管角螺代謝物對浮游植物增殖的影響
2016-11-23 01:56:06羅杰劉皓劉楚吾曹伏君蘇棟杰
水產養殖 2016年10期
關鍵詞:生長
羅杰,劉皓,劉楚吾,曹伏君,蘇棟杰
(廣東海洋大學水產學院,南海水產經濟動物增養殖廣東普通高校重點實驗室,廣東湛江 524025)
管角螺代謝物對浮游植物增殖的影響
羅杰,劉皓,劉楚吾,曹伏君,蘇棟杰
(廣東海洋大學水產學院,南海水產經濟動物增養殖廣東普通高校重點實驗室,廣東湛江 524025)
采集管角螺的糞便,烘干處理后測定其營養成分組成;并在實驗室條件下,研究了管角螺(Hemifusus tuba)代謝物對湛江等鞭金藻(Isochrysis zhanjiangensis)、牟氏角毛藻(Chaetoceros muelleri)和亞心型扁藻(Platymonas subcordiformis)生長的影響。結果表明:管角螺糞便中含有粗蛋白質(9.8%)、磷(0.38%)等微藻生長所需的營養物質;管角螺的代謝物對三種單胞藻的生長有明顯的促進作用(P<0.05),三種微藻的生長與代謝物的添加量呈正比相關;管角螺代謝物對不同微藻的生長作用隨種類的不同而具有明顯的差異,對湛江等鞭金藻和牟氏角毛藻的增殖作用明顯高于亞心型扁藻(P<0.05)。
管角螺;代謝物;微藻;生長
近年來我國海水養殖業發展迅猛,產量已經躍居世界第一,其中貝類產量在海水養殖產量中的比例逐年上升,2014年達到1 341.67萬t,已經成為海水養殖產業和沿海地區經濟發展的支柱產業[1]。但隨著局部海區過度開發,使得某些海區自身污染加重,生態系統失調,養殖總量超過水域容納量,加上我國的養殖海區由于養殖的魚、貝排泄物以及陸地養殖對蝦的廢水向海區的過度排放,造成了海區的富營養化,一些主要養殖海區赤潮發生頻繁,環境污染加劇,病害面積不斷擴大,每年造成巨大的經濟損失。
蔣艾青等[2]在研究水生生物的排泄物對水中有機物影響時指出,魚類1 g排泄物產生的NO2-N要比飼料多,1 g排泄物所產生的NH3-N、COD比死魚多。在我國的一些地區采取海藻與貝類混養的海洋生態鏈的養殖模式,其主要原理是藻類可以利用海水中的碳、氮、硫、磷等物質作為自身的營養物質,而貝類(主要為濾食性貝類)濾食海洋中的一些微小生物,同時其排出的廢物被藻類吸收,促進藻類的生長繁殖,對海洋生態環境起到調節作用[3-5]。微藻是海洋中的微小浮游植物,處于海洋生物食物鏈中最底層,但卻是海洋初級生產者和能量的主要轉換者,它的數量變動直接或間接地影響魚、蝦、貝及其幼體的存活和生長,因而通過研究浮游植物的數量變動可以對漁業產量進行估算[6-7]。目前一些學者在研究養殖生物對淺海生態系統的影響及評估不同海域的養殖容量時,大多數集中在濾食性貝類和浮游植物食性魚類方面的研究[8-9]。王俊等[8]研究指出,濾食性貝類菲律賓蛤仔(Ruditapes philippinarum)、太平洋牡蠣(Crassastrea gigas)和櫛孔扇貝(Chlamys farreri)對浮游植物有明顯的促生長作用,并且浮游植物的增長與貝類密度呈正相關。而對肉食性貝類影響生態系統方面的研究報道較少。
