渤海東部海域秋季底層游泳動物種類組成及群落多樣性
2016-11-28 07:23:42任中華魏佳麗呂振波高彥潔叢旭日
生態學報 2016年17期
任中華,李 凡,魏佳麗,3,呂振波,*,高彥潔,叢旭日
1 山東省海洋資源與環境研究院,山東省海洋生態修復重點實驗室,煙臺 264000 2 上海海洋大學,海洋科學學院,上海 201306 3 上海海洋大學,水產與生命學院,上海 201306
?
渤海東部海域秋季底層游泳動物種類組成及群落多樣性
任中華1,2,李 凡1,魏佳麗1,3,呂振波1,*,高彥潔1,2,叢旭日1,2
1 山東省海洋資源與環境研究院,山東省海洋生態修復重點實驗室,煙臺 264000 2 上海海洋大學,海洋科學學院,上海 201306 3 上海海洋大學,水產與生命學院,上海 201306
根據2014年9月在渤海東部海域(37°40′—38°20′N、120°00′—121°20′E)進行的游泳動物底拖網調查數據,應用相對重要性指數、生物多樣性指數、群落結構多元統計分析等方法對該海域游泳動物種類組成及群落多樣性特征進行了分析。結果表明:調查海域共捕獲游泳動物54種,以頭足類槍烏賊(Loliolusspp.)為絕對優勢種;游泳動物資源密度為28.82kg/h和5166尾/h。游泳動物種類Margalef豐富度指數(D)的變化范圍為1.99—3.67,Shannon-Wiener多樣性指數(H′)變化范圍為0.89—2.28,Pielou均勻度指數(J′)變化范圍為0.29—0.69。群落結構多元統計分析表明,渤海東部海域游泳動物群落結構以60%的相似性可劃分為3個組群:長島群島海域組群(A組群)、龍口海域組群(B組群)和蓬萊海域組群(C組群);ANOSIM分析表明,組群之間差異極顯著,兩兩之間差異亦極顯著。受增殖放流影響,C組群資源密度和種類多樣性均保持較高的水平,而A、B組群人為活動頻繁,過度捕撈嚴重,對海域生態環境和漁業資源整體結構的破壞較大。
渤海東部海域;游泳動物;種類組成;優勢種;多樣性;群落結構
游泳動物洄游特性分為3種,長距離洄游種、短距離洄游種和地方性種類,渤海東部海域作為短距離洄游種和地方性種類的主要洄游路線具有重要意義。該海域處于黃渤海的鏈接地帶,通有渤海海峽,自南向北水深逐漸加深,且平均水深高于渤海平均水深18 m[1];溫度適宜,餌料充足,許多短距離洄游種類經黃海進入石島、煙威沿岸行生殖洄游。幼體成長后向渤海海峽北部深水區移動,與成體混群棲息。可以看出,渤海東部海域以其得天獨厚的自然條件為短距離洄游種類和地方性種類的繁殖、索餌、生長提供了便利,相較于渤海其他海域,該海域的生物多樣性和資源量分布更能保持一個較高的水平。
20世紀80年代以來,由于近海主要經濟魚類的過度捕撈以及頻繁的海洋開發活動,近海魚類資源已經嚴重衰退,許多游泳動物種類已瀕臨滅絕。據調查,龍口附近海域的填海造陸活動對其附近海域的生態系統造成極大破壞,長島群島海域的旅游發展對附近海域的環境污染尤為嚴重,渤海東部海域的近岸水產養殖活動和過度捕撈壓力造成該海域游泳動物趨向小型化發展。部分種類如許氏平鲉(Sebastesschlegeli)、中國明對蝦(Fenneropenaeuschinensis)等只能靠靠增殖放流來維持種群數量[2]。
近年來,針對渤海游泳動物群落結構、多樣性及群落聚類等方面的研究文章較多,但主要集中在除渤海東部海域以外的其他渤海海域[3- 11],渤海東部海域游泳動物相關研究未見報道,無可參考對比。本文主要根據2014年9月在渤海東部海域進行的底拖網調查數據,利用生態優勢度、群落多樣性、多元統計分析等方法分析了該海域秋季游泳動物群落結構及多樣性變化特征,主要了解該海域秋季游泳動物的群落結構特征和種類多樣性,為后續研究該海域的群落結構年際變化季節變化作出指引,同時為渤海東部海域漁業資源的合理開發及保護提供科學建議。
1 材料與方法
1.1 數據來源
調查區域位于渤海東部37°40′—38°20′N、120°00′—121°20′E海域范圍內,大致分為龍口海域、蓬萊海域和長島群島海域3個海域,于2014 年9月在該海域進行1個航次底拖網調查。調查海域設置21個調查站位(表1)。調查船只功率260 kW,調查網具為單船底拖網,網口周長30.6 m,網囊網目20 mm,拖曳時網口寬度約8 m。每站拖曳1 h,拖速3 kn。漁獲物樣品參照《海洋調查規范(GB/T 12763.6—2007)[12]進行生物學測定和分析處理。游泳動物種類名稱及分類地位以《中國海洋生物名錄》[13]為依據。數據進行分析之前,將每個站位的漁獲量統一標準化為生物量資源密度和尾數資源密度(kg/h, 尾/h)。
