泰山螭霖魚線粒體16S rRNA基因片段序列及分子系統發育分析
2016-12-07 05:15:17楊玲鞏俊霞李嫻張龍崗付佩勝張金路
長江大學學報(自科版) 2016年33期
楊玲,鞏俊霞,李嫻 張龍崗,付佩勝,張金路
山東省淡水水產遺傳育種重點實驗室,農業部黃河下游漁業資源環境科學觀測實驗站,山東省淡水漁業研究院,山東濟南250017
?
泰山螭霖魚線粒體16S rRNA基因片段序列及分子系統發育分析
楊玲,鞏俊霞,李嫻 張龍崗,付佩勝,張金路
山東省淡水水產遺傳育種重點實驗室,農業部黃河下游漁業資源環境科學觀測實驗站,山東省淡水漁業研究院,山東濟南250017
以泰山螭霖魚(Varicorhinusmacrolepis)30個個體為材料,對其線粒體16S rRNA基因片段進行了PCR擴增和序列對比,檢測了泰山螭霖魚的遺傳多樣性;結合從GenBank中下載的同源性較高的鯉科7屬部分魚類的同源序列,利用MGEA軟件計算遺傳距離,采用聚類分析方法,構建了NJ和UPGMA系統進化樹。結果顯示:30個體泰山螭霖魚線粒體16S rRNA基因片段長度為468bp,無堿基的插入和缺失,共檢測到1個多態位點,存在2種單倍型,其單倍型多樣性(Hd)為0.067,核苷酸多樣性(Pi)為0.00015,平均核苷酸差異數(K)0.067。遺傳距離和NJ、UPGMA系統進化樹表明,泰山螭霖魚與多鱗白甲魚的16SrRNA序列一致,泰山螭霖魚所在的突吻魚屬與光唇魚屬的親緣關系最近,與扁吻魚屬遺傳距離最遠。
泰山螭霖魚(Varicorhinusmacrolepis);16S rRNA;遺傳多樣性
泰山螭霖魚(Varicorhinusmacrolepis)屬鯉科鲃亞科突吻魚屬,是生活在泰山海拔270~800m山澗溪流中的小型野生魚類,其肉嫩味美、營養豐富,有獨特的藥用保健價值[1],是泰山特有的珍貴魚類,為我國“五大貢魚”之一[2]。自20世紀90年代起,魚類學者們對生態習性和生物學特點[2]、人工馴化和繁殖[3]、解剖學[4]、生長性能[5]、同工酶[6]、染色體核型[7]進行了研究。