勢對水稻化感作用的影響在不同環境下有著明顯的差異,也解釋了植物在不利環境中產生化感作用物質的數量有所增加的原因。
將不同化感特性的水稻品種雜交,可培育出既抗草又增產的雜交水稻品種。本研究表明,在溫度大于20℃時,長雄野生稻及其具有較強化感潛力的后代均表現出強化感作用,且在不同葉齡其化感潛力的發揮受溫度影響顯著。現有研究對利用長雄野生稻化感特性,培育高產抗稗草水稻材料具有指導作用,然而,上述研究在人工控制的單因子條件下進行,而在自然界復雜條件下其材料的遺傳穩定性及其化感作用對溫度響應的穩定性,還有待田間試驗進一步確證。
[1]Khanh T D, Xuan T D, Chung M I. Rice allelopathy and the possibility for weed management.AnnApplBiol, 2007, 151:325-339.
[2]Kong C H, Hu F, Wang P, et al. Effect of allelopathic rice varieties combined with cultural management options on paddy field weeds.PestManagSci, 2008, 64(3): 276-282.
[3]Duke S O. Allelopathy: Current status of research and future of the discipline: A commentary.AlleloJ, 2010, 25(1): 17-30.
[4]Kim K U. Trends and expectations for research and technology in the Asia-Pacific region.WeedBiolManag, 2001, 1(1): 20-24.
[5]Rice E L. Biological control of weeds and plant diseases: Advances in applied allelopathy. Norman, Oklahoma: Univ Oklahoma Press, 1995, 448.
[6]Pērez de Vida F B, Laca E A, Mackill D J, et al. Relating rice traits to weed competitiveness and yield: A path analysis.WeedSci, 2006, 54(6): 1122-1131.
[7]Kong C H, Zhao H, Xu X H, et al. Activity and allelopathy of soil of flavone O-Glycosides from rice.JAgricFoodChem, 2007, 55(15): 6007-6012.
[8]Putnam A R, Duke W B. Biological suppression of weeds: Evidence for allelopathy in accessions of cucumber.Science, 1974, 185(4148): 370-372.
[9]朱紅蓮, 孔垂華, 胡飛, 等. 水稻種質資源的化感潛力評價方法. 中國農業科學, 2003, 36(7): 788-792.
Zhu H L, Kong C H, Hu F, et al. Evaluation methods for allelopathic potentials of rice germplasms.SciAgricSin, 2003, 36(7): 788-792. (in Chinese with English abstract)
[10]孔垂華. 植物與其它有機體的化學作用-潛在的有害生物控制途徑. 中國農業科學, 2007, 40(4): 712-720.
Kong C H. Chemical interactions between plant and other organisms: A potential strategy for pest management.SciAgricSin, 2007, 40(4): 712-720. (in Chinese with English abstract)
[11]張付斗, 郭怡卿, 余柳青, 等. 野生稻和非洲栽培稻抗稗草種質資源篩選和評價. 作物學報, 2004, 30(11): 1140-1144.
Zhang F D, Guo Y Q, Yu L Q, et al. Evaluation and screening of resistance to barnyardgrass in germplasm of wild rice(Oryzasativa) and African cultivar.ActaAgronSin, 2004, 30(11): 1144-1148. (in Chinese with English abstract)
[12]Guo Y Q, Zhang F D, Tao D, et al. Preliminary studies on the allelopathic potential of wild rice (Oryza) germplasm.AllelJ, 2005, 15(1): 13-20.
[13]余柳青, 徐正浩, 郭怡卿, 等. 野生稻和非洲栽培稻抗稗草作用研究初報. 中國水稻科學, 2002, 16(3): 288-290.
Yu L Q, Xu Z H, Guo Y Q, et al. Resistance function on barnyardgrass in wild rice and African rice cultivars.ChinJRiceSci, 2002, 16(3):288-290. (in Chinese with English abstract)
[14]Soriano I R , Schmit V, Brar D S, et al. Resistance to rice roo-knot nematodeMeloidogynegraminicolaidentified inOryzalongistaminataandO.glaberrima.Nematology, 1999, 1(4): 395-398.
