水稻淡綠葉突變體HM133的遺傳分析與基因定位
2016-12-07 07:56:33施勇烽賀彥郭丹呂向光黃奇娜吳建利
中國水稻科學 2016年6期
關鍵詞:水稻
施勇烽 賀彥 郭丹 呂向光,2 黃奇娜 吳建利,*
(1中國水稻研究所 水稻生物學國家重點實驗室/國家水稻改良中心, 杭州 310006; 2中國農業科學院 作物科學研究所, 北京 100081;# 共同第一作者; *通訊聯系人, E-mail: beshangd@163.com)
?
水稻淡綠葉突變體HM133的遺傳分析與基因定位
施勇烽1,#賀彥1,#郭丹1呂向光1,2黃奇娜1吳建利1,*
(1中國水稻研究所 水稻生物學國家重點實驗室/國家水稻改良中心, 杭州 310006;2中國農業科學院 作物科學研究所, 北京 100081;#共同第一作者;*通訊聯系人, E-mail: beshangd@163.com)
SHI Yongfeng, HE Yan, GUO Dan, et al. Genetic analysis and gene mapping of a pale green leaf mutantHM133 in rice. Chin J Rice Sci, 2016, 30(6): 603-610.
EMS誘導秈稻品種IR64獲得淡綠葉突變體HM133。與野生型IR64相比,HM133播種后的第6周和第15周的光合色素含量以及抽穗期的凈光合速率顯著降低,氣孔導度則明顯上升;此外,突變體株高、每穗實粒數和結實率等農藝性狀也較野生型顯著下降。葉綠體超微結構分析表明,分蘗期HM133類囊體基粒片層形狀不規則,堆疊凌亂、排列疏松。遺傳分析表明HM133淡綠葉性狀受單隱性核基因控制。通過分子標記將該基因定位于第3染色體長臂RM143和RM3684之間。該區間內包含編碼鎂螯合酶D亞基的基因OsCHLD。序列分析表明HM133中該基因第10外顯子上有一個從G突變為A的單堿基變異,導致編碼的氨基酸由精氨酸變成谷氨酸,推測OsCHLD基因即為控制HM133淡綠葉表型的候選基因。
水稻; 淡綠葉; 基因定位; 鎂螯合酶; 光合作用
葉片是植物進行光合作用的主要場所。葉綠體是進行光合作用的重要細胞器,葉綠體發育異常、光合色素含量改變以及光合作用復合體的穩定性等都會影響影響植物的光合作用效率。常見的葉色變異大多與光合色素合成與降解,葉綠體的結構、組成和生理狀態異常有關,通常會導致葉片出現白化、黃化、淡綠、黃綠和條紋等突變表型。在水稻[1,2]、玉米[3,4]、大麥[5]、擬南芥[6]和油菜[7]等植物中均有葉色突變體的報道。
目前水稻中已經鑒定了大量的葉色突變體(http://www.shigen.nig.ac.jp/rice/oryzabase/),分別涉及葉綠素代謝、光敏色素代謝以及葉綠體發育異常。其中,影響葉綠素生物合成途徑相關基因的突變是研究報道最為深入的一類。對擬南芥的研究發現葉綠素生物合成途徑共包括15步反應,涉及15種酶,并分離了 27個編碼這些酶的基因[8]。水稻中目前也已克隆多個葉綠素合成途徑中相關酶的基因,如OsCHLH、OsCHLD、OsCHLI、DVR、CAO1、CAO2、YGL1和PORB[2,9-14]。另外,四吡咯代謝分支途徑即光敏色素合成途徑中相關基因的突變也能造成葉色突變,例如水稻亞鐵血紅素加氧酶基因的變異能反饋作用于葉綠素合成途徑,從而使葉綠素的合成發生異常[15,16]。除了約100個葉綠體基因組自身編碼的蛋白外,葉綠體中含有大約3000個由核基因編碼的蛋白,相關蛋白的突變會影響葉綠體的正常發育,導致葉色變異、植株發育遲緩等癥狀,如編碼類囊體結合蛋白基因ZN突變可導致水稻產生斑馬葉表型[17],編碼PPR蛋白的基因YSA的突變致使水稻產生白化表型[18],編碼葉綠體SRP43蛋白[19]和SRP54蛋白[20]基因的變異均導致水稻葉色變成淡綠色。
