鵝掌楸DHHC型鋅指蛋白家族基因的克隆及表達分析
2016-10-28 01:39:43徐嘉娟李火根
廣西植物 2016年9期
徐嘉娟, 李火根
( 南京林業大學 林木遺傳與生物技術省部共建教育部重點實驗室, 南京 210037 )
?
鵝掌楸DHHC型鋅指蛋白家族基因的克隆及表達分析
徐嘉娟, 李火根
( 南京林業大學 林木遺傳與生物技術省部共建教育部重點實驗室, 南京 210037 )
棕櫚酰化修飾是一種最普遍且唯一可逆的翻譯后脂質修飾方式,賦予蛋白質多樣化的生理功能。DHHC(Asp-His-His-Cys)蛋白家族是一類與棕櫚酰化修飾相關的蛋白,多數DHHC蛋白家族成員具有蛋白質酰基轉移酶(protein S-acyltransferase,PAT)活性。該研究以鵝掌楸葉芽為材料,采用RT-PCR和RACE技術,克隆獲得了3個鵝掌楸DHHC蛋白家族基因cDNA全長,命名為LcPAT7、LcPAT22、LcPAT23。序列分析結果表明:LcPAT7、LcPAT22、LcPAT23基因全長分別為1 933、2 592、2 217 bp,各包含1 332、1 839、1 662 bp的開放閱讀框(Open Reading Frame,ORF),編碼433、612、533個氨基酸,預測蛋白分子量分別為40.04、67.3、60.57 kDa,理論等電點為9.15、9.03、7.29。3個基因編碼的蛋白均有4個跨膜區,并且都在跨膜域(transmembrane domain,TM)TM2和TM3之間存在一個DHHC蛋白家族典型的DHHC-CRD結構域。同源性分析表明:鵝掌楸LcPAT7、LcPAT22、LcPAT23編碼的氨基酸序列與其他植物中預測的PAT具有較高的相似性。利用熒光定量PCR技術檢測3個基因在鵝掌楸不同組織中的表達特性,發現3個基因在不同組織中均有表達,但表達量具有明顯區別。同一家族基因表達模式的變化表明其功能非冗余。該研究結果將為鵝掌楸生長發育與形態建成,以及逆境響應信號傳導等相關基因的調控研究提供了參考。
鵝掌楸, 棕櫚酰化修飾, 蛋白質酰基轉移酶, DHHC蛋白家族, 表達分析
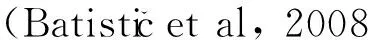
近年來,對哺乳動物棕櫚酰基轉移酶的研究發現,一些富含DHHC結構域的蛋白與人類某些疾病相關。如DHHC9被證明與早期智力殘疾相關的X-連鎖的精神遲鈍有關(Mitchell et al, 2014),膀胱癌和肝母細胞瘤的發生與DHHC11密切相關(Yamamoto et al, 2007;Wu et al, 2013),DHHC17(HIP14)負責亨廷頓蛋白(Htt)的棕櫚酰化,與亨廷頓舞蹈癥相關(Sanders et al, 2015)。高等植物基因組中也存在數目不等的DHHC蛋白基因,例如玉米(Zeamays)中有40個,楊樹(Populustrichocarpa)39個,水稻(Oryzasativa)30個,擬南芥(Arabidopsisthaliana)24個(Yuan et al, 2013)。