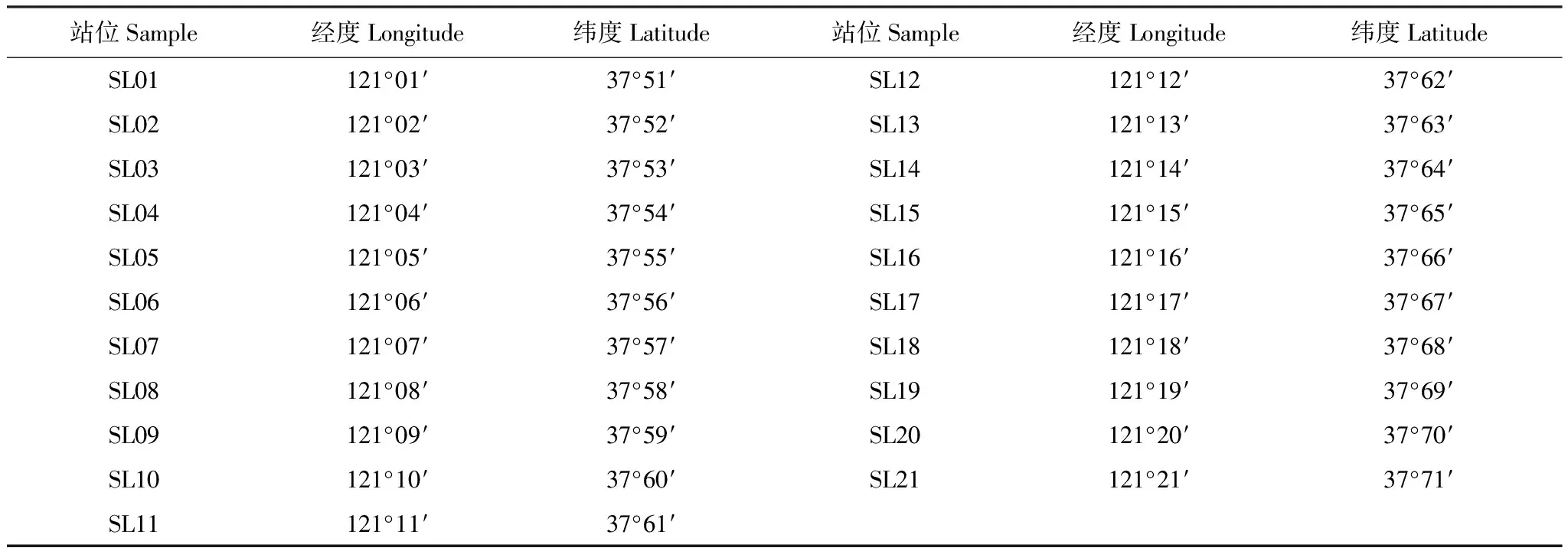
表1 渤海東部海域游泳動物調查站位表
1.2 分析方法
1.2.1 相對重要性指數
采用PINKAS等[14]相對重要性指數(IRI, index of relative importance)確定種類在群落中的重要性,計算公式為:
IRI=(N+W)×F
式中,N為某一種類的尾數資源密度占總尾數資源密度的百分比;W為某一種類的生物量資源密度占總生物量資源密度的百分比;F為某一種類出現的站位數占調查總站位數的百分比。將IRI值大于等于1000的種類定義為優勢種。
1.2.2 群落多樣性
采用Margalrf種類豐富度指數(D)[15]、Shannon-Wiener多樣性指數(H′)[16]和Pielou均勻度指數(J′)[17]分析游泳動物群落結構多樣性。
Margalrf種類豐富度指數:
D=(S-1)lnN
Shannon-Wiener多樣性指數:
Pielou均勻度指數:
J′=H′/lnS
式中,S為種類數,N為總尾數資源密度,Pi為第i種生物量資源密度占總生物量資源密度的比例,多樣性的計算采用PRIMER 6軟件完成。
1.2.3 群落結構的多元統計分析
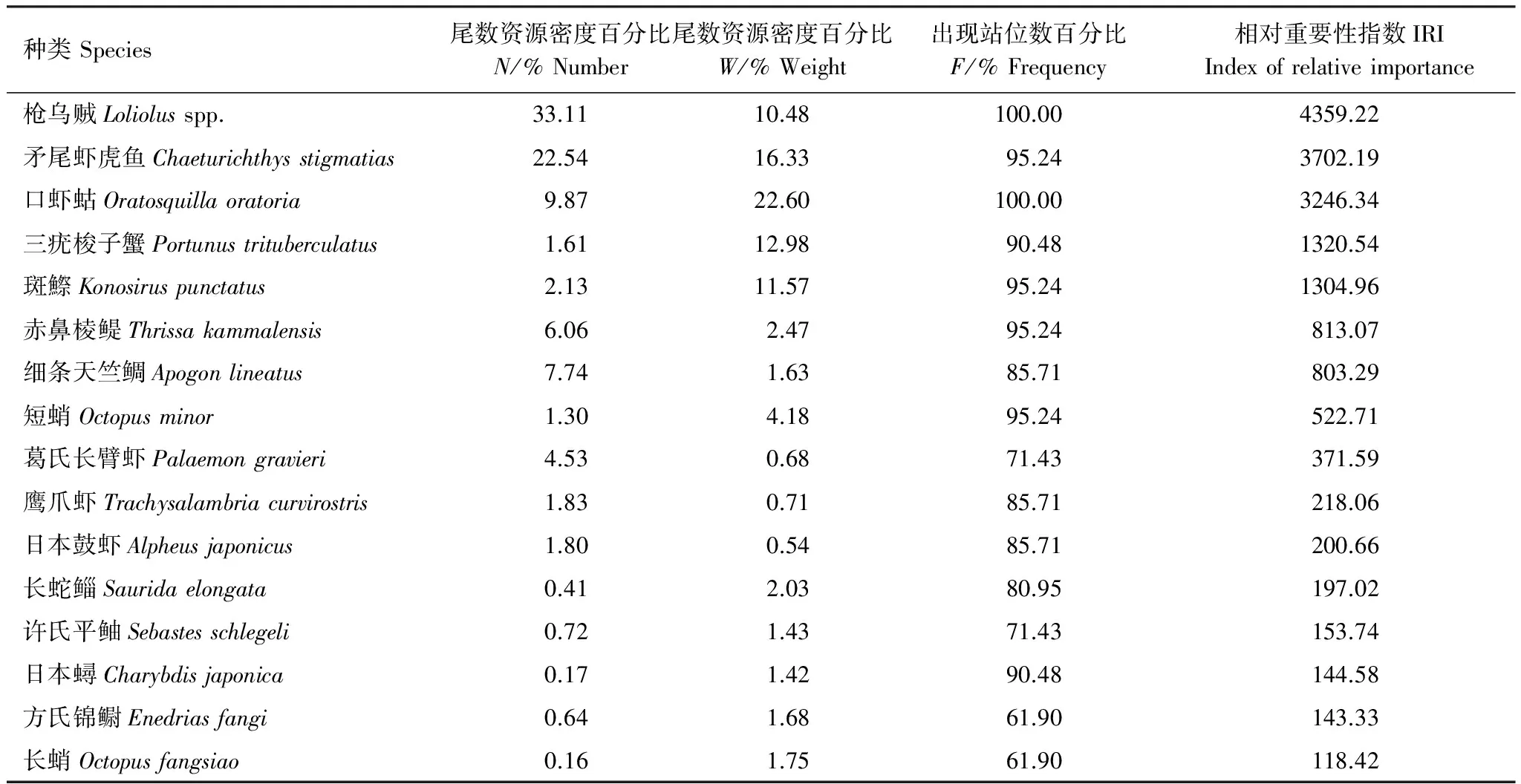
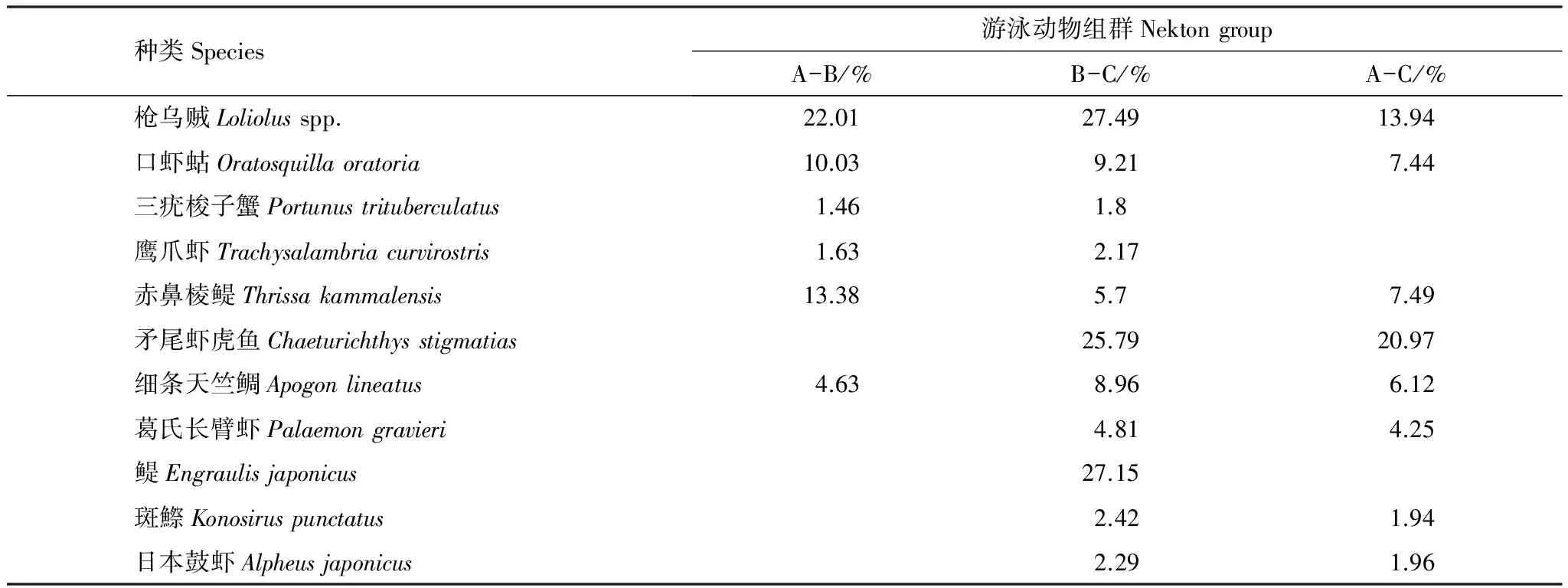
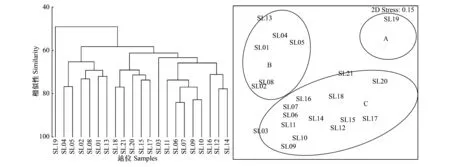
對渤海東部漁獲游泳動物的生物量資源密度進行四次方根轉換,并計算Bray-Curtis 相似性系數矩陣,采用非度量多維標度分析(MDS)與等級聚類分析(CLUSTER)來研究游泳動物群落結構特征。同時用脅迫系數(Stress)來衡量MDS二維點圖的優劣,一般認為:當0.1 2.1 種類組成 調查海域共捕獲魚類34種,隸屬于23科33屬,其中鱸形目種類最多,共16種,占魚類調查總種數的47.06%;其次為鯡形目,共6種,占魚類調查總種數的17.65%。甲殼類17種,隸屬于13科16屬,其中口足目16種,占甲殼類調查總種數的94.12%,口足目一種,占5.88%。頭足類3種,隸屬于2科2屬,分別為短蛸,槍烏賊和長蛸(附表1)。 調查研究表明,萊州灣游泳動物共155種,其中魚類115種,甲殼類40種,萊州灣和渤海魚類種類基本一致[20- 21],可以看出,渤海東部海域游泳動物相對萊州灣海域種類數減少。歷史資料中,張春霖等報道渤海魚類116種[22];田明誠等調查記錄渤海魚類156種[23],朱鑫華等根據渤海調查記錄魚類119種[24]。 2.2 優勢種 本次調查優勢種為5種,以頭足類槍烏賊(Loliolusspp.)為絕對優勢種,重要種有11種。表2可以看出,斑鰶(Konosiruspunctatus)、赤鼻棱鳀(Thrissakammalensis)為渤海東部海域主要中上層魚類;矛尾蝦虎魚(Chaeturichthysstigmatias)為主要的底層魚類;口蝦蛄(Oratosquillaoratoria)、三疣梭子蟹(Portunustrituberculatus)為主要的甲殼類。16種主要種類的重量累積百分比為92.49%,尾數累積百分比為94.63%,在調查海域漁業資源分布中占有很大比例,存在絕對優勢。槍烏賊、斑鰶、赤鼻棱鳀和矛尾蝦虎魚等占比重最大的種類均為經濟價值低、個體小的種類,鄧景耀等指出20世紀初渤海海域游泳動物群落結構已趨向小型化、低質化發展[6]。 2.3 資源量及空間分布 調查海域游泳動物平均資源密度分別為28.82kg/h和5166尾/h,其中魚類平均生物量資源密度和甲殼類相仿,分別為12.59kg/h、 11.51kg/h,頭足類為最低,為4.73kg/h。如表3所示,生物量資源密度以SL12、SL14、SL15、SL16、SL17站位捕獲量最大,集中于長島群島海域,優良的水質條件和索餌水域為游泳動物的生長提供了便利的條件;而尾數資源密度以SL03、SL06、SL07、SL09、SL12、SL15、SL17站位最大,這些站位主要分在在龍口海域,該區域漁獲游泳動物以赤鼻棱鳀(Thrissakammalensis)、鳀(Engraulisjaponicus)和槍烏賊(Loliolusspp.)占比重最大,3種優勢種類均為個體小、營養級低的小型中上層魚類和無脊椎動物,資源開發的主要是當年生幼魚[25],造成該海域生物量資源密度低而尾數資源密度較高。 