本研究利用秈稻IR64經EMS誘導后篩選獲得一份葉色突變體HM133,對其開展表型性狀、生理特性等方面的研究,同時對突變基因開展了遺傳分析和基因定位,結合測序分析結果推測水稻鎂螯合酶D亞基基因(OsCHLD)是控制HM133淡綠葉性狀的候選基因。
1 材料與方法
1.1 供試材料
本研究采用的淡綠葉突變體HM133是由秈稻品種IR64經過EMS誘變得到。經過連續多代自交,該葉色突變性狀無論種植于海南陵水或浙江富陽,在溫室條件或大田環境下均能穩定遺傳。2015年將該突變體種植于中國水稻研究所富陽實驗基地,在成熟期隨機選取野生型和突變體各3株考查株高、穗長、有效穗數、結實率和千粒重等農藝性狀,取平均值進行分析。
1.2 群體構建及遺傳分析
以突變體HM133為母本,分別與正常葉色品種IR24和熱研1號配制雜交組合,觀察F1的表型并收獲F1單株種子,觀察并統計HM133/IR24的F2群體中正常葉和淡綠葉水稻的株數,用于遺傳分析。HM133/熱研1號的F2群體中淡綠葉表型單株用于DNA提取和基因定位。
1.3 光合色素含量和光合參數測定
播種后第6周和第15周分別取野生型和突變體HM133倒2葉樣品,稱取約0.3 g,剪成0.3~0.5 cm長度的片段,在常溫黑暗條件下用95%乙醇浸提48 h,然后用PerkinElmer分光光度計分別測定在470 nm、649 nm和665 nm三個波長處的光吸收值。播種后第6周和第15周各取3株設定生物學重復,參照Arnon和Wellburn等[21,22]的方法計算葉綠素和類胡蘿卜素的含量。
于水稻抽穗期利用便攜式光合測定儀LI-6400(LI-COR,USA)在晴好天氣上午9:00-11:00,田間測定野生型和突變體HM133劍葉光合作用指標,光合參數設定參照Huang等[23]的方法。每株重復3次測定,取平均值。
1.4 葉綠體超微結構觀察
選取野生型和突變體HM133分蘗期倒2葉葉片切成1 mm片段,在2.5% 戊二醛溶液中抽真空固定。參照Lv等[19]的方法進行樣品制備,于浙江大學農業與生物技術學院用Tecnai G2F20 S-TWIN型透射電子顯微鏡(FEI,USA)觀察葉綠體超微結構。
1.5 基因定位
采用簡易法提取親本和單株DNA[24],分別取HM133/熱研1號F2群體正常葉和淡綠色葉單株各10株,以等量葉片構建野生型DNA池和突變體DNA池。利用水稻12條染色體上172對SSR標記,對親本HM133和熱研1號間進行多態性標記篩選,再將篩選到的標記用于野生型池和突變體池間的多態性分析,找到兩池間有多態的標記,用于F2群體中淡綠葉單株的基因型分析,初步確定突變基因的位置。從Gramene數據庫(http://www.gramene.org/)下載SSR標記引物序列,由上海生工生物工程有限公司進行合成。參照Shi等[25]的方法進行PCR和產物檢測。
1.6 候選基因OsChlD的測序和驗證
利用水稻基因組信息注釋系統(http://rice.plantbiology.msu.edu),查詢目標區間內的基因。根據OsChlD基因的cDNA序列設計引物(正向引物5′-TCTCTCCCTCCCCTCCCATG-3′,反向引物5′-GCTCTCCAGGATCACGAACTCT-3′),并對HM133、IR64和IR24進行PCR,由鉑尚生物技術(上海)有限公司進行測序。
根據測序結果的比對,找到突變位點,在突變位點附近根據OsChlD基因組序列設計引物(正向引物5′-TGCTTGGCACCTTTATCACA-3′,反向引物5′-GCCATTCTTTTGGCTCTCAT-3′),對HM133、 IR24及其F2單株進行DNA擴增,擴增產物用限制性內切酶TaqⅠ進行酶切驗證。
表1 實時定量PCR引物
Table 1. Primers used in real-time PCR.