一些肉食性貝類雖然不能直接利用水中的微小生物,但其排泄物中有一部分營養成分能夠促進藻類的生長。管角螺為一種腹足類軟體動物,生活在淺海潮間帶,以肉食性為主,大小規格可影響其排氨率[10-12],從而直接影響養殖水體的氨氮含量。氮是海洋植物所需的基本營養元素,其在水中的含量對藻類的豐度影響較大。湛江等鞭金藻(Isochrysis zhanjiangensis)、牟氏角毛藻(Chaetoceros muelleri)和亞心型扁藻(Platymonas subcordiformis)是自然海域中常見種類三種微藻,在水產動物的人工育苗階段作為餌料被廣泛應用。實驗通過采集管角螺的排泄物,烘干處理后送檢測定其糞便營養成分組成,并以生態學的方法研究了管角螺代謝物對湛江等鞭金藻、牟氏角毛藻和亞心型扁藻三種微藻生長的影響,為綜合研究肉食性貝類對海區浮游藻類數量動態的變化,建立持續的海水養殖模式提供理論依據。
1 材料與方法
1.1 實驗材料和儀器
實驗用的管角螺來自人工育苗,在水泥池養殖6個月的幼螺。挑選活力好、外表無損傷的個體,平均殼高6.9 cm的管角螺20個,清除掉表面的附著物,放入水族箱中暫養一個星期,飼養期間投喂足量的貝類、小雜魚;每天清洗缸底后換水100%。湛江等鞭金藻、角毛藻及亞心形扁藻為廣東海洋大學藻類實驗室一級培養的單胞藻,均處于指數生長期。海水取自東海島海洋生物研究基地,無污染,水溫28~30.5℃,鹽度28~30,pH值為8.2。海水經沉淀、砂濾后使用。
1 000 mL量筒、1 000 mL的三角燒瓶,經高溫消毒后備用;15 L玻璃水槽。
1.2 方法
1.2.1 管角螺糞便采集水族箱養殖的管角螺,剛產出的糞便明顯,時間長了由于海水的浸泡和水流的影響很容易散開。因此,在換水前用吸管輕輕吸取糞便于濾紙上,然后糞便在40℃環境下干燥處理24 h。連續取樣5 d。
1.2.2 實驗分組和微藻培養取20 L經過砂濾的自然海水煮沸,冷卻后在容量為15 L并經過消毒處理的玻璃水槽中加入10 L,然后放入20個經暫養消毒過的管角螺進行養殖,期間不投喂餌料,24 h后取出,將養殖過管角螺的海水用300目篩絹過濾去除雜質。