表2 渤海東部海域群落優勢種組成 表3 渤海游泳動物平均生物量、尾數資源密度分布 2.4 生物多樣性 調查海域生物種類多樣性指數平均為1.70,變化范圍為0.89—2.28,最高值出現在SL18站位,最低值出現在SL13站位;物種均勻度指數平均為0.53,變化范圍0.29—0.69,最高值出現在SL18站位,最低值出現在SL13站位;物種豐富度指數平均為2.85,變化范圍1.99—3.67,最高值出現在SL17站位,最低值出現在SL01站位。渤海東部海域處于黃渤海連接地帶,水質優良、餌料充足、棲息地保存較為完好,游泳動物物種多樣性得以保持較高的水平;除了沿岸的養殖活動外,該海域捕撈強度相對較少,各站位均勻度較為一致;由于捕撈季節為秋冬,非游泳動物產卵繁殖時間,加之洄游等因素,因而各站位豐富度指數較小。 2.5 群落結構多元統計分析 根據PRIMER6多元統計分析,聚類結果及空間分布效果明顯。圖1為渤海東部海域游泳動物群落結構CLUSTER聚類分析和MDS分析的結果,根據MDS分析的脅迫系數(stress=0.15),MDS排序結果可用二維點陣圖表示。 游泳動物群落以60%的相似性劃分為3組,分別為A組群(SL19號站)、B組群(SL01—SL02,SL04—SL05,SL08和SL13號站)及C組群(SL03,SL06—SL07,SL09—SL12,SL14—SL18和SL20—SL21號站)。A組群位于長島群島的單個站位,多樣性指數均處于較低的水平;C組群廣泛分布于蓬萊海域和長島群島海域;B組群集中分布于龍口海域,3個群落由近海近岸海域向東北方向分布。ANOSIM分析表明,3個組群之間差異極顯著(P<0.01),兩兩之間差異亦極顯著(P<0.01),說明分組結果較好。本次調查無海域水質和底質調查,因而通過種類組成和群落結構作出初步分組,為后續游泳動物群落結構與環境因子相結合分組研究作出輔助。 SIMPER分析表明,本次調查聚類分析結果中,對組內相似性貢獻較大的典型種如表4所示,對A組群相似性貢獻較大的種類為槍烏賊,累積相似性貢獻率為97.45%;對B組群相似性貢獻率較大的種類為槍烏賊、口蝦蛄、三疣梭子蟹、鷹爪蝦、赤鼻棱鳀,累積相似性貢獻率為90.50%;對C組群相似性貢獻率較大的種類為槍烏賊、口蝦蛄、赤鼻棱鳀、矛尾蝦虎魚、細條天竺鯛、葛氏長臂蝦,累積相似性貢獻率為91.46%。對各兩組群間的組間相異性貢獻百分比較大的分歧種如表5所示,其中對A和B組群、A和C組群以及B和C組群相異性貢獻均較大的種類有槍烏賊、口蝦蛄、赤鼻棱鳀和細條天竺鯛,為小型化、低質化種類,經濟價值低,一定程度上反映出該海域游泳動物種類變化趨勢。 表4 各組群典型種及其對組內相似性貢獻百分比 表5 各兩組群分歧種及其對組間差異性貢獻百分比 A-B表示比較A組群和B組群之間共有生物種的對該組群的差異性貢獻百分比;B-C表示比較B組群和C組群之間共有生物種的對該組群的差異性貢獻百分比; A-C表示比較A組群和C組群之間共有生物種的對該組群的差異性貢獻百分比. 圖1 渤海東部游泳動物CLUSTER與MDS聚類分析結果Fig.1 Results of CLUSTER analysis and MDS analysis of nekton community 3.1 渤海東部海域游泳動物種類組成分析 渤海東部海域位處黃渤海的鏈接地帶,平均水深高于渤海平均水深18m[1],資源豐富,海水理化性質穩定,適宜海洋生物的生長繁殖[26],因其得天獨厚的優越條件,長期以海洋漁業和水產養殖作為該地區的主要經濟來源,并伴隨海島旅游業的不斷發展。 近年來隨著環渤海地區社會經濟的不斷發展和人類對海洋資源的開發和利用,使得海洋生態環境受到一定程度的干擾和破壞。從國家海洋局海洋環境質量公報中可知,嚴重污染海域主要集中在遼東灣近岸、渤海灣、黃河口和萊州灣,這都與渤海的地形與環流特征密切相關[1]。渤海主要海水運動形式為潮汐運動,環流主要表現為潮余流[27- 28],并且潮余流在渤海是形成污染物擴散的主要應力之一。渤海東部海域連接黃渤海,通有渤海海峽,潮流運動合理,海域水交換能力的強,能充分利用自凈能力,能最大化地減少污染對渤海生態系統的負面影響。 自20世紀50年代以來受人類活動(包括過度捕撈、水利建設、陸源污染、海洋海岸工程、水產養殖等)以及氣候變化的影響,渤海東部海域游泳動物群落受到了嚴重干擾,游泳動物種數特別是魚類種數總體呈下降趨勢。蓬萊海域每年的增殖放流活動在一定程度上能補充游泳動物資源量,較之于萊州灣目前放流種類[20],渤海東部海域放流種類較少,在本調查中僅漁獲到許氏平鲉、黑棘鯛、三疣梭子蟹和中國明對蝦4種,其他增殖放流種類沒有捕獲到可能與調查站位布設有關。 3.2 優勢種 目前渤海東部海域游泳動物群落主要種類主要為個體小、營養級低、經濟價值低的種類(表6),優勢種類的營養級以3級(食物鏈中的次級消費者)為主。