近年來,利用分子標記技術對泰山螭霖魚的遺傳多樣性[8,9]和部分功能基因進行了研究[10~12],陳紅菊[11]對其線粒體cyt b和D-loop進行過研究,但有關泰山螭霖魚線粒體16S rRNA基因的研究尚未見報道。為此,本研究通過對泰山螭霖魚的16S rRNA基因部分序列的分析,探討其與鯉科7個屬魚類的進化關系,確定泰山螭霖魚的分類地位,以期為其種質資源的管理與保護及遺傳育種提供分子生物學依據。
1 材料與方法
1.1 材料
實驗所用泰山螭霖魚取自泰安市泰山螭霖魚原種場,樣本數量為30個; PCR引物由上海生工生物技術有限公司合成;DNA提取試劑盒購自上海生工;Taq DNA聚合酶、dNTP 、MgCl2、Marker等為大連寶生物工程有限公司生產;PCR儀為Takara TP650型。
1.2 方法1.2.1 基因組DNA的提取
取泰山螭霖魚背部肌肌肉約20mg,用DNA提取試劑盒(上海生工)提取樣品的基因組DNA,經D260/D280比值和瓊脂糖凝膠電泳來檢測所提取DNA的濃度和純度后,置-20℃冰箱備用。
1.2.2 引物設計及PCR擴增
根據GenBank(NC_NC_023799.1)中的相應序列設計引物,用于16S rRNA基因序列的擴增,引物序列16S-F:5’-GAAAATCACCTAACCTCCC-3’,16S-R:5’-GTTGAACAAACGAACCCTT-3’。
PCR擴增體系50μL,擴增條件:94℃預變性5min;之后進行35個循環(94℃ 40s,55℃ 50s,72℃ 50s);最后72℃延伸10min。PCR擴增產物經1 %的瓊脂糖凝膠電泳(0.5×TBE 5V/cm恒壓)檢測,EB染色,凝膠成像系統觀察拍照,送上海生工生物技術有限公司,純化后采用上述引物進行雙向測序。
1.2.3 序列分析
測序結果經校對,與從GenBank中下載的同源性比較高的鯉科7屬部分魚類的同源序列用Clustal W方法進行比對分析,用MEGA6.06軟件統計序列的堿基組成和變異位點,采用Kimura 2-parameter模型計算遺傳距離,分別用鄰接法(Neighbor-Joining,NJ)和UPGMA方法進行聚類分析,并采用bootstrap 1000次重復檢驗分子系統樹各分枝置信度;用DNASP V5.10軟件計算單倍型多樣性(Hd)、核苷酸多樣性(Pi)和平均核苷酸差異數(K)。