金藻實驗組,用1 000 mL三角燒瓶分別量取上述飼養過管角螺的海水100 mL,按0%、25%、50%、75%、100%的比例加入已經煮沸過的自然海水(不添加營養鹽)與之混合,這樣調配好的海水容量分別為100、125、150、175和200 mL,對應的編號為A、B、C、D和E,然后往編號為A、B、C、D和E的三角燒瓶接種處于指數生長期的湛江等鞭金藻,并使每個瓶的金藻密度約為12.5×104cell/mL。牟氏角毛藻、亞心形扁藻實驗組,按上述方法調配好A、B、C、D、E五瓶混合海水,然后分別接種處于指數生長期的角毛藻和亞心形扁藻,密度分別為8.6×104cell/mL、5.3×104cell/mL。每個實驗組設三個平行組,結果取平均值。
1.2.3 單胞藻的管理和藻細胞密度的測定微藻培養的光照為正常光照時間,每天搖動三角燒瓶2~3次,防止藻體沉積瓶底;實驗開始后24、48、72、96、120、144和168 h,每天11:00時用移液槍各取1 mL藻液,加入適量的碘液殺死藻細胞,搖晃使藻細胞在水體中分布均勻,用移液管迅速吸出,采用血球計數板法測定細胞密度[13-14],每個樣品重復計數3次,然后取平均值。
1.2.4 數據處理與分析采用SPSS11.5統計軟件對數據進行分析處理,并進行多重分析比較,有統計學意義水平規定為P﹤0.05。實驗數據均表示為平均值±標準差(Mean±SE)。
相對最大生長率=(最大藻細胞密度-初始藻細胞密度)/初始藻細胞密度×100%
2 結果
2.1 管角螺糞便組成
把烘干的管角螺糞便進行檢測,糞便中的組成成分主要有粗蛋白質、粗脂肪、粗纖維、中性纖維、鈣、磷和纖維,還有鐵、銅等微量元素及氨基酸。在這些成分中,粗蛋白質、磷和一些微量元素如鐵是微藻生長的營養物質(見表1)。
2.2 管角螺對單胞藻的促長作用
從圖1中可以看出,管角螺代謝物對湛江等鞭金藻、牟氏角毛藻和亞心型扁藻三種微藻的生長具有較為明顯的促進作用。在培養過管角螺一定容量的海水中,比較按0%、25%,50%、75%、100%的比例分別加入自然海水,三種微藻的生長曲線基本一致,生長率隨著自然海水增加量的增加而降低,同時維持生長的時間也相對縮短。沒有加入自然海水(0%)一組,微藻的生長速度最快,也獲得最大生長率,其指數生長期明顯,72 h達到生長高峰,維持生長的時間最長,達到144 h;而加入100%自然海水組,細胞密度增長不明顯,只維持生長48 h或72 h其生長就相對下降,然后進入死亡期。通過分析比較,湛江等鞭金藻、牟氏角毛藻和亞心型扁藻組5個系列培養其細胞生長存在顯著差異(P﹤0.05)。

表1 管角螺糞便主要成分%
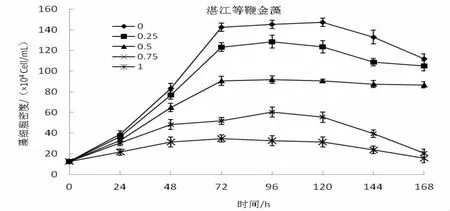
圖1 管角螺代謝物對湛江等鞭藻增長的影響
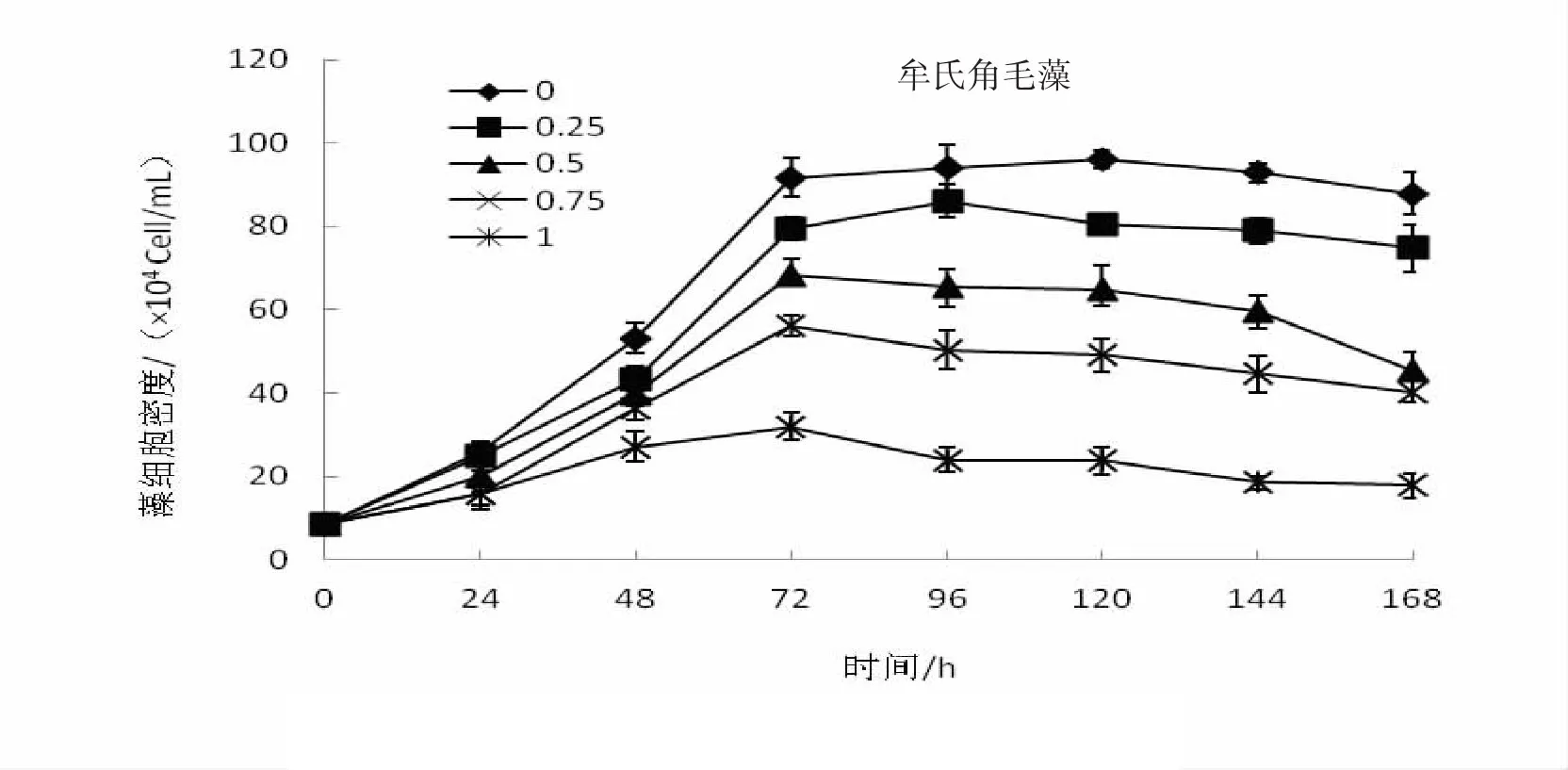
圖2 管角螺代謝物對牟氏角毛藻增長的影響
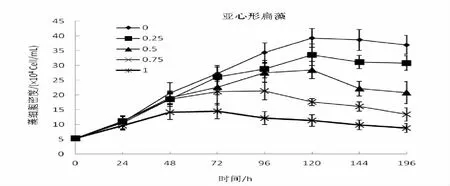