但與哺乳動物和酵母相比,對植物PATs的研究還相對較少(Hemsley, 2013)。DHHC型鋅指蛋白在植物的生長發育過程中起著非常重要的作用。例如,從擬南芥突變體中分離得到的DHHC型鋅指蛋白TIPl,與酵母的Akrl和人的HIPl4有很大的相似性,它們都具有蛋白酰基轉移酶的活性。TIPl在擬南芥中參與調節植物細胞的生長,對植株的生長和株型發育具有重要調控作用(Hemsley et al, 2005)。AtPAT10(At3g51390)對植株的生長發育起到重要的作用,同時參與植株對外界鹽脅迫的響應過程(Zhou et al, 2013)。擬南芥DHHC型鋅指蛋白基因At5g04279和水稻DHHC型鋅指蛋白基因OsDHHC1與株型調控有關(Xiang et al, 2010;周波, 2011)。
木蘭科(Magnoliaceae)鵝掌楸屬(Liriodendron)現僅存鵝掌楸(L.chinense)和北美鵝掌楸(L.tulipifera)2個種。鵝掌楸主要分布于我國長江流域以南的亞熱帶中、低山區,其干形通直、樹形美觀、葉形奇特、花色艷麗、木材紋理通直、結構細密,材質輕軟,是珍貴用材與園林觀賞兼用樹種(蔡偉建等, 2011;姚俊修等, 2012)。本研究以鵝掌楸葉芽為材料,根據篩選到的DHHC鋅指蛋白家族基因的部分序列,采用RT-PCR和RACE技術克隆了3個鵝掌楸DHHC蛋白基因LcPAT7、LcPAT22、LcPAT23,并對其進行生物信息學分析。同時,采用實時熒光定量PCR技術檢測了3個基因在鵝掌楸不同組織中的表達特性,期望為鵝掌楸生長發育、形態建成以及逆境響應信號傳導等相關基因的調控研究提供參考。
1 材料與方法
1.1 植物材料
鵝掌楸(LS)采自南京林業大學下蜀實習林場的鵝掌楸屬種源試驗林,樹齡20 a,分別取其幼嫩葉片、花芽、葉芽和盛花期的花瓣、雄蕊、雌蕊,液氮速凍后置于-80 ℃超低溫冰箱保存備用。
1.2 總RNA的提取及cDNA第1鏈的合成
利用RNAprep Pure植物總RNA提取試劑盒(TIANGEN)分別提取上述6個不同組織樣品的總RNA,用紫外分光光度計檢測RNA的濃度和OD值,并取適量體積RNA用1%的瓊脂糖凝膠電泳檢測。
以葉芽總RNA為模板,按照RevertAid strand cDNA Synthesis Kit(Thermo Scientific)合成cDNA第1鏈,參照TaKaRa公司的3′-Full RACE Core Set with PrimeScript RTase和5′-Full RACE Kit試劑盒操作說明分別合成3′cDNA和5′cDNA;以6個不同組織樣品總RAN為模板,均一化濃度后,按照M-MLV First Strand Kit(Invitrogen)合成用于熒光定量PCR的cDNA,存于-20 ℃備用。
1.3 基因全長cDNA的克隆