2 結果與分析
2.1 PCR擴增結果
采用引物16S-F和16S-R對30尾泰山螭霖魚的基因組DNA進行PCR擴增,結果都擴增出1條約500bp的條帶(圖1)。
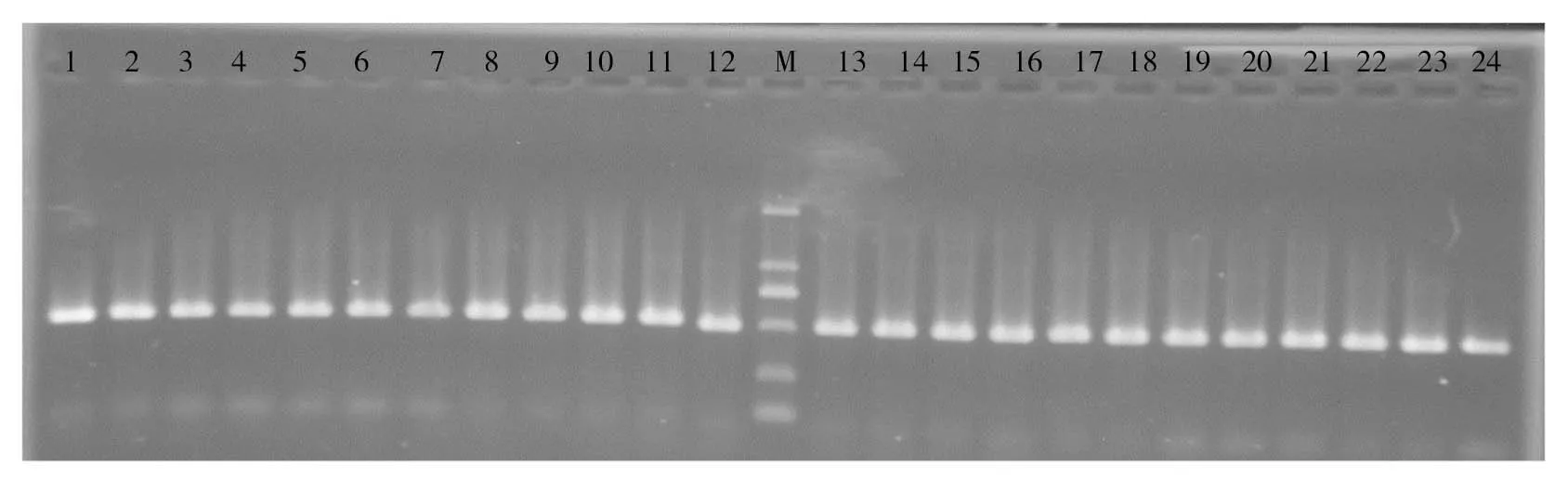
1~24:泰山螭霖魚個體,M:DL-2000 Marker圖1 泰山螭霖魚16S rRNA基因PCR產物電泳圖
2.2 16S rRNA基因片段的序列分析2.2.1 序列分析
泰山螭霖魚30個樣本線粒體16S rRNA基因的測序結果經校對,去除部分端序列后,獲得468bp的片段。采用MEGA6.06軟件經BLAST分析,該片段與GenBank中多鱗白甲魚(Onychostomamacrolepis)的16S rRNA基因片段同源性高達100%,確定該產物為泰山螭霖魚16S rRNA基因的部分序列。
用MGEA 6.06 軟件對測得序列進行比對分析,在測定的468bp的片段中,無堿基的插入與缺失,其中保守位點467個,變異位點1個,占整個序列的0.2%,變異位點為轉換(C←→G),發生在第3位密碼子上,所編碼的氨基酸沒有發生變化。
用DNASP 5.10 軟件分析30個樣本16S rRNA序列,共檢測到2種單倍型,絕大多數個體為單倍型Hap-1,單倍型Hap-2出現的頻率很低,只有1個樣本。
2.2.2 堿基組成和遺傳距離
泰山螭霖魚堿基含量平均為:T 22.6%、C 23.1%、A 32.7%、G 21.6%,G+C含量(44.7%)低于A+T含量(55.3%)(表1)。基于Kimura-2-parameter計算遺傳距離,30個個體的相對遺傳距離為0~0.22%。
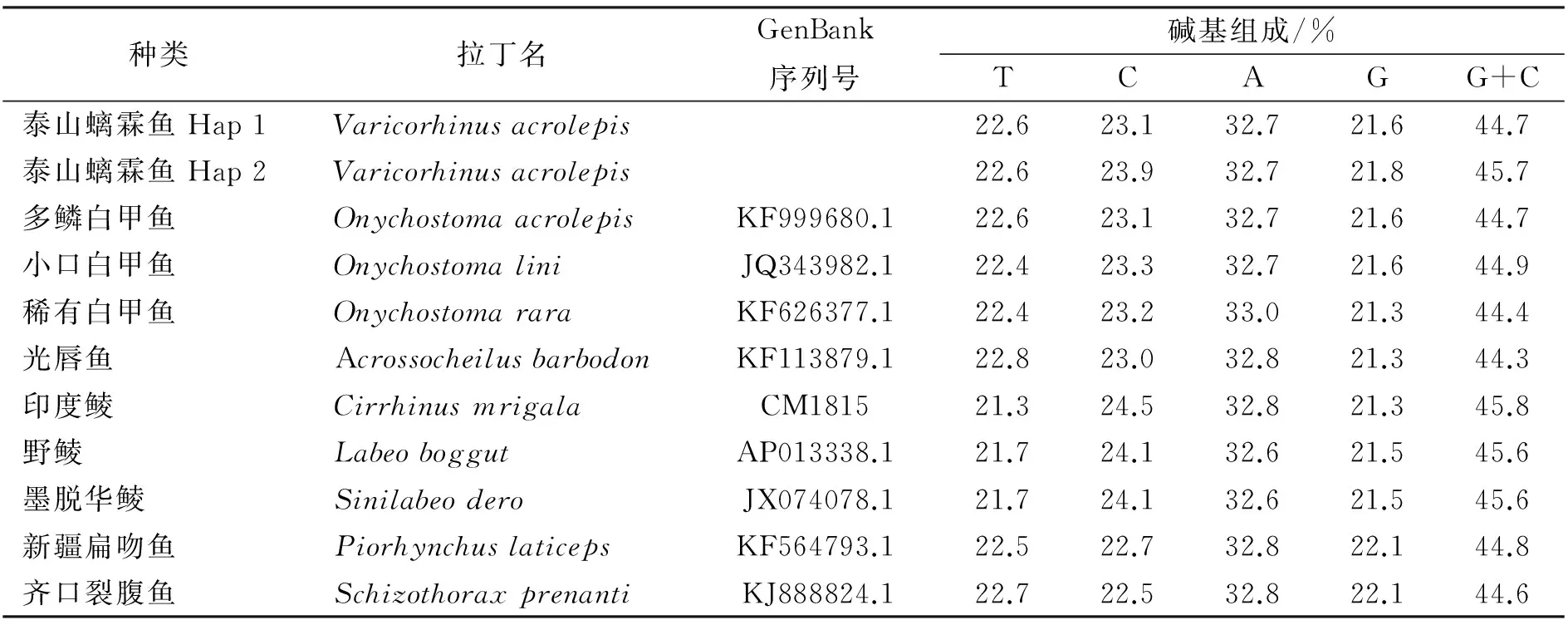
表1 樣品種類、GenBank序列號、堿基組成