基因Gene正向引物(5'-3')Forwardprimer(5'-3')反向引物(5'-3')Reverseprimer(5'-3')OsChlDGGAAAGAGAGGGCATTAGCAATACGATCAAGTAAGTGTTOsChlIAGTAACCTTGGTGCTGTGAATCCATCAACATTCAACTCTGOsChlHCTATACATTCGCCACACTTATCACACAACTCCCAAGHEMA1CGCTATTTCTGATGCTATGGGTTCTTGGGTGATGATTGTTTGGPORATGTACTGGAGCTGGAACAACAAGAGCACAGCAAAATCCTAGACGCAO1GATCCATACCCGATCGACATCGAGAGACATCCGGTAGAGCCab1RAGATGGGTTTAGTGCGACGAGTTTGGGATCGAGGGAGTATTTCab2RTGTTCTCCATGTTCGGCTTCTGCTACGGTCCCCACTTCACTpsaAGCGAGCAAATAAAACACCTTTCGTACCAGCTTAACGTGGGGAGpsbACCCTCATTAGCAGATTCGTTTTATGATTGTATTCCAGGCAGAGCrbcLCTTGGCAGCATTCCGAGTAAACAACGGGCTCGATGTGATArbcSTCCGCTGAGTTTTGGCTATTTGGACTTGAGCCCTGGAAGGYGL1CAGTCTCCAATGGCCACCTTGCTTTCATCAGTGGCTGGSPPCGGAGAGGAAACATAATGACATAGGCATTTGTCTTTGTCTCPPR1CTAAGACCGAATGACAAATGCGCACTGCCAACAAGAATACCDVRCGAGCCCAGGTTCATCAAGGTGCCCTCCCGATCTTGCCGAACTCCUbiquitinGCTCCGTGGCGGTATCATCGGCAGTTGACAGCCCTAG
1.7 RNA提取和RT-PCR
采用Trizol法提取IR64與淡綠葉突變體HM133分蘗期倒2葉的總RNA,利用ReverTra Ace qPCR RT Master Mix with gDNA Remover試劑盒(TOYOBO, 日本)將RNA反轉錄為cDNA。采用SYBR?PremixExTaqTMⅡ (Tli RNaseH Plus)試劑盒(TaKaRa,日本)和Thermal Cycle Dice?Real Time System(TaKaRa, 日本)進行實時定量PCR,分析葉綠素合成與代謝途徑中相關基因的表達情況。以水稻Ubiquitin為內參基因,相關基因的特異性引物見表1。
2 結果與分析
2.1 HM133的表型及主要農藝性狀
突變體HM133葉色在苗期、分蘗期均表現為淡綠色(圖1),該表型伴隨整個生育期。突變體HM133的主要農藝性狀如有效穗數、穗長和千粒重與野生型相比未發生明顯變化,但結實率、株高和每穗實粒數均顯著低于野生型(表2)。

A—野生型IR64和突變體HM133幼苗表型;B—野生型IR64和突變體HM133分蘗期的表型。
A, Phenotype of the wide type IR64 and the mutantHM133 at the seedling stage; B, Phenotype of the wide type IR64 and the mutantHM133 at the tillering stage.
圖1 野生型IR64和突變體HM133的不同時期植株表型
Fig. 1. Phenotype of the wide-type IR64 and the mutant HM133 at different growth stages.
2.2 HM133的光合色素含量和光合參數
在播種后第6周和第15周分別測定了突變體和野生型葉綠素和類胡蘿卜素含量。第6周和第15周時突變體的總葉綠素含量與野生型相比分別下降了46.0%和47.7%,胡蘿卜素含量下降了38.8%和47.6%,兩個時期HM133的總葉綠素含量、葉綠素a和葉綠素b含量與野生型相比均存在極顯著差異(表3)。
表2 野生型IR64和突變體HM133的主要農藝性狀
Table 2. Agronomic traits of the wild-type IR64 and mutant HM133.

材料Material株高Plantheight/cm有效穗數No.ofproductivepanicles穗長Paniclelength/cm每穗實粒數Numberoffilledgrainsperpanicle結實率Seed-settingrate/%千粒重1000-grainweight/gIR64113.0±1.714.0±1.025.0±1.481.2±7.174.6±1.627.52±0.38HM133108.7±0.6*12.7±2.124.9±0.362.9±7.3*68.8±2.1*28.35±0.35
*HM133與IR64在0.05水平上差異顯著。
*Significant difference between IR64 andHM133 at 0.05 level.