(1)以NCBI的擬南芥TIP1基因序列(NC_003076.8)為信息探針,從本實驗室鵝掌楸轉錄組數據(Yang et al, 2014)中篩選出3個DHHC蛋白基因EST序列。利用Oligo 6軟件分別設計引物P7F1、P7R1,P22F1、P22R1和P23F1、P23R1(表1),送上海捷瑞生物公司合成。以cDNA模板,進行PCR擴增。反應體系(50 μL):10 × LA PCR Buffer II(Mg2+Free)5 μL,Mgcl2(25 mol·L-1)5 μL,dNTP Mixture(2.5 mol·L-1each)8 μL,引物各2 μL,TaKaRaLATaq(5 U·μL-1)0.5 μL,cDNA 2μL,無菌雙蒸水25.5 μL。擴增程序:預變性(94 ℃,3 min);變性(94 ℃,30 s),退火(30 s),延伸(72 ℃,2 min),30個循環;總延伸(72 ℃,10 min)。擴增3個基因的退火溫度分別59、56、58 ℃。將得到的PCR產物經2%瓊脂糖凝膠電泳檢測后,切下目的條帶,用膠回收試劑盒AxyPrep DNA Gel Extraction Kit(Axygen)回收,產物與pEASY-T1 Cloning Vector(TransGen Biotech)載體相連接,轉化Trans1-T1感受態細胞(TransGen Biotech),進行藍白斑篩選,挑取白斑進行菌落PCR鑒定后,將篩選到的陽性克隆菌液送往上海英駿生物技術有限公司測序,獲得3個基因的部分序列。
(2)根據(1)所得3個基因測序驗證的部分序列,利用Oligo 6軟件按照RACE試劑盒巢式PCR的要求分別設計3個基因3′末端和5′末端各2條末端擴增引物(表1)。
3′ RACE:以3′RACE cDNA為模板,用P7-3GSP1、P22-3GSP1、P23-3GSP1分別與3′Outer primer引物進行巢式第1輪PCR擴增,反應體系同3′RACE試劑盒說明書,擴增條件:預變性(94 ℃,3 min);變性(94 ℃,30 s),退火(56 ℃,30 s),延伸(72 ℃,2 min),25個循環;總延伸(72 ℃,10 min)。以第一輪PCR的產物為模板,用P7-3GSP2、P22-3GSP2、P23-3GSP2分別與3′Inner primer引物進行巢式第2輪PCR擴增,反應體系同3′RACE試劑盒說明書。擴增條件同上,退火溫度分別為58、61、62 ℃,共30個循環。克隆測序參照1.3的方法。
5′ RACE:以側芽5′RACE cDNA為模板,以5′RACE引物P7-5GSP1、P22-5GSP1、P23-5GSP1(表1)分別與5′Outer primer引物進行巢式第一輪PCR擴增,反應體系同5′RACE試劑盒說明書,擴增條件:預變性(94 ℃,3 min);變性(94 ℃,30 s),退火(58 ℃,30 s),延伸(72 ℃,2 min),25個循環;總延伸(72 ℃,10 min)。以第一輪PCR的產物為模板,用P7-5GSP2、P22-5GSP2、P23-5GSP2(表1)分別與5′Inner primer引物進行巢式第二輪PCR擴增,反應體系同5′RACE試劑盒說明書,擴增條件同上,退火溫度分別61、63、64 ℃,共30個循環。克隆測序參照1.3的方法。