[15]張付斗, 徐高峰, 李天林, 等. 水氮互作下長雄野生稻化感作用與田間抑草作用. 作物學報, 2011, 37(1): 170-176.
Zhang F D, Xu G F, Li T L, et al. Allelopathy and weed-suppression ofOryzalongistaminataunder water-nitrogen interactions in the field.ActaAgronSin, 2011,37(2):170-176. (in Chinese with English abstract)
[16]郭怡卿, 張付斗, 陶大云, 等. 野生稻化感抗(耐)稗草種質資源的初步研究. 西南農業學報, 2004, 17(3): 295-298.
Guo Y Q, Zhang F D, Tao D Y,et al. A preliminary study on the allelopathic activity of wild rice germplasm.SouthWestChinaJAgricSci, 2004, 17(3): 295-298. (in Chinese with English abstract)
[17]孔垂華, 徐效華, 胡飛, 等. 以特征次生物質為標記評價水稻品種及單植株的化感潛力. 科學通報, 2002, 47(3): 203-206.
Kong C H, Xu X H, Hu F, et al. Evaluation for allelopathic potentials of rice germplasms and single plant species based on the characteristics of secondary metabolites.ChinSciBull, 2002, 47(3): 203-206.
[18]胡飛, 孔垂華, 陳雄輝, 等. 不同水肥和光照條件對水稻化感特性的影響. 應用生態學報, 2003, 14(12): 2265-2268.
Hu F, Kong C H, Chen X H, et al. Effects of different water, fertility, and light conditions on allelopathic traits of rice.ChinJApplEcol, 2003, 14(12):2265-2268. (in Chinese with English abstract)
[19]徐高峰, 申時才, 張付斗, 等. 長雄野生稻及其后代抑草效果與化感潛力和農藝性狀的關系. 中國生態農業學報, 2014, 22(11): 1348-1356.
Xu G F, Shen S C, Zhang F D, et al. Relationships among the weed suppression effect, allelopathy and agronomic characters ofOryzalongistaminataand their descendants.ChinJEco-Agric, 2014, 22(11): 1348-1356. (in Chinese with English abstract)
[20]孔垂華,徐效華,梁文舉,等. 水稻化感品種根分泌物中非酚酸類化感物質的鑒定與抑草活性. 生態學報, 2004, 24(7): 1317-1322.
Kong C H, Xu X H, Liang W J, et al. Non-phenolic allelochemicals in root exudates of an allelopathic rice variety and their identification and weed-suppressive activity.ActaEcolSin, 2004, 24(7): 1317-1322.(in Chinese with English abstract)
[21]Williamson G B, Richardson D. Bioassays for allelopathy: Measuring treatment responses within dependent controls.JChemEcol, 1988, 14(1): 181-187.
[22]廖周瑜, 趙則海, 侯玉平, 等. 五爪金龍對薇甘菊的化感效應研究. 生態環境, 2007, 16(3): 939-943.
Liao Z Y, Zhao Z H, Hou Y P, et al.Allelopathic effects ofIpomoeacairicaonMikaniamicrantha.EcolEnviron, 2007, 16(3): 939-943. (in Chinese with English abstract)
[23]Lin W X, Fang C X, Chen T, et al. Rice allelopathy and its properties of molecular ecology.FrontBiol, 2010, 5: 255-262.
[24]Wang P, Kong C H, Hu F, et al. Allantoin involved in species interactions with rice and other organism in paddy soil.PlantSoil, 2007, 296: 43-51.
[25] Natividad C L, Juan A G, Teresa S D, et al. Allelopathic potential ofCistusladaniferchemicals in response to variations of light and temperature.Chemoecology, 2002, 12:139-145.