表3 不同生長時期HM133與IR64葉片光合色素含量的比較
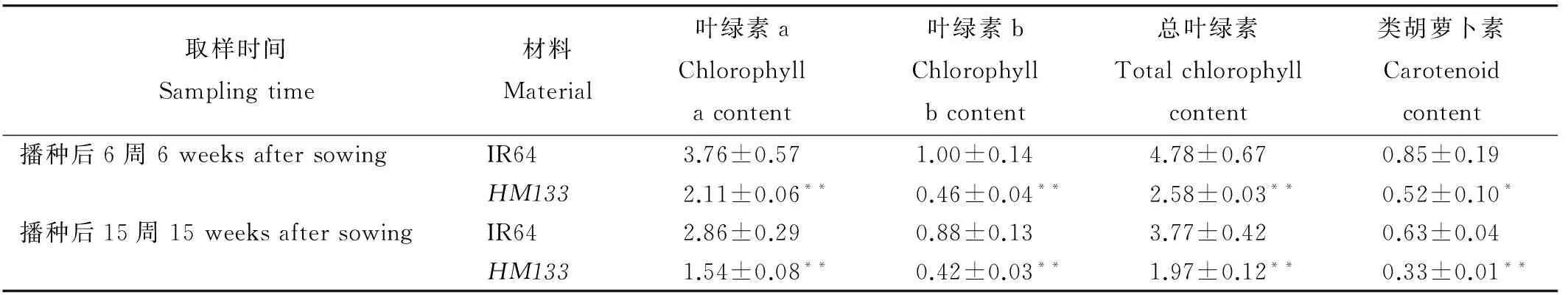
Table 3. Comparison of photosynthetic pigment contents between HM133 and IR64 at different growth stages. mg/g
*HM133與IR64在0.05水平上差異顯著;**HM133與IR64在0.01水平上差異顯著。
*Significant difference between IR64 andHM133 at 0.05 level;**Significant difference between IR64 andHM133 at 0.01 level.
表4 抽穗期野生型IR64和突變體HM133的劍葉光合特性
Table 4. Photosynthetic parameters of flag leaf of IR64 and HM133 at the heading stage.

材料Material凈光合速率Pn/(μmol·m-2s-1)氣孔導度GS/(mol·m-2s-1)胞間CO2濃度Ci/(μmol·mol-1)蒸騰速率Tr/(mmol·m-2s-1)IR6414.45±1.780.57±0.13309.13±7.304.12±0.64HM13312.74±1.38*0.73±0.09*313.75±6.414.26±0.25
*HM133與IR64在0.05水平上差異顯著。
*Difference between IR64 andHM133 was significant at 0.05 level.
為分析突變體光合色素含量的降低對光合作用的影響,抽穗期對IR64和HM133的光合參數進行測定,發現突變體的凈光合速率降低了11.8%,氣孔導度上升了26.3%,而胞間CO2濃度和蒸騰速率沒有顯著差異(表4)。
2.3 葉綠體超微結構觀察
分蘗期分別對野生型和淡綠葉突變體HM133葉片進行葉綠體結構的電鏡觀察。結果顯示野生型IR64的葉綠體基質濃密、基粒片層堆疊較厚、排列緊密(圖2-A、B),而HM133的基粒數量明顯減少,基粒片層形狀不規則,片層堆疊較少、排列疏松(圖2-C、D)。結果表明HM133葉綠體的發育受到了嚴重影響。
2.4 HM133的遺傳分析
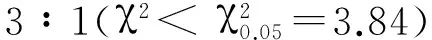
2.5 pglHM133基因定位及其候選基因預測
HM133/熱研1號的F2群體中85株淡綠葉單株作為定位群體,并采用分池法構建了淡綠葉和正常葉色DNA池各1個。選用分布于12條染色體上的172對SSR標記,對HM133和熱研1號進行多態性標記的篩選,獲得的128對多態性標記用于淡綠葉與正常綠葉DNA池的多態性分析,其中第3條染色體上RM520、RM85和RM143表現DNA池的多態。進一步用這3個標記分析F2群體的85株淡綠葉單株,分別檢測到20株、28株和4株重組個體,其中RM143與標記基因緊密連鎖。為更準確定位該基因,根據已經公布的SSR引物在RM143附近找到RM16108、RM16103、RM16097和RM3684在親本間有多態的標記,進一步分析發現僅標記RM3684檢測到10株重組個體,其他3個標記均無交換發生,最終將該基因定位于第3染色體的長臂RM3684和RM143之間,物理距離約為3.1 Mb(圖3-A)。
A和B-野生型IR64; C和D-突變體HM133。 S-淀粉粒; C-葉綠體; G-基粒; SL-基質片層; OG-嗜餓顆粒。
A and B, IR64; C and D,HM133. S, Starch granule; C, Chloroplast; G, Granum; SL, Stroma lamella; OG, Osmiophilic granule.
圖2 野生型IR64和突變體HM133的葉綠體超微結構
Fig. 2. Chloroplast ultrastructure of IR64 and HM133.