采用MGEA 6.06軟件比較泰山螭霖魚和GenBank中查到的同源性較高的鯉科白甲魚屬(Onychostoma)、光唇魚屬(Acrossocheilus)、鯪屬(Cirrhinus)、華鯪屬(Sinilabeo)、野鯪屬(Labeo)、裂腹魚屬(Schizothorax)、扁吻魚屬(Aspiorhynchus)7屬9種魚的同源序列進行比對分析,結果見圖2。得到471個比對位點,有45個變異位點,包括4個插入/缺失位點、29個簡約信息位點和14個單突變子。泰山螭霖魚單倍型Hap1與多鱗白甲魚序列完全一致;與扁吻魚序列間的變異位點最多,有23個。

圖2 泰山螭霖魚及鯉科9種魚類線粒體16S rRNA序列直接的變異位點
泰山螭霖魚16S rRNA基因片段2種單倍型與GenBank中鯉科7屬部分魚類的遺傳距離見表2。可以看出,泰山螭霖魚2個單倍型間的遺傳距離為0.21%,泰山螭霖魚單倍型(Hap-1)與白甲魚屬幾種魚的的遺傳距離在0~3.07%,其中與多鱗白甲魚的遺傳距離為0。泰山螭霖魚與白甲魚屬幾種魚類遺傳距離最近,其次是光唇魚屬,再次是裂腹魚屬,與野鯪屬、華鯪屬、鯪屬魚類遺傳距離較遠,與扁吻魚屬遺傳距離最遠。
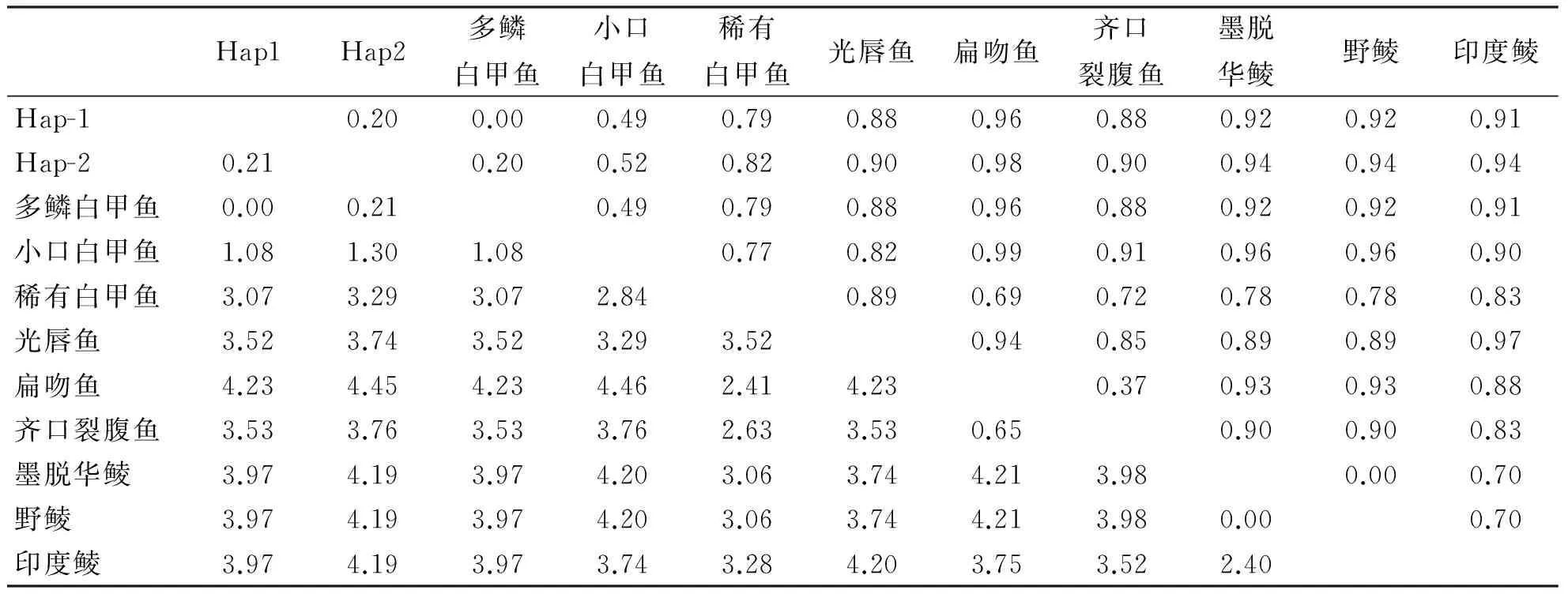
表2 泰山螭霖魚2種單倍型與鯉科9種魚類的遺傳距離 %
2.2.3 遺傳多樣性分析
利用DNASP 5.10軟件分析30個樣品泰山螭霖魚16S rRNA基因的遺傳多樣性,結果表明,其單倍型數(h)為2,單倍型多樣性(Hd)為0.067,核苷酸多樣性(Pi)為0.00015,平均核苷酸差異數(K)為0.067,遺傳多樣性水平非常低。
2.2.4 聚類及系統發育分析
將泰山螭霖魚16S rRNA基因2個單倍型序列與同源性較高的鯉科7屬部分魚類同源序列一起進行聚類分析,采用雙參數模型(Kimura 2-parameter)作為距離參數,分別以鄰接法(neighber-joining,NJ)和UPGMA法構建分子系統樹,自舉檢測(bootstrap)1000次計算各分枝置信度(圖3、圖4)。