1.4 基因開放閱讀框(ORF)預測及擴增
將1.3中擴增得到的序列在DNAMAN 6.0軟件中進行拼接獲得基因全長,利用在線分析工具FGENESH,對拼接得到的基因全長序列的ORF進行預測。將預測得到的ORF編碼的氨基酸序列在NCBI中進行比對以進一步確認ORF的準確性,并用Oligo 6設計引物(表1),以cDNA為模板,用2×TransStartFastPfuPCR SuperMix(TransGen Biotech)進行ORF擴增,反應體系(50 μL):2×TransStartFastPfuPCR SuperMix 25 μL,引物各2 μL,cDNA 2 μL,無菌雙蒸水19 μL。反應條件:預變性(94 ℃,3 min);變性(94 ℃,30 s),退火(30 s),延伸(72 ℃,2min),30個循環;總延伸(72 ℃,10 min)。退火溫度分別為60 ℃、58 ℃、58 ℃。克隆測序方法同1.3,產物用pEASY-Blunt克隆載體(TransGen Biotech)連接。

表 1 克隆用引物
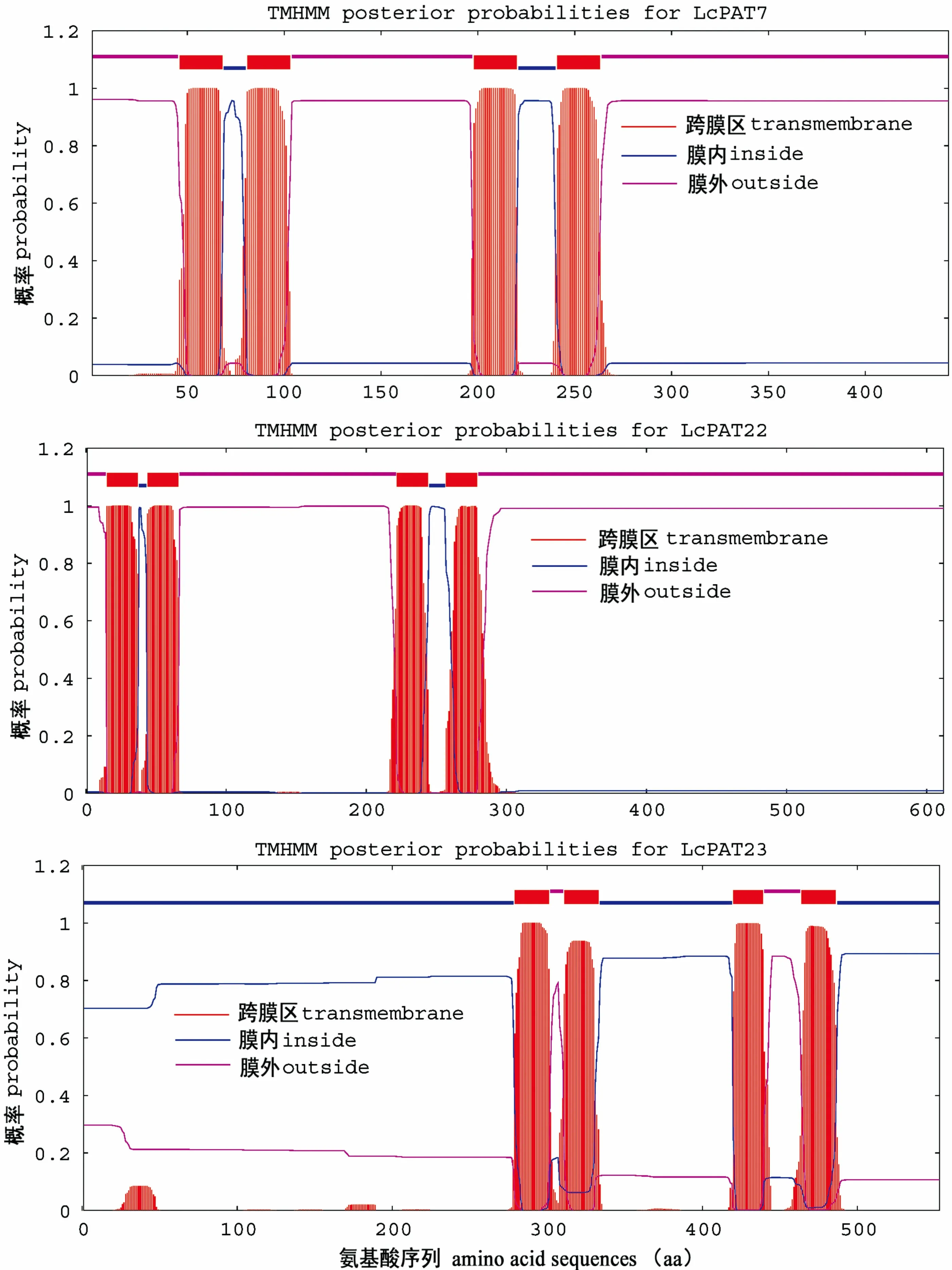
圖 1 LcPAT7、LcPAT22、LcPAT23的生物信息學分析 (跨膜結構分析)Fig. 1 Bioinformatics analysis of LcPATs (Prediction of transmembrane helices in proteins)
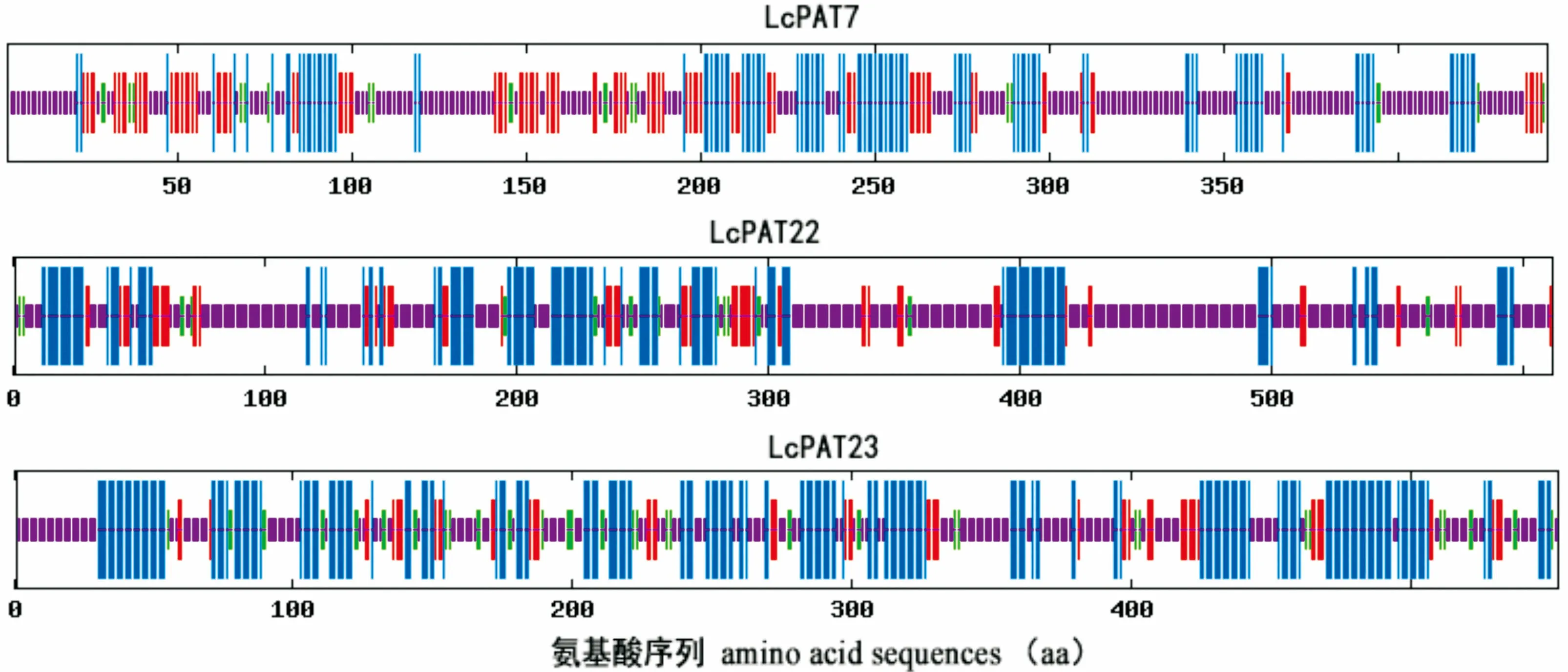
圖 2 LcPAT7、LcPAT22、LcPAT23的生物信息學分析 (二級結構預測)Fig. 2 Bioinformatics analysis of LcPATs (Secondary structure prediction)
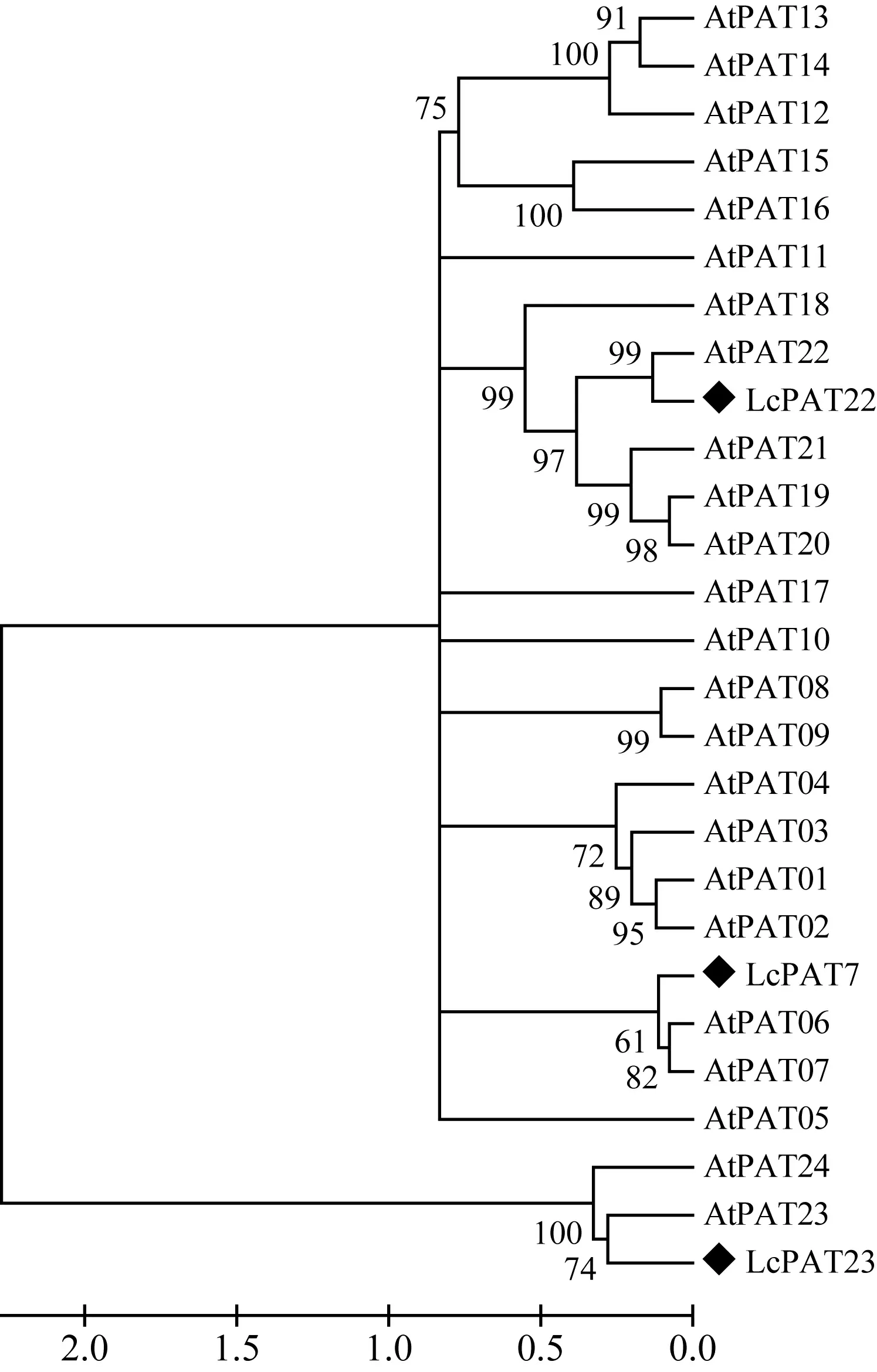
圖 3 LcPAT7、LcPAT22、LcPAT23與擬南芥PAT基因家族系統進化樹 At. 擬南芥; Lc. 鵝掌楸。 節點上的數值表示Bootstrap驗證中基于1 000次重復該節點可信度的百分比。Fig. 3 Phylogenetic tree of LcPATs and Arabidopsis thaliana PAT family At. A. thaliana; Lc. Liriodendron chinense. The number at the nodes represents the reliability percent (%) of bootstraps values based on 1 000 replications.