通過水稻基因組信息注釋系統(http://rice.plantbiology.msu.edu/)查詢pglHM133目標區間內有超過200個候選基因,區間中包含已被Zhang等[10]注釋為OsChlD基因(登錄號為LOC_Os03g59640)。OsChlD編碼鎂螯合酶D亞基,參與葉綠素合成過程,對葉綠體發育具有重要作用。序列分析表明,IR64與IR24的OsChlD基因cDNA序列一致,而突變體HM133的第10外顯子上存在一個單堿基突變,由G突變為A(圖3-B),對應的氨基酸由精氨酸變為谷氨酸。該位點突變影響了限制性內切酶TaqⅠ 的特異性識別,用IR24/HM133的親本、F1和F2群體進行驗證,通過DNA擴增獲得包含突變位點約193 bp大小片段,用TaqⅠ酶切后發現F2中淡綠葉單株和HM133的擴增片段不能被TaqⅠ酶切,而IR24能被TaqⅠ完全酶切,F1則被部分酶切,F2中正常葉單株有部分酶切和完全酶切兩種情況(圖3-C)。這一結果表明突變體HM133的淡綠葉表型可能是由OsChlD基因的單堿基替換導致。
2.6 葉綠素合成和葉綠體發育相關基因的表達
利用實時熒光定量PCR比較HM133及其野生型IR64中OsChlD基因和其他葉綠素合成、葉綠體發育相關基因的表達情況。結果表明,與野生型相比,突變體中編碼鎂螯合酶亞基的OsChlD、OsChlI和OsChlH的表達量明顯下降,葉綠素合成相關基因HEMA1(谷氨酰t-RNA還原酶)、YGL1(葉綠素合成酶)、DVR(聯乙烯還原酶)和CAO1(葉綠素酸酯氧化酶)的表達量也顯著下降;葉綠體發育相關的psbA(D1蛋白)、psaA(光合系統Ⅰ葉綠素脫輔基蛋白)、rbcL(RUBP羧化酶大亞基)、SPP(基質加工肽酶)表達顯著下調,而Cab1R和Cab2R(捕光葉綠素a/b結合蛋白)表達量顯著上升;PORA(原葉綠素酸酯氧化還原酶)、rbcS(RUBP羧化酶小亞基)、PPR1(三角狀五肽重復蛋白)的表達并沒有顯著變化(圖4)。上述結果表明,OsChlD的變異導致與葉綠素合成以及葉綠體發育相關的眾多基因的表達發生明顯改變。
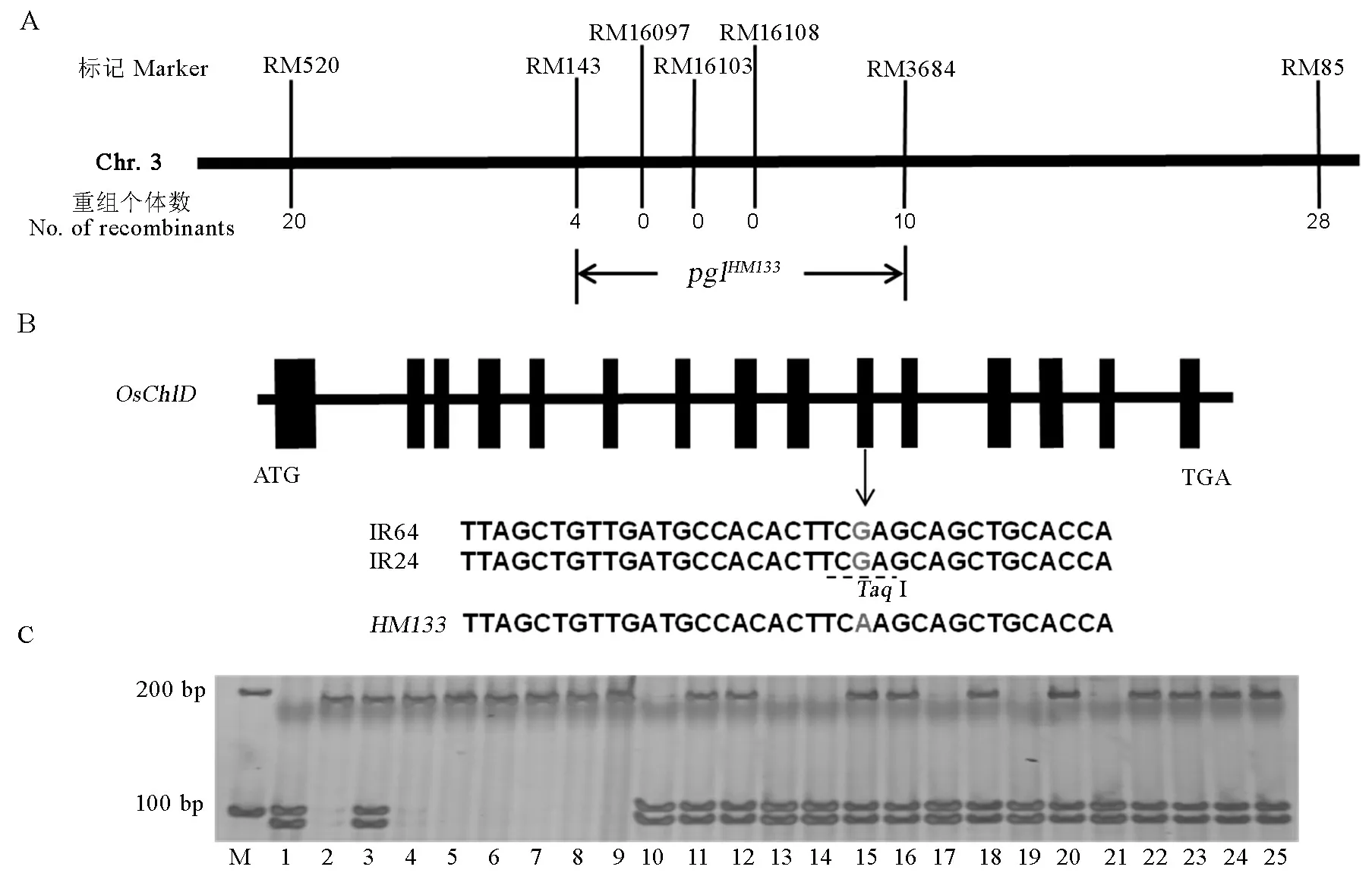
A-pglHM133初步定位; B-候選基因OsChlD序列分析. 灰色部分表示HM133的1541位堿基G突變為A; C-IR24/HM133 F2群體的TaqⅠ酶切驗證. M-分子量標記; 1-IR24; 2-HM133; 3-F1; 4~9-F2中淡綠葉單株; 10~25-F2群體中正常葉單株。