由2種聚類圖均可看出,7個屬的魚類可以聚為2個大的分支,泰山螭霖魚與白甲魚屬及光唇魚屬聚為一大類,另外5屬魚類聚為一大類。泰山螭霖魚的2個單倍型先與多鱗白甲魚聚在一起,再與粗須白甲魚、小口白甲魚聚在一起,然后與光唇魚屬聚為一類,泰山螭霖魚絕大多數個體為單倍型Hap-1,與多鱗白甲魚關系最近,與光唇魚屬親緣關系次之,與其他5屬的親緣關系較遠。
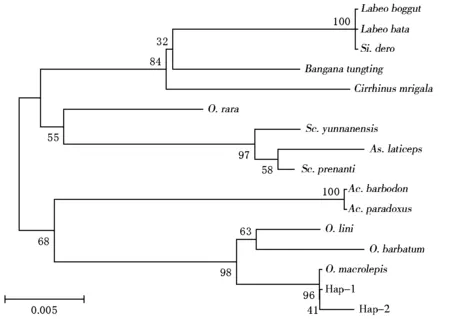
O.macrolepis:多鱗白甲魚;O.barbatum:粗須白甲魚;O.lini:小口白甲魚;Ac.barbodon:光唇魚;Ac.Paradoxus臺灣石賓魚;O.rara:稀有白甲魚;Sc.Prenanti:齊口裂腹魚;Sc.yunnanensis:云南裂腹魚;As.laticeps:新疆扁吻魚;Si.dero:墨脫華鯪;Labeoboggut:野鯪;Labeobata:巴塔野鯪;Banganatungting:洞庭華鯪;Cirrhinusmrigala:印度鯪。圖3同。圖3 基于16SrRNA序列構建的NJ系統樹
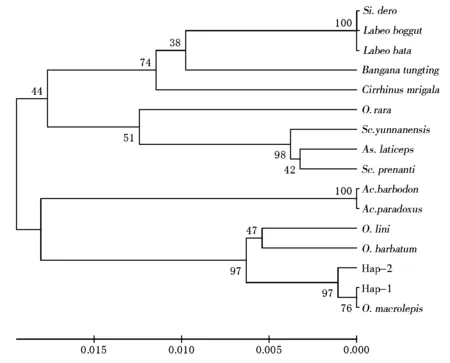
圖4 基于16S rRNA序列構建的UPGMA系統樹
3 討論
動物線粒體DNA(mitochondrial DNA,mt DNA)具有結構簡單、進化速度快、變異性大、母系遺傳及易于擴增等優點,在動物種群遺傳結構分析、物種及品系鑒定方面得到廣泛應用,線粒體16S rRNA基因進化速率低,比較保守,適合于種以上水平的分子系統學研究及遺傳分析研究[13]。在魚類進化研究中,16S rRNA基因已得到廣泛應用,如王茜等[13]、童馨等[14]、孫悅娜等[15]、邵愛華等[16]、任崗等[17]、陳子安等[18]分別對尖塘鱧、黃姑魚、日本沼蝦、暗紋東方鲀、石鱸科魚類、星蟲等線粒體16S rRNA基因序列和分子系統進化進行了研究。但是對于泰山螭霖魚16S rRNA的研究較少,目前尚未見報道,本研究通過分析泰山螭霖魚16S rRNA基因序列的差異及其與同亞科其他屬魚類同源序列進行比對,在分子水平上探討泰山螭霖魚與鲃亞科及鯉科部分魚類的系統發育關系,以確定其分類地位。