[26]Metlen K L, Aschehoug E T, Callaway R M. Plant behavioural ecology: Dynamic plasticity in secondary metabolites.PlantCellEnviron, 2009, 32(9): 641- 653.
[27]何華勤, 林文雄, 董章杭, 等. 水稻對受體植物化感作用的遺傳生態學研究. 應用生態學報, 2002, 13(12) : 871-875.
He H Q, Lin W X, Dong Z H,et al. Genetic ecology of rice allelopathy on receiver plant.ChinJApplEcol, 2002, 13(12): 871-875. ( in Chinese with English abstract)
[28]林文雄, 董章杭, 何華勤,等. 不同環境下水稻化感作用的動態雜種優勢分析. 中國農業科學, 2003, 36(9): 985-990.
Lin W X, Dong Z H, He H Q, et al. Analysis on dynamic heterosis of rice allelopathy under different environmental conditions.SciAgricSin, 2003, 36(9): 985-990. ( in Chinese with English abstract)
Allelopathic Response to Different Temperature Conditions of Wild Rice (Oryza longistaminata) and Its Descendants
XU Gao-feng, SHEN Shi-cai, ZHANG Fu-dou*, ZHANG Yu-hua
(Agricultural Environment & Resource Research Institute, Yunnan Academy of Agricultural Sciences, Kunming 650205, China;*Corresponding author, E-mail: fdzh@vip.sina.com)
Rice allelopathy is considered as a complex phenomenon of chemical ecology, and temperature plays an important role in regulating allelopathic potential. Former studies have demonstrated that the wild rice (Oryzalongistaminata) has high allelopathic potential.Therefore,understanding the allelopathic responses to temperature of wild rice and its descendants is very important to finally exploit and utilize allelopathic genes of wild germplasm resources.The root exudates of different allelopathic potential rice accessions(OL, RD23, F1, RL159, RL169 and RL219)were respectively collected by the resin of AG 50W-X8,followed by the evaluation of their allelopathic potential to barnyardgrass.The results demonstrated: 1) Allelopathic response to temperature,leaf ages and temperature×leaf age were significantly different among different allelopathic potential rice accessions. The allelopathy was significantly affected by temperature, leaf age and temperature×leaf age of OL, F1, RL159 and RL169, but low allelopathic rice accessions of RD23 and RL219 followed an opposite trend. 2) Regardless of leaf ages,the root exudates have weakest inhibitory ability to the growth of barnyardgrass at low temperature(15℃). The effects of allelopathic rice accessions OL, F1, RL159 and RL169 were greater than low allelopathic ones at low temperature(15℃) condition. 3) High temperature was conducive to the allelopathic enhancement of allelopathic rice, but the allelopathy was not always enhanced with increasing temperature. The allelopathy was strengthened with increasing temperature of allelopathic rice accessions OL, F1, RL159 and RL169 when they were at the 6-leaf stage and 8-leaf stage, but at the 2-leaf stage and 4-leaf stage, the allelopathy was enhanced first and then decreased with increasing temperature, and the allelopathy was strongest at 25℃. 4)Allelopathic responses to leaf ages were significantly different among different allelopathic potential rice. The allelopathy is the highest at the 2-leaf stage for allelopathic rice accessions OL, F1, RL159 and RL169 except for 15℃, then the allelopathy weakened with the increasing rice leaf ages.This finding suggests that allelopathic enhancement by temperature conditions at different leaf ages were worthwhile in future for the understanding of allelopathic responses to associated weeds.
wild rice (Oryzalongistaminata); root exudates; temperature; leaf age; allelopathy
2015-10-30; 修改稿收到日期: 2016-03-22。
國家自然科學基金資助項目(31260453); 云南科技創新人才計劃資助項目(2014HB039)。
Q946.8; S451.1
A
1001-7216(2016)05-0559-08
徐高峰, 申時才, 張付斗, 等. 溫度對不同葉齡長雄野生稻及其后代化感作用的影響. 中國水稻科學, 2016, 30(5): 559-566.
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