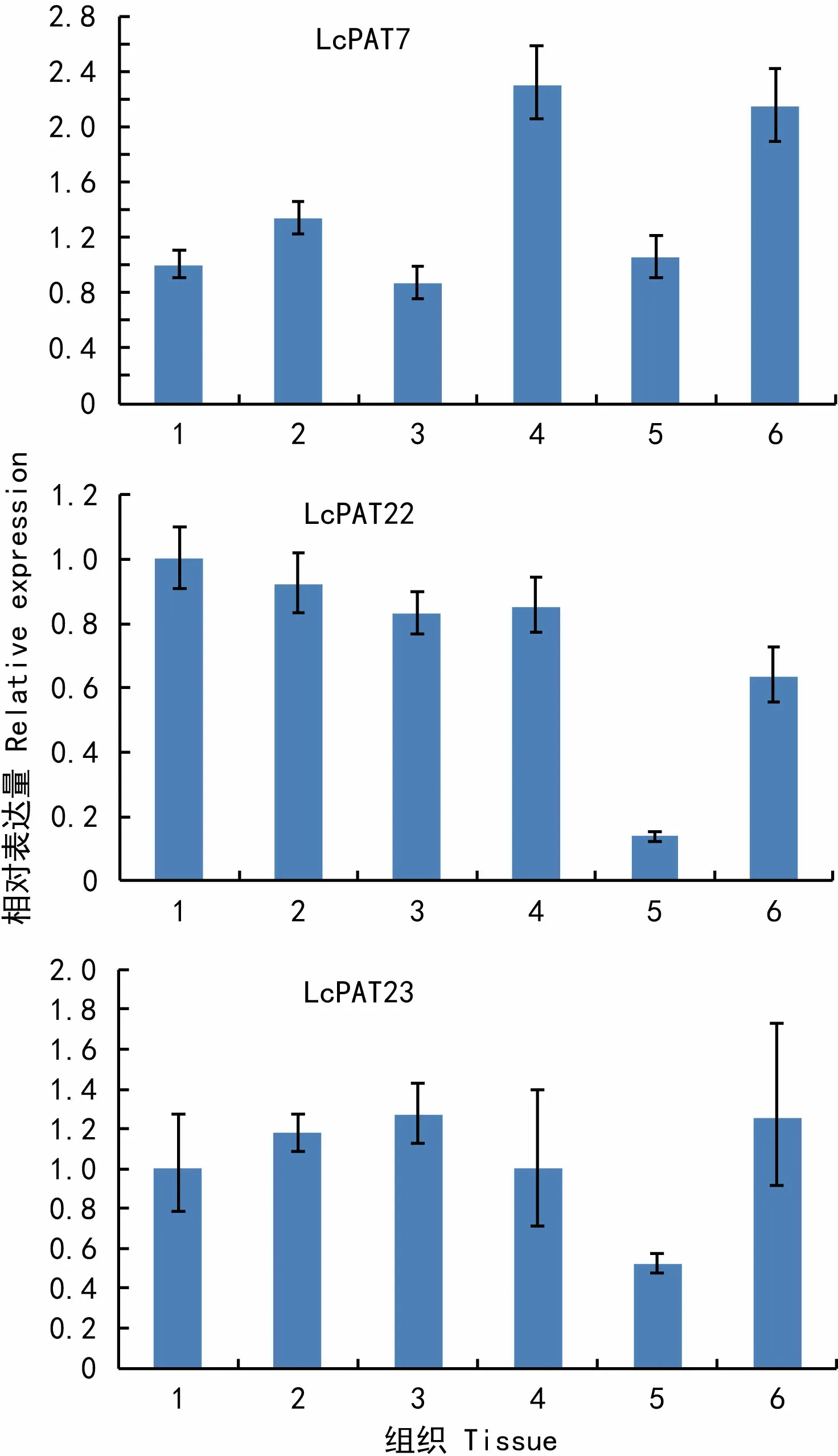
圖 4 LcPAT7、LcPAT22、LcPAT23在鵝掌楸不同組織中的表達情況 1. 葉芽; 2. 葉片; 3. 花芽; 4. 花瓣; 5. 雄蕊; 6. 雌蕊。Fig. 4 Expression level of PATs in different tissues of Liriodendron chinense 1. Leaf bud; 2. Leaf; 3. Floral bud; 4. Petal; 5. Stamen; 6. Pistil.
1.5 基因的組織表達分析
利用1.2所得到的cDNA進行實時定量PCR(quantitative real-time PCR,qRT-PCR)分析,以檢測目的基因在不同組織的相對表達量。采用Primer Express Software version 3.0(Applied Biosystems)軟件進行qRT-PCR引物設計,以鵝掌楸肌動蛋白基因Actin為內參,所用引物見表1。用SYBRPremixExTaq(Tli RNaseH Plus)(TaKaRa)實時熒光定量PCR試劑盒進行試驗,在ABI 7500 PCR儀上完成,每個反應重復3次。反應體系(20 μL):SYBRPremixExTaq(Tli RNaseH Plus)(2×)10 μL,ROX Reference Dye II(50×)0.4 μL,Forward/Reverse primer(10 μmol·L-1)0.4 μL,cDNA 2 μL,無菌雙蒸水6.8 μL。反應條件:預變性(95 ℃,30 s);變性(95 ℃,5 s),退火延伸(58 ℃,34 s),40個循環。
1.6 基因生物信息學分析
利用FGENESH(http://mendel.cs.rhul.ac.uk/mendel.php?topic=fgen)軟件進行ORF確定及氨基酸序列預測。用在線工具Expasy ProtParam(http://web.expasy.org/protparam/)分析蛋白的分子質量、理論等電點、氨基酸組成。通過Pfam數據庫(http://pfam.xfam.org/search)推測蛋白質結構域。使用SignalP 4.1 Server(http://www.cbs.dtu.dk/services/SignalP/)進行信號肽分析。利用TMHMM Server v.2.0(http://www.cbs.dtu.dk/services/TMHMM-2.0/)程序進行蛋白序列跨膜區分析。蛋白質二級結構分析采用SOPMA(http//npsa-pbil.ibcp.fr/gi-bin/npsa_automat.pl?page=/NPSA/npsa_sopma.html)工具進行,利用MEGA 5.1鄰接(Neighbor Joining,NJ)算法構建系統進化樹。
2 結果與分析
2.1 鵝掌楸DHHC蛋白基因全長克隆與序列分析
以鵝掌楸cDNA為模板,分別以P7F、P7R,P22F、P22R和P23F、P23R為引物,擴增得到1 358、2 070、1 687 bp的片段。通過3′RACE和5′RACE技術分別獲得cDNA的3′端(1 103、1 044、713 bp)和5′端(1 235、1 062、1 127 bp);采用DNAMAN 6.0軟件拼接出3個DHHC蛋白基因的全長cDNA序列(1 933、2 592、2 217 bp);利用在線工具FGENESH進行ORF預測,并在NCBI蛋白質數據庫中進行Blast比對。結果顯示,目的序列與其他植物中預測的PAT基因高度相似,并根據與NCBI中擬南芥PAT基因的比對結果,將其分別命名為LcPAT7(1 933 bp)、LcPAT22(2 592 bp)、LcPAT23(2 217 bp)。
設計引物P7OF、P7OR,P22OF、P22OR,P23OF、P23OR進行ORF擴增,測序驗證。結果表明,LcPAT7含一個1 332 bp的開放閱讀框,編碼443個氨基酸。其預測蛋白的分子量為49.04 kDa,理論等電點為9.15。酸性氨基酸(Asp,Glu)總數35個,堿性氨基酸(Arg,Lys)總數46個;GRAVY(grand average of hydropathicity)值為-0.021;不穩定指數為32.08,該蛋白質為穩定蛋白。LcPAT22含一個1 839 bp的開放閱讀框,編碼612個氨基酸,其預測蛋白的等電點和分子質量分別為9.03和67.3 kDa。酸性氨基酸(Asp,Glu)總數53個,堿性氨基酸(Arg,Lys)總數67個;GRAVY值為-0.174;不穩定指數為53.92,該蛋白質為不穩定蛋白。LcPAT23含一個1 662 bp的開放閱讀框,編碼553個氨基酸,其預測蛋白的等電點和分子質量分別為7.29和60.57 kDa。酸性氨基酸(Asp,Glu)總數50個,堿性氨基酸(Arg,Lys)總數50個;GRAVY值為-0.080;不穩定指數為32.83,該蛋白質為穩定蛋白。
2.2 蛋白質結構分析及系統進化樹構建
以鵝掌楸葉芽cDNA為模板分離得到的3個DHHC蛋白基因,通過SignalP 4.1對3個基因進行信號肽預測。結果表明,該3個DHHC蛋白基因均不存在信號肽酶切位點,都屬非分泌型蛋白。蛋白質跨膜區分析結果表明,3個基因所編碼蛋白均具有4個跨膜區(圖1)。Pfam數據庫檢索顯示,LcPAT7、LcPAT22和LcPAT23均含有zf-DHHC結構域,且均位于跨膜域TM2和TM3之間,其中LcPAT23在N端有2個錨蛋白重復序列(Ank_2)。此外,3個基因所編碼蛋白均具有DHHC蛋白家族典型的結構域:C-X2-C-X9-HC-X2-C-X2-C-X4-DHHC-X5-C-X4-N-X3-F-X4(C代表半胱氨酸,H代表組氨酸,X代表任意氨基酸)(Putilina et al, 1999),說明其編碼的蛋白屬于DHHC蛋白家族成員。