圖3 管角螺代謝物對亞心形扁藻增長的影響
同時,管角螺代謝物對單胞藻的促長作用隨種類的不同而具有較大的差異。湛江等鞭金藻組,各系列培養24 h細胞的生長即出現較為明顯的差別,加入0%自然海水系列72 h達到生長高峰,細胞密度為142.5×104(cells/mL),相對最大生長率達1077.36%,144 h出現死亡期;25%,50%、75%和100%系列,相對最大生長率分別為928%、645.04%、381.10%和178.4%,死亡時間為120、120、120和96 h。牟氏角毛藻,相對最大生長率為1 016.28%、876.74%、697.68%、553.84%、271.28%,出現死亡期分別為144、120、96、96和96 h;培養亞心型扁藻,24 h內藻細胞的生長差別不大,但隨著培養時間的增長,細胞增長量出現較為明顯的差別,其相對最大生長率641.51%、533.97%、437.74%、303.02%和130.19%,0%的最大生長率為100%的近5倍,死亡期出現在144、144、144、120和96 h。分析比較,管角螺代謝物培養這三種單胞藻其生長差異有統計學意義(P﹤0.05)。
3 結論與討論
管角螺糞便含有粗蛋白質、粗脂肪、粗纖維、中性纖維、鈣、磷和纖維,還有鐵、銅等微量元素及氨基酸,其溶于水體中的氮、磷是微藻生長的營養元素。管角螺代謝產物對湛江等鞭金藻、牟氏角毛藻和亞心型扁藻三種微藻都有很好的促生長作用,但對湛江等鞭金藻、牟氏角毛藻促生長效果明顯優于亞心型扁藻。
3.1 管角螺糞便的營養成分分析
天然水體中可溶性無機化合物氮、磷、硅元素在水生植物的生長繁殖過程中被吸收利用,成為生物體的重要組成元素,被視為水生植物生長所需的營養鹽。但這些元素在天然水體中的含量通常很低,遠遠不如構成生物體的其他元素那樣豐富。各種微藻類所需要的營養物質不盡相同,營養鹽的需求量隨種類的不同而有很大的差異,只有在滿足藻類對營養鹽需求的條件下,藻類才會有較高的增長率[9,15]。在浮游植物大量繁殖季節,他們有效形態的含量甚至降至吸收臨界值之下,從而影響藻類的生長繁殖,限制了水體初級生產的速率和產量。有些貝類是濾食性,主要濾食水體中的微小顆粒,包括一些微藻、細菌及有機碎屑等,而有些貝類是肉食性,如東風螺、管角螺,主要攝食一些動物的腐敗尸體或小型貝類,由于攝食的對象不同,產出的糞便所含的營養成分也存在差異。柴雪良等[16]研究指出,腹足類的代謝產物主要有氨、尿酸、尿素、氨基酸等,其中氨占總排量的70%或更高;吳小平等[17]研究了魚類糞便營養成分,其營養組成成分(干物質)組成如下:粗蛋白質10%~13%、粗脂肪0.9%~1.6%、粗纖維24%~26%,磷0.50%~1.1%,還有鐵、銅、鋅、鉀、鉻等微量元素及二十余種氨基酸。通過對管角螺糞便檢測,粗蛋白含量為9.8%,磷為0.38%,N/P接近28,而微藻生長所需的營養元素主要為氮、磷,這兩種營養物質在水中的含量直接影響微藻的繁殖生長速度及豐度。