研究指出,1959年萊州灣優勢種為帶魚(Trichiurusjaponicus)、小黃魚(Larimichthyspolyactis)、半滑舌鰨(Cynoglossussemilaevis)、對蝦(Penaeuschinensis)和白姑魚(Argyrosomusargentatus)等5 種大型底層魚蝦類[6];1982 、1992—1993 年和1998 年進行的3 次調查結果表明, 上述優勢種群逐步為黃鯽(Setipinnataty)、鳀(Engraulisjaponicus)、赤鼻棱鳀(Scutengrauliskammalensis)、斑鰶(Clupanodonpunctatus)、槍烏賊(Loligospp.)、青鱗小沙丁魚(Harengulazunasi)等小型中上層魚類所代替,優勢種向小型化和低質化發展,游泳動物群落結構變化;1982年以來,萊州灣優勢種變化不大,但漁獲的資源量卻逐年下降。相較于萊州灣,其毗鄰海域渤海東部海域的優勢種發展趨勢相同:底質化和小型化,且漁獲資源量較低。 表6 渤海東部海域游泳動物主要種類特征 I:小型Small sized;II中小型Small and medium-sized;III:中型Medium sized;IV:大中型Large and medium-sized;V:大型Large sized;A:優質類Very high;B:經濟類High;C:次經濟類Medium;D:低質類Low; *數據引自唐啟升和葉懋中[29];**數據引自Fishbase數據庫Froese & Pauly和楊紀明[30- 31]的文獻 孫中之等[32]研究指出:20世紀50年代末至今游泳動物群落優勢種沿帶魚(Trichiurusjaponicus)→黃鯽(Setipinnataty)→鳀→赤鼻棱鳀→槍烏賊方向,不斷朝向更小型化的方向演替,與其對比,本次漁獲游泳動物優勢種類鳀、赤鼻棱鳀和槍烏賊等與小型化趨勢相仿,個體較小、營養價值和經濟價值均較低。 3.3 游泳動物群落多樣性分析 龍口海域尾數資源密度較高,生物量資源密度較小,這可能與龍口市填海造地的活動有關,同時,龍口上方海域的養殖活動對N、P、K等水質環境因子影響較大,可能會對游泳動物個體的生長造成影響[33]。渤海東部海域游泳動物多樣性指數整體較高,均與度指數次之,豐富度指數整體較低。SL01、SL03、SL04和SL13站位位于龍口上方海域養殖區內,指數偏低可能與該區域N、P污染較重,富營養化程度較高有關[25,30];SL19站位于長島群島風景區,受人為擾動程度較大。SL10站位出現的細條天竺鯛漁獲資源量高達648尾/h,其次為矛尾蝦虎魚、葛氏長臂蝦、日本鼓蝦,漁獲資源量分別為372尾/h、360尾/h和324尾/h;SL17站位矛尾蝦虎魚漁獲資源量高達3432尾/h,赤鼻棱鳀、槍烏賊和細條天竺鯛次之,分別為1608尾/h、840尾/h和720尾/h,兩個站位主要漁獲種類的個體較小,但尾數資源密度較高,豐富度指數相對較高。除SL01、SL03、SL13和SL19站位外,其余展位的多樣性指數均處于較高水平。SL03和SL13站位位于養殖區域,養殖活動產生的污染造成附近海域水質狀況不佳,游泳動物群落生長、繁殖受到阻礙,生物多樣性顯著降低。目前針對渤海東部海域游泳動物群落結構及其多樣性的研究較少,筆者建議調查海域管理部門應根據當地的經濟社會發展合理規劃海域使用,減少人為發展、建設活動中對海域生態環境的破壞,加強對受污染海域的治理,,合理利用漁業資源,使海域生態環境、漁業資源可持續發展。 3.4 游泳動物群落結構空間格局 游泳動物群落以60%的相似性劃分為3組,分別為A組群(SL19號站)、B組群(SL01—SL02,SL04—SL05,SL08和SL13號站)及C組群(SL03,SL06—SL07,SL09—SL12,SL14—SL18和SL20—SL21號站)。A組群包含單個站位,SL19站位位于長島群島風景區,旅游開發的海釣、海上娛樂、觀賞等活動造成該海域生態環境破壞,改變游泳動物的群落結構,使該海域游泳動物的生物多樣性減小,SL19站位游泳動物種類數為18種,均為小型種類,鳀、槍烏賊和赤鼻棱鳀漁獲資源量最高,分別為1824尾/h、1182尾/h和720尾/h,其余種類漁獲資源量均小于100尾/h,該站位漁獲種類75%屬于中上層種類,垂直分布不均勻,這可能與底層生態環境遭到破壞有關。C組群主要集中于龍口上方海域,該海域調查站位漁漁獲游泳動物種類數較少,豐富度指數很低,槍烏賊占主導地位,與口蝦蛄、三疣梭子蟹、鷹爪蝦和赤鼻棱鳀的相似性的貢獻率達到90.50%。 C組群位于蓬萊上方海域,漁獲種類數多,矛尾蝦虎魚和槍烏賊占主導地位,垂直分布均勻,同時,該海域水深相對較深,溫度相對較高,適合大部分游泳動物的冬季越冬。