SOPMA分析表明,α螺旋(Alpha helix)和無規則卷曲(Random coil)是這3個DHHC蛋白二級結構的主要成分。LcPAT7蛋白二級結構中含有23.70%的α螺旋、20.99%的延伸鏈(Extended strand)、4.74%的β轉角(Beta turn)和50.56%的無規則卷曲;LcPAT22蛋白二級結構中含有25.33%的α螺旋、11.11%的延伸鏈3.43%的β轉角和60.13%的無規則卷曲;LcPAT23蛋白二級結構中含有41.05%的α螺旋、10.49%的延伸鏈8.32%的β轉角和40.14%的無規則卷曲(圖2)。ProtComp 9.0蛋白亞細胞定位分析表明,LcPAT7定位于質膜,與AtPAT7一致;LcPAT22定位于質膜;而LcPAT23定位于高爾基體,與AtPAT23和AtPAT24(TIP1)定位一致。
通過NCBI上的protein blast進行同源性搜索,發現與它們同源的序列大部分都為預測的蛋白質酰基轉移酶(PAT),LcPAT7與蓮(Nelumbonucifera)、葡萄(Vitisvinifera)、大豆(Glycinemax)、煙草(Solanumtuberosum)、擬南芥(A.thaliana)中預測的PAT7相似性分別為81%、77%、74%、74%、70%;LcPAT22與蓮、葡萄、胡楊(Populuseuphratica)、海棗(Phoenixdactylifera)、煙草、擬南芥中預測的PAT22相似性分別為72%、70%、68%、67%、64%、61%;LcPAT23與蓮、海棗、葡萄、大豆、擬南芥中預測的PAT23相似性分別為77%、75%、72%、71%、60%。
應用Clustal W軟件將3個鵝掌楸DHHC蛋白家族基因與NCBI數據庫下載的24個擬南芥PATs蛋白進行多重比對,再利用MEGA 5.1軟件中的鄰接法構建系統進化樹,結果見圖3。從圖3可看出,LcPAT7與擬南芥中的AtPAT6和AtPAT7關系最近,LcPAT22與擬南芥中的AtPAT22關系最近,LcPAT23與擬南芥中的AtPAT23關系最近,且LcPAT23與AtPAT24(TIP1)歸為同一個小的分支。這為推測LcPAT23的功能提供了一定的線索。TIP1是擬南芥中第一個進行功能研究的蛋白質棕櫚酰基轉移酶,研究發現TIP1突變體不僅在根毛及花粉管的生長上表現出異常,還表現出節間縮短、植株矮小、蓮座葉變小、葉表皮細胞變小,且早花早衰,說明TIP1基因不但參與極性細胞的生長過程,同時還參與多種細胞的發育過程(Hemsley et al, 2005)。
2.3 DHHC基因在鵝掌楸不同組織中的表達特性
利用實時熒光定量PCR技術對3個基因在鵝掌楸不同組織中的表達模式進行分析,結果表明(圖4),3個基因在鵝掌楸葉芽、花瓣、雌蕊、花芽、雄蕊、葉片中均有表達,但表達豐度存在著明顯的差異。LcPAT7在花瓣中表達豐度最高,其次為雌蕊,在花芽和葉芽中的表達豐度相對較低;LcPAT22在葉芽中的相對表達豐度最高,葉片和花芽中次之,雌蕊中的表達豐度較低,而雄蕊中最低;LcPAT23在花芽和雌蕊中表達豐度較高,葉片、花瓣、花芽中表達豐度相差不大,而在雄蕊中表達豐度較低。
3 討論
本研究首次從鵝掌楸中克隆得到3個DHHC蛋白家族基因LcPAT7、LcPAT22、LcPAT23的全長cDNA序列。其編碼的蛋白均具有4個跨膜區,這與已報導的動植物DHHC蛋白跨膜域數目(4~6)一致,并且均在跨膜域TM2和TM3之間存在一個DHHC蛋白家族典型的結構域:C-X2-C-X9-HC-X2-C-X2-C-X4-DHHC-X5-C-X4-N-X3-F-X4,該結構域是動植物中S-酰基轉移酶的活性位點(Putilina et al, 1999)。DHHC中的半胱氨酸(Cysteine,C)殘基突變為絲氨酸(Serine,S)后,不會改變蛋白質的空間結構,但是會導致棕櫚酰基轉移酶喪失酶活性,表明該高度保守的半胱氨酸殘基是酶活性的關鍵決定位點(Greaves & Chamerlain, 2011)。
高等生物的基因表達具有組織特異性和發育階段特異性,不同基因隨時間、空間選擇性表達。基因的時空表達決定了生命的所有過程,如發育和分化、對逆境的反應、細胞分裂、衰老等。王倩等(2011)對擬南芥PAT基因家族中20個基因的研究表明:AtPAT7(At3g26935)在花、成熟果莢、莖中有表達,在花中表達量較高其他組織中表達量很低;AtPAT22(At1g69420)在果莢、花、幼苗、莖中有表達,在成熟果莢和花中表達量較高;AtPAT23(At2g14255)在幼苗、果莢、花、根、葉、莖中均有表達,在幼苗中表達量較高。熒光實時定量PCR分析結果表明,鵝掌楸LcPATs表達具有組織的差異性。LcPAT7、LcPAT22、LcPAT23在鵝掌楸花芽和盛花期的葉片、葉芽、花瓣、雄蕊、雌蕊中均有表達。LcPAT7在花瓣和雌蕊中表達豐度最高,功能上可能與AtPAT7具有一定的相似性,參與花瓣的擴展、心皮及胚的發育;LcPAT23在花芽和雌蕊中表達量相對較高,可能參與花發育過程;LcPAT22在葉芽中的表達量最高,其次是葉片、花芽,相對于LcPAT7、LcPAT23的表達模式,LcPAT22在葉芽、花芽中的表達豐度較其他組織高,可能參與莖端分生組織的生長調節。
ABANKWA D, GORFE AA, HANCOCK JF, 2007. Ras nanoclusters: molecular structure and assembly [J]. Semin Cell Dev Biol, 18(5): 599-607.

CAI JW, GUO X, GAO HD, et al, 2011. Research advances on cultivation ofLiriodendronplantation [J]. J Fujian For Sci Technol, 38(2): 164-170. [蔡偉建, 郭鑫, 高捍東, 等, 2011. 鵝掌楸屬植物人工林培育研究進展 [J]. 福建林業科技, 38(2): 164-170.]CHAROLLAIS J, VAN DER GOOT FG, 2009. Palmitoylation of membrane proteins (Review) [J]. Mol Membr Biol, 26(1): 55-66.