3.2 管角螺螺代謝物對微藻的促生長作用
藻類分布的范圍極廣,對環境條件要求不嚴,適應性較強,在只有極低的營養濃度、極微弱的光照強度和相當低的溫度下也能生活。但是營養元素的不足會限制藻類吸收營養元素的速率,從而限制浮游植物的總產量。在培養液中加入藻類生長所需的營養鹽,藻細胞繁殖生長表現出明顯的特定模式:延緩期、指數生長期、生長相對下降期、靜止期和死亡期5個階段,期間某個時期可能持續時間很短,表現不是十分明顯[14]。用管角螺代謝物培養湛江等鞭金藻、牟氏角毛藻和亞心型扁藻,微藻生長的上述5個階段較為明顯,特別是濃度較高的0%組,三種微藻最大生長密度分別為剛接種時的11.8倍、11.2倍和7.4倍,生長時間可達120 h;而加入100%自然海水的組,三種微藻生長的緩慢,曲線平穩,很快進入靜止期,最大生長密度分別為剛接種時的2.6倍、2.8倍和2.7倍。管角螺代謝物對三種微藻促長作用效果不同,這主要取決于藻類對營養鹽的需求量。很多學者認為,單胞藻的繁殖生長除取決于氮、磷營養鹽濃度外,還與培養水體中N/P比值相關,過高或過低均對微藻生長不利[18-21]。林忠洲等[22]在研究不同氮磷質量濃度下青島大扁藻和牟氏角毛藻的種間競爭關系時發現,青島大扁藻對磷有較高的需求,而牟氏角毛藻對較低的磷濃度具有更好的適應性;孫育平等[23]研究得出亞心形扁藻最適N/P比值為17,Austin等[24]、于瑾等[25]認為牟氏角毛藻最佳N/P比值分別為28、23,均說明扁藻在磷濃度較高的環境下才能生長好。張青田等[26]研究了水體中氮、磷含量變化及氮、磷交互作用對金藻增殖的影響,并比較了不同形態的氮鹽對該藻增殖的效應。試驗得出該藻生長的氨氮(氯化銨)最適濃度水平為150 μmol-N/L,相應的無機磷的最適濃度水平為10 μmol-P/L;有機氮(尿素)的最適濃度水平為250 μmol-N/L,相應的無機磷最適濃度水平為5 μmol-P/L。本研究通過對管角螺糞便檢測,N/P接近28,水體中P的濃度相對N來說較低,用管角螺代謝物培養亞心形扁藻,效果沒有湛江等鞭金藻、牟氏角毛藻明顯,側面可以證明扁藻對營養物質的需求量遠大于金藻、牟氏角毛藻,與上述學者的研究結論相吻合。一般來說,在自然海水及水生生物糞便中,磷的含量相對低于氮的含量,該含量無法維持亞心形扁藻較長時間生長,在剛開始培養階段雖然無明顯差別,待水體中的磷消耗殆盡時生長差異就表現出來。