3.1 遺傳多樣性
泰山螭霖魚30個樣本擴增的16S rRNA基因468bp片段中,有1個突變位點,存在2種單倍型,核苷酸多樣性(Pi)為0.00015,平均核苷酸差異數(K)0.0067,其單倍型多樣性(Hd)為0.067,表明泰山螭霖魚群體具有較低的遺傳多樣性水平。這與公維華等[8]利用微衛星標記對泰山螭霖魚的遺傳多樣性的研究結果相似,即泰山螭霖魚野生群體和養殖群體遺傳結構無差異,是一個非常純合的群體。這是泰山螭霖魚特殊的生存環境和生活習性所決定的,它生存在泰山,分布在海拔270~800m的山澗中,缺乏與外界水域的交流,經長期的近交繁殖和瓶頸效應,多數基因已純合,加之環境的選擇壓力對其影響也不大,形成基因突變和重組的概率很小,所以會呈現出遺傳多樣性水平低的現象。
3.2 分類地位及分子系統進化分析
泰山螭霖魚的學名為Varicorhinusmacrolepis,屬突吻魚屬,有學者認為突吻魚屬和白甲魚屬是同物異名;也有學者認為突吻魚屬(Varicorhinus)分2個亞屬,即白甲魚亞屬(Onychostoma)和鏟頜魚亞屬(Scaphesthes)[19]。陳紅菊[11]通過對泰山螭霖魚線粒體Cyt b 和D-loop序列分析,發現泰山螭霖魚和多鱗鏟頜魚聚在一起,二者之間無任何分別,確定泰山螭霖魚就是多鱗鏟頜魚。本研究通過對泰山螭霖魚同源性較高的鯉科7屬部分魚類基于線粒體16S rRNA基因片段的聚類和系統發育分析表明,泰山螭霖魚2個單倍型中,絕大多數個體為單倍型為Hap1,與多鱗白甲魚(O.macrolepis)的16S rRNA基因片段序列完全一致,泰山螭霖魚Hap1首先與多鱗白甲魚聚在一起,再與粗須白甲魚、小口白甲魚聚為一類。泰山螭霖魚系統進化分析結果與陳紅菊的研究結果相似,即泰山螭霖魚所在的突吻魚屬與光唇魚屬的親緣關系最近,與其他屬魚類的親緣關系較遠。
[1]岳永生.泰山螭霖魚的資源保護與小康村建設[J].山東農業科學,2006,(增刊):74~75.