CHOI Y, LEE Y, HWANG J, 2014.ArabidopsisROP9 and ROP10 GTPases differentially regulate auxin and ABA responses [J]. J Plant Biol, 57(4): 245-254.
GREAVES J, CHAMBERLAIN LH, 2011. DHHC palmitoyl transferases: substrate interactions and (patho)physiology [J]. Trends Biochem Sci, 36(5): 245-253.
GUAN XM, FIERKE CA, 2011. Understanding protein palmitoylation:biological significance and enzymology [J]. Sci Chin Chem, 54(12): 1 888-1 897.
HEMSLEY PA, KEMP AC, GRIERSON CS, 2005. The TIP GROWTH DEFECTIVE1 S-acyl transferase regulates plant cell growth inArabidopsis[J]. Plant Cell, 17(9): 2 554-2 563.
HEMSLEY PA, 2013. Assayingprotein S-acylation in plants [J]. Meth Mol Biol, 1 043: 141-146.
HUANG K, EL-HUSSEINI A, 2005. Modulation of neuronal protein trafficking and function by palmitoylation [J]. Curr Opin Neurobiol, 15(5): 527-535.
LAVY M, YALOVSKY S, 2006. Association ofArabidopsistype-II ROPs with the plasma membrane requires a conserved C-terminal sequence motif and a proximal polybasic domain [J]. Plant J, 46(6): 934-947.
LECLERCQ J, RANTY B, SANCHEZ-BALLESTA MT, et al, 2005. Molecular and biochemical characterization of LeCRK1, a ripening-associated tomato CDPK-related kinase [J]. J Exp Bot, 56(409): 25-35.
LINDER ME, DESCHENES RJ, 2007. Palmitoylation: policing protein stability and traffic [J]. Nat Rev Mol Cell Biol, 8(1): 74-84.
MITCHELL DA, HAMEL LD, REDDY KD, et al, 2014. Mutations in the X-linked intellectual disability gene, zDHHC9, alter autopalmitoylation activity by distinct mechanisms [J]. J Biol Chem, 289(26): 18 582-18 592.
MITCHELL DA, VASUDEVAN A, LINDER ME, et al, 2006. Thematic review series: lipid posttranslational modifications. Protein palmitoylation by a family of DHHC protein S-acyltransferases [J]. J Lipid Res, 47(6): 1 118-1 127.
PEDRO B, VéRONIQUE A, LILLIAN RK, et al, 2012. Systematic functional prioritization of protein posttranslational modifications [J]. Cell, 150(2): 413-425.
PUTILINA T, WONG P, GENTLEMAN S, 1999. The DHHC domain: a new highly conserved cysteine-rich motif [J]. Mol Cell Biochem, 195(1-2): 219-226.
RESH MD, 2006. Trafficking and signaling by fatty-acylated and prenylated proteins [J]. Nat Chem Biol, 2(11): 584-590.
ROTH AF, FENG Y, CHEN L, et al. 2002. The yeast DHHC cysteine-rich domain protein Akr1p is a palmitoyl transferase [J]. J Cell Biol, 159(1): 23-28.RUAN BJ, DAI P, WANG W, et al, 2014. Progress on post-translational modification of proteins [J]. Chin J Cell Biol, 36(7): 1 027-1 037. [阮班軍, 代鵬, 王偉, 等, 2014. 蛋白質翻譯后修飾研究進展 [J]. 中國細胞生物學學報, 36(7): 1 027-1 037.]