在自然生態環境中,貝類和浮游植物處于同一個生態環境中,不僅存在濾食性貝類對浮游植物的攝食和促長雙重作用,而且浮游植物的數量變動也直接影響一些貝類的發育[27]。而一些肉食性貝類,不能濾食水中的微小顆粒及浮游生物,其自身的排泄物能對浮游植物的繁殖生長起促進作用。因此,我們在規模化養殖時應注意兩種貝類合理搭配,防止養殖海域富營養化而導致浮游植物大量繁殖,給生產造成損失。
[1]農業部漁業局.中國漁業年鑒[M].北京:中國農業出版社,2014:65-84.
[2]蔣艾青.殘餌、死魚及排泄物腐解對山區精養池養殖水質的影響[J].現代農業科技,2007(10):143-144.
[3]文海翔,張濤,楊紅生,等.溫度對硬殼蛤Mercenaria mercenaria(Linnaeus,1758)呼吸排泄的影響[J].海洋與湖沼,2004,35(6):549-555.
[4]伊紹武,廖經球,黃海,等.東風螺生物學及養殖生態學研究研究進展[J].水產科學,2007,26(11):632-636.
[5]溫成器.海藻與貝類混養的優勢[J].漁業致富指南,2006(12):50.
[6]Drenner RW.Experimental studies of size-selective phytoplankton grazing by a filter-feeding cichlid and the cichild's effects on plankton community structure[J].Limnol Oceanogr,1987(32):1138-1144.
[7]費尊樂,毛興華.渤海生產力研究:葉綠素a初級生產力與漁業資源開發潛力[J].海洋水產研究,1991(12):55-69.
[8]王俊,姜祖輝,董雙林.濾食性貝類對浮游植物群落增殖作用的研究[J].應用生態學報,2001,12(5):765-768.
[9]王俊,姜祖輝,董雙林.櫛孔扇貝對單細胞藻類生長間接作用的研究[J].海洋水產研究,2001(02):42-46.
[10]羅杰,曹伏君,劉楚吾,等.我國管角螺的研究現狀[J].海洋漁業,2014,36(3):282-288.
[11]羅杰,劉楚吾,李鋒,等.管角螺窒息點及晝夜代謝規律[J].應用生態學報,2008,19(9):2092-2096.
[12]羅杰,劉楚吾,李鋒,等.鹽度及規格對管角螺耗氧率和排氨率的影響[J].海洋科學,2008,32(5):46-50.
[13]陳坤,張前前,史海燕,等.浮游植物計數方法比較研究[J].海洋環境科學,2007,26(4):383-385.
[14]成永旭.生物餌料培養[M].北京:中國農業出版社,2007:66-67.
[15]Maestrini SY.Fertilite et leneurser sels nutritifs des eauxmarines[J].J Rech Oceanogr,1979(4):17-25.