本次調查發現的4種增殖放流種類均集中于C組群調查站位,可能受增殖放流的影響,該海域游泳動物種類多樣性保持較高的水平,漁獲資源量相對較高。 致謝:感謝山東省海洋資源與環境研究院環境監測中心何鑫、蘇博以及魯昌漁64193船長陳奎賢和船員在出海調查取樣過程中給予的幫助。 [1] 李裴. 渤海海域潮流及潮余流的數值模擬[D]. 天津: 天津大學, 2009. [2] 逢志偉, 徐賓鐸, 陳學剛, 任一平. 膠州灣中部海域蝦類群落結構及其多樣性. 中國水產科學, 2013, 20(2): 361- 371. [3] 高音, 劉明勇, 湯勇, 李九奇, 邢彬彬. 遼東灣漁業資源及生態環境的調查分析. 大連海洋大學學報, 2013, 28(2): 211- 216. [4] 吳忠鑫, 張磊, 張秀梅, 張沛東, 李文濤. 榮成俚島人工魚礁區游泳動物群落特征及其與主要環境因子的關系. 生態學報, 2012, 32(21): 6737- 6746. [5] 金顯仕, 鄧景耀. 萊州灣漁業資源群落結構和生物多樣性的變化. 生物多樣性, 2000, 8(1): 65- 72. [6] 鄧景耀, 金顯仕. 萊州灣及黃河口水域漁業生物多樣性及其保護研究. 動物學研究, 2000, 21(1): 76- 82. [7] 李凡, 呂振波, 魏振華, 王田田, 徐炳慶, 王忠全. 2010年萊州灣底層漁業生物群落結構及其季節變化. 中國水產科學, 2014, 20(1): 137- 147. [8] 孫鵬飛, 單秀娟, 吳強, 陳云龍, 金顯仕. 萊州灣及黃河口水域魚類群落結構的季節變化. 生態學報, 2014, 34(2): 367- 376. [9] 鄭亮, 呂振波, 李凡, 張嵐, 于文宋. 調水調沙期間黃河口海域魚類群落結構特征. 中國水產科學, 2014, 21(3): 602- 610. [10] 張波, 吳強, 金顯仕. 萊州灣魚類群落的營養結構及其變化. 漁業科學進展, 2013, 34(2): 2- 9. [11] 吳強, 王俊, 李忠義, 陳瑞盛, 孫堅強, 金顯仕. 黃渤海春季甲殼類群落結構的空間變化. 水產學報, 2012, 36(11): 1686- 1693. [12] 國家質檢總局, 國家標準化委員會. 海洋調查規范第6部分: 海洋生物調查. 北京: 中國標準出版社, 2007: 56- 62. [13] 劉瑞玉. 中國海洋生物名錄. 北京: 科學出版社, 2008: 903- 1066. [14] Pinkas L, Oliphant M S, Iverson I L K. Food habits of albacore, bluefin tuna, and bonito in California waters. Fish Bulletin, 1971, 152: 1- 105. [15] Margalef R. Information theory in ecology. General System, 1958, 25(1): 393- 399. [16] Wilhm J L. Use of biomass units in Shannon′s formula. Ecology, 1968, 49(1): 153- 156. [17] Pielou E C. Ecological Diversity. New York: Wiley, 1975. [18] Clarke K R, Ainsworth M. A method of linking multivariate community structure to environmental variables. Marine Ecology-Progress Series, 1993, 92: 205- 219. [19] Clarke K R, Warwick R M. Changes in Marine Communities: An Approach to Statistical Analysis and Interpretation. 2nd ed. Plymouth: PRIMER-E Ltd., 2001. [20] 李凡, 張煥軍, 呂振波, 徐炳慶, 鄭亮. 萊州灣游泳動物群落種類組成及多樣性. 生物多樣性, 2013, 21(5): 537- 546. [21] Chen D G, Shen W Q, Liu Q, Jiao Y, Zeng X Q, Ren Y P. The geographical characteristics and fish species diversity in the Laizhou Bay and Yellow River estuary. Journal of Fishery Sciences of China, 2000, 7(3): 46- 52. [22] 張春霖, 成慶泰, 鄭葆珊. 