[2]楊越峰.泰山赤鱗魚生物學特性及其人工繁殖技術[J].中國水產,2006,49(3):28~39,51.
[3]張安才.泰山赤鱗魚的生物學及人工養殖技術[J].齊魯漁業,2005,22(8):31~32.
[4]張金花,王樹迎.泰山螭霖魚腸道的解剖學研究[J].山東農業大學學報,2003,34(4):579~581.
[5]劉自杰,肖順利,黃金果,等.四種營養素對泰山螭霖魚生長效果影響的研究[J].淡水漁業,2002,32(2):34~37.
[6]張慶朝,王慧秦,孜娟泰,等.泰山赤鱗魚同工酶的研究[J].動物學研究,1994,15(2):62~67.
[7]PANG Q X,ZHAO B S,YAO D F,etal.The Karyotype of Chi-lin Fish (Varicorhinusmacrolepis) from Taishan Mountain[J].Animal Science,2012,13:649~651.
[8]公維華,宋憬愚,姜運良,等.用微衛星標記分析泰山螭霖魚的遺傳多樣性[J].山東農業大學學報,2004,35(3):339~342.
[9]郭金峰.三個黃顙魚群體遺傳多樣性的微衛星標記分析[D].泰安:山東農業大學,2007.
[10]汪艷宏,李潔,黃麗波,等.赤鱗魚GH全長cDNA擴增和生物信息學分析[J].畜牧與獸醫,2012,44(增刊):336~339.
[11]陳紅菊.泰山赤鱗魚BMP11基因表達規律及分子進化研究[D].泰安:山東農業大學,2008.
[12]王慧,高建剛,王樹迎.泰山赤鱗魚全同胞個體甲基化位點的差異研究[A].中國水產學會學術年會論文集[C].2008.
[13]王茜,齊興柱,駱劍,等.尖塘鱧屬魚類線粒體16S rRNA基因序列變異及分子系統進化[J].海南大學學報,2009,27(3):245~251.
[14]童馨,杜博,喻達輝,等.淺色黃姑魚線粒體16S rRNA基因片段序列特征分析[J].海洋水產研究,2007,28(3):85~91.
[15]孫悅娜,馮建彬,李家樂,等.日本沼蝦三群體線粒體16S rRNA基因片段序列的差異與系統進化[J],動物學雜志,2007,42(1):59~66.
[16]邵愛華,杜建,陳葵,等.暗紋東方鲀線粒體DNA 16S rRNA基因克隆、測序及在分子系統發育分析中的應用[J].江蘇農業科學,2009,37(2):15~19.
[17]任崗,章群,錢開誠,等.12種石鱸科魚類線粒體16S rRNA基因的部分序列分析[J].熱帶海洋學報,2007,26(3):48~52.
[18]陳子安,杜曉東,王慶恒,等.3種星蟲線粒體16S rRNA、COⅠ和Cyt b基因片段的序列比較[J].廣東海洋大學學報,2007,24(4):3~10.
[19]尹邦一,代應貴.白甲魚屬魚類研究進展[J].水產科學,2014,33(1):63~68.
2016-07-19
山東省農業良種工程項目;山東省現代農業產業體系魚類創新團隊項目(SDAIT-12-04)。
楊玲(1967-),女,研究員,主要從事魚類遺傳育種研究。通信作者:張金路,18953160608@163.com。
S917; Q731
A
1673-1409(2016)33-0037-06
[引著格式]楊玲,鞏俊霞,李嫻,等.泰山螭霖魚線粒體16S rRNA基因片段序列及分子系統發育分析[J].長江大學學報(自科版),2016,13(33):37~42.