SANDERS SS, HOU J, SUTTON LM, et al, 2015. Huntingtin interacting proteins 14 and 14-like are required for chorioallantoic fusion during early placental development [J]. Dev Biol, 397(2): 257-266.SCH?NICHEN A, WEBB BA, JACOBSON MP, et al, 2013. Considering protonation as a posttranslational modification regulating protein structure and function [J]. Ann Rev Biophys, 42: 289-314.
SUN PG, MIAO HX, XU BY, et al. 2013. Proceedings in biological function of small GTPases gene ROP of plant [J]. North Hortic, (22): 188-192. [孫佩光, 苗紅霞, 徐碧玉, 等. 2013. 植物小G蛋白基因ROP生物學功能研究進展 [J]. 北方園藝, (22): 188-192.]WANG Q, SUN JL, BAO L, et al, 2011. Twenty putative palmitoyl-acyl transferase genes show distinct expression pattern inArabidopsisthaliana[J]. Afr J Biotechnol, 10(52): 10 575-10 584.
WOLFF J, ZAMBITO AM, BRITTO PJ, et al, 2000. Autopalmitoylation of tubulin [J]. Protein Sci, 9(7): 1 357-1 364.
WU JF, LEE CH, CHEN HL, et al, 2013. Copy-number variations in hepatoblastoma associate with unique clinical features [J]. Hepatol Int, 7(1): 208-214.
XIANG J, LIN JZ, TANG DY, et al, 2010. A DHHC-type zinc finger protein gene regulates shoot branching inArabidopsis[J]. Afr J Biotechnol, 9(45): 7 759-7 766.
YAMAMOTO Y, CHOCHI Y, MATSUYAMA H, et al, 2007. Gain of 5p15.33 is associated with progression of bladder cancer [J]. Oncology, 72(1-2): 132-138.
YANG Y, XU M, LUO QF, et al, 2014. De novo transcriptome analysis ofLiriodendronchinensepetals and leaves by Illumina sequencing [J]. Gene, 534(2): 155-162.
YAO JX, LI HG, BIAN LM, et al, 2012. Inbreeding depression inLiriodendronrevealed by both phenotypic and genotypic characteristics [J]. J NE For Univ, 40(4): 1-5. [姚俊修, 李火根, 邊黎明, 等, 2012. 鵝掌楸屬樹種近交衰退分析 [J]. 東北林業大學學報, 40(4): 1-5.]
YUAN XW, ZHANG SZ, SUN MH, et al, 2013. Putative DHHC-cysteine-rich domain S-acyltransferase in plants [J]. PLoS ONE, 8(10): e75985.
ZHOU B, 2011. Function analysis of the DHHC type znic finger protein geneOsDHHC1 in the plant architecture construction of rice(Oryzasativa L.) [D]. Changsha:Hunan University. [周波, 2011. DHHC型鋅指蛋白基因OsDHHC1在水稻株型構建中的功能分析 [D]. 長沙:湖南大學.]
ZHOU LZ, LI S, FENG QN, et al, 2013. Protein S-ACYL Transferase10 is critical for development and salt tolerance inArabidopsis[J]. Plant Cell, 25(3): 1 093-1 107.
Gene cloning and expression analysis of DHHC protein family genes fromLiriodendronchinense
XU Jia-Juan, LI Huo-Gen
(KeyLabofForestGeneticsandBiotechnology,NanjingForestryUniversity,MinistryofEducation, Nanjing 210037, China )
Protein S-acylation is a common and unique reversible way in posttranslational lipid modification, thus confers diverse physiological functions on target proteins, such as DHHC (Asp-His-His-Cys) protein family. There was evidence that the DHHC domain was directly involved in the palmitoyl transfer reaction. In this paper, we reports gene cloning and expression analysis of DHHC protein family genes inLiriodendronchinense. Three full-length cDNA of DHHC gene were cloned fromLiriodendronleaf buds using the rapid amplification of cDNA ends (RACE) strategy, namedLcPAT7,LcPAT22 andLcPAT23 respectively. The full-length cDNAs ofLcPAT7,LcPAT22 andLcPAT23 were 1 933 bp, 2 592 bp and 2 217 bp, and contain 1 332 bp, 1 839 bp and 1 662 bp ORF, encoding 433, 612 and 533 amino acids, respectively. The predicted molecular weights of the proteins encoded byLcPAT7,LcPAT22 andLcPAT23 were 40.04, 67.3 and 60.57 kDa, respectively. The predicted isoelectric points are 9.15, 9.03 and 7.29, respectively. All the three proteins contained four putative transmembrane (TM) domain structures, and contained a typically DHHC-CRD domain between TM2 and TM3 as most known DHHC protein. Homology analysis showed that the three genes showed high similarity with predicted PAT (protein S-acyltransferase) of other plants. Tissue expression profile by Real-time PCR showed that all the three genes were expressed in various tissues, although the expression levels varied significantly in different tissues. Changes in gene expression patterns in the same gene family indicated their functionally non-redundancy. These results above will provide clues for exploring the underlying mechanism of gene regulation on growth, development, morphogenesis and signal transduction of stress response inL.chinense.
Liriodendronchinense, protein S-acylation, protein S-acyltransferase (PAT), DHHC protein family, expression analysis
10.11931/guihaia.gxzw201503026
2015-03-17
2015-04-13
國家自然科學基金(31470660,31170621);江蘇省高校優勢學科[Supported by the National Natural Science Foundation of China (31470660,31170621); Priority Academic Program Development of Jiangsu High Education Institutions]。
徐嘉娟(1988-),女,云南臨滄人,碩士研究生,研究方向為林木分子遺傳,(E-mail)rrylxjj@163.com
李火根,教授,研究方向為林木遺傳育種,(E-mail)hgli@njfu.edu.cn.
Q943
A
1000-3142(2016)09-1052-09
徐嘉娟, 李火根. 鵝掌楸DHHC型鋅指蛋白家族基因的克隆及表達分析 [J]. 廣西植物, 2016, 36(9):1052-1060
XU JJ, LI HG. Gene cloning and expression analysis of DHHC protein family genes fromLiriodendronchinense[J]. Guihaia, 2016, 36(9):1052-1060