黃渤海魚類調查報告. 北京: 科學出版社, 1955. [23] 田明誠, 孫寶齡, 楊紀明. 渤海魚類區系分析. 海洋科學集刊, 1993, (34): 157- 167. [24] 朱鑫華, 吳鶴洲, 徐鳳山, 葉懋中, 趙紫晶. 黃渤海沿岸水域游泳動物群落多樣性及其相關因素的研究. 海洋學報, 1994, 16(3): 102- 112. [25] 山東省海洋與漁業廳. 山東近海經濟生物資源調查與評價. 北京: 海洋出版社, 2010. [26] 趙瑞. 長島海域大型底棲動物生態學研究[D]. 青島: 中國海洋大學, 2014. [27] 王輝, 蘇志清, 馮士笮, 孫文心. 渤海三維風生-熱鹽-潮致 Lagrange 余流數值計算. 海洋學報, 1993, 15(1): 9- 21. [28] 王強. 渤海環流的季節變化及浮游生態動力學模擬[D]. 青島: 中國海洋大學, 2004. [29] 唐啟升, 葉懋中. 山東近海漁業資源開發與保護. 北京: 農業出版社, 1990. [30] Froese R, Pauly D. FishBase. [2015-03-01]. http://www.fishbase.org. [31] 楊紀明. 渤海無脊椎動物的食性和營養級研究. 現代漁業信息, 2001, 16(9): 8- 16. [32] 孫中之, 周軍, 王俊, 趙振良, 莊申, 陳孛, 閻偉, 許玉甫, 王成華, 高文斌, 孟維東. 黃渤海區拖網漁具綜合定性調查及特點分析. 漁業科學進展, 2011, 32(5): 126- 134. [33] 任中華, 鄭亮, 李凡, 呂振波. 萊州灣海域蝦類群落結構及其多樣性. 海洋漁業, 2014, 36(3): 193- 201. Autumnal species composition and community diversity of nekton in the eastern Bohai Sea REN Zhonghua1,2, LI Fan1, WEI Jiali1,3, Lü Zhenbo1,*, GAO Yanjie1,2, CONG Xuri1,2 1 Shandong Marine Resource and Environment Research Institute, Shandong Provincial Key Laboratory of Restoration for Marine Ecology, Yantai 264000, China 2ShanghaiOceanUniversity,InstituteofMarineScience,Shanghai201306,China3ShanghaiOceanUniversity,InstituteofFisheriesandLifeScience,Shanghai201306,China The species composition and community structural diversity of nekton was examined in the autumn at 21 stations in the eastern Bohai Sea (37°40′—38°20′N, 120°00′—121°20′E) using bottom trawl surveys. Analyses were performed using an index of relative importance, ecological diversity indices, multivariate analysis MDS, and CLUSTER using PRIMER 6 software and other methods. We identified 54 nekton species: 34 fishes, 17 crustaceans and 3 cephalopods, belonging to 23 families and 33 genera. There were 5 dominant species, withLoliolusspp. being the most dominant. There were also 11 relatively dominant species identified, includingKonosiruspunctatus,Thrissakammalensis,Chaeturichthysstigmatias,Oratosquillaoratoria, andPortunustrituberculatus. The