[16]柴雪良,張炯明,方軍,等.樂清灣、三門灣主要濾食性養殖貝類碳收支的研究[J].上海海洋大學學報,2006,15(1): 52-58.
[17]吳小平,吳宗文,高啟平,等.碳匯漁業養殖工程裝備及效能研究[J].重慶水產,2012(2):28-33.
[18]李鋒,葛長字,方建光.不同溫度和接種密度下亞心形扁藻增殖的初步研究[J].海洋水產研究,2007,28(6):61-66.
[19]楊東方,陳生濤,胡均.光照、水溫和營養鹽對浮游植物生長重要影響大小的順序[J].海洋環境科學,2007,26(3):201-207.
[20]錢振明,邢榮蓮,湯寧.光照和鹽度對8種底棲硅藻生長及其生理生化成分的影響[J].煙臺大學學報:自然科學與工程版,2008,21(1):46-52.
[21]Sommer U.Nutrient competition between phytoplankt on inmultispecies chemostat experiments[J].Arch Hydrobiol,1983,96(2):399-416.
[22]林忠洲,徐善良,邵波,等.不同氮磷質量濃度下青島大扁藻和牟氏角毛藻的種間競爭關系[J].寧波大學學報:理工版,2013,26(1):12-17.
[23]孫育平,趙日水.不同氮、磷濃度下亞心形扁藻的生長及水中氮、磷變化[J].水產科學,2011,30(4):197-201.
[24]Austin A P,Ridley T C l,Lucey W P.Effects of nutrient enrichment on marine periphyton:Implications for abalone culture[J].Botanic Marina,1990,33(3):235-239.
[25]于瑾,蔣霞敏,梁洪.氮、磷、鐵對牟氏角毛藻生長速率的影響[J].水產科學,2006,25(3):121-124.
[26]張青田,張兆琪,董雙林.氮、磷對金藻的增殖效應[J].海洋湖沼通報,2002,(2):45-51.
[27]Marie-Laure V.Impact des priduct d'excretion azotes des huitressur la fertilite deseaux desclaires ostreicoles[J].J Rech Oceanogh,1986,11(1):33-35.
The effects of Hemifusus tuba metabolites on proliferation of phytoplankton
Luo Jie,Liu Hao,Liu Chuwu,Cao Fujun,Su Dongjie
(Fisheries College,Guangdong Ocean University,Key Laboratory of Aquaculture in South China Sea for Aquatic Economic Animal of Guangdong Higher Education Institutes,Zhanjiang 524025,China)
Collection the dung of Hemifusus tuba,after drying treatment and measure its nutritional ingredients.And under laboratory conditions,the influence of growth to Isochrysis zhanjiangensis、Chaetoceros muelleri and Platymonas subcordiformis of Hemifusus tuba metabolites were studied.The results showed that the waste of Hemifusus tuba contains coarse protein(9.8%),p(0.38%),and other nutrients needed for the growth of microalgae;the metabolites of Hemifusus tuba could obviously promote the growth of Isochrysis zhanjiangensis、Chaetoceros muelleri and Platymonas subcordiformis(P<0.05).Their growth with the adding amount of the metabolites were positively related;and the effect metabolites to growth of different microalgae had obvious differences,according to the different types of Isochrysis zhanjiangensis and Chaetoceros muelleri proliferation effect was significantly higher than the Platymonas subcordiformis(P<0.05).
Hemifusus tuba;metabolites;microalgae;growth
10.3969/j.issn.1004-2091.2016.10.001

Q175
A
1004-2091(2016)10-0001-06
2016-04-29)
廣東省科技計劃項目(2013B020503063)
羅杰(1967-),男,教授級高工,主要從事水產經濟動物人工繁育與增養殖技術研究.E-mail:hydxlj163.com
劉楚吾,教授.E-mail:liucw@gdou.edu.cn
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