relative nekton resources were 28.82 kg/kg2and 5166 ind./h, which had a variable spatial distribution. Much of the resource was centralized, with peak densities congregated around the Long Island Archipelago. Mantissa resources were mainly concentrated around the Long Island Archipelago and in the upper waters of Longkou. Stations SL12, SL14, SL15, SL16, and SL17 had the highest biomass, while stations SL03, SL06, SL07, SL09, SL12, SL15, and SL17 had the highest mantissa. Margalef′s species richness index (D), Shannon-Wiener diversity index (H′), and Pielou′s evenness index (J′) of the community structure ranged from 1.99 to 3.67, 0.89 to 2.28, and 029 to 0.69, respectively. The MDS had a stress level of 0.15, which showed that the community structure of nekton in the eastern Bohai Sea Bay can be divided into three groups: the Long Island Archipelago group (Group A), the Longkou group (Group B), and the Penglai group (Group C). Group A included a single station (SL19), group B included stations SL01, SL02,SL04, SL05, SL08, and SL13, and group C included SL03, SL06, SL07, SL09, SL10, SL11, SL12, SL14, SL15, SL16, SL17, SL18, SL20, and SL21. ANOSIM analysis showed that in the winter, species compositions in the three groups were very significantly different(P<0.01, with very significant differences between any two groups (P<0.01). The typical within-group species and discriminating species between groups had a significant influence on the community structure. The typical within-group species of Group A wasLoliolusspp., with a 97.45% cumulative contribution similarity rate. For group C,Loliolusspp.,Oratosquillaoratoria,Thrissakammalensis,Chaeturichthysstigmatias,Apogonlineatus, andPalaemongraviericombined to create a cumulative contribution similarity rate of 91.46%. For Group B,Loliolusspp.,Oratosquillaoratoria,Portunustrituberculatus,TrachysalambriacurvirostrisandThrissakammalensiscomprised the cumulative contribution similarity rate of 90.50%. The species discriminating between groups A and C wereLoliolusspp.,Oratosquillaoratoria,Thrissakammalensis,Chaeturichthysstigmatias,Apogonlineatus,Palaemongravieri,Engraulisjaponicas,KonosiruspunctatusandAlpheusjaponicas, whose dissimilarity cumulative contribution rate was 91.26%. For Groups A and B, the dissimilarity was due toLoliolusspp.,Oratosquillaoratoria,Portunustrituberculatus,Trachysalambriacurvirostris,Thrissakammalensis, andApogonlineatus, with a cumulative dissimilarity contribution rate of 53.14%. For Groups B and C, the species contributing to the dissimilarity wereLoliolusspp.,Oratosquillaoratoria,Portunustrituberculatus,Trachysalambriacurvirostris,Thrissakammalensis,Chaeturichthysstigmatias,Apogonlineatus,Palaemongravieri,KonosiruspunctatusandAlpheusjaponicus, with a cumulative dissimilarity contribution rate of 90.64%. The division between groups was relatively stable, as the assemblages conformed to the water environments in the different groups. The resource density and species diversity of Group C maintained a high level, due to restocking, whereas the marine ecological environment of Group A and B was damaged by frequent human activities, causing a reduction in nekton size, and leading to low diversity indices and resource density. This study aims to provide a scientific basis for the rational development and protection of nekton resources in the eastern Bohai Sea. eastern Bohai Sea; nekton community; composition; relative importance index; diversity; multivariate analysis 海洋公益性行業科研專項經費項目(200905019);泰山學者崗位經費資助項目(水生動物營養與飼養) 2015- 03- 04; 日期:2016- 02- 02 10.5846/stxb201503040417 *通訊作者Corresponding author.E-mail: ytlvzhenbo@163.com 任中華,李凡,魏佳麗,呂振波,高彥潔,叢旭日.渤海東部海域秋季底層游泳動物種類組成及群落多樣性.生態學報,2016,36(17):5537- 5547. Ren Z H, Li F, Wei J L, Lü Z B, Gao Y J, Cong X R.Autumnal species composition and community diversity of nekton in the eastern Bohai Sea.Acta Ecologica Sinica,2016,36(17):5537- 5547.2 結果與分析


3 討論