A,Primary mapping ofyglHM133on chromosome 3; B, Sequence analysis of the candidate geneOsChlD. The gray letters indicate single base substitution from G to A at position 1541; C, Mutation base ofOsChlDconfirmed withTaqI restriction enzyme digestion. M, Molecular marker; 1, IR24; 2,HM133; 3, F1; 4-9, Pale green plant of F2; 10-25, Normal plant of F2.
圖3 pglHM133定位及候選基因預測
Fig. 3. Location of pglHM133and candidate gene prediction.
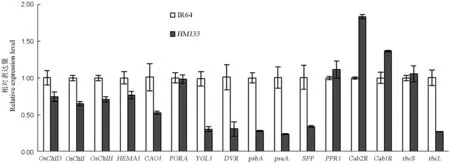
圖4 RT-PCR分析野生型IR64與HM133 中葉綠素合成、葉綠體發育相關基因的表達
Fig. 4. Expression of genes associated with chlorophyll biosynthesis and chloroplast development in IR64 and HM133 by real-time PCR.
3 討論
植物中葉綠素的累積使葉片呈現綠色,葉綠素的生物合成需要包括鎂螯合酶在內的15種酶的參與。鎂螯合酶是由D、I和H三亞基組成的復合體,四吡咯化合物原卟啉Ⅸ在鎂螯合酶催化下與鎂離子螯合形成鎂原卟啉Ⅸ[26]。植物葉色突變體中已鑒定了一些鎂螯合酶的突變體,大麥的突變體xantha-g44和xantha-g45是由于編碼鎂螯合酶D亞基突變造成葉綠素合成受阻,分別產生黃葉和黃綠葉表型[5],擬南芥T-DNA插入突變體AtchlD-KO存在白化致死表型,水稻中突變體Chlorina-1和ygl7在D亞基基因的不同位點發生突變,ygl7在整個生育期均表現為黃綠葉,Chlorina-1僅幼苗期的前2~3周出現黃綠葉表型[10,13]。玉米鎂螯合酶I亞基突變體Oy1為半顯性突變體,表現為黃化致死[4],水稻和擬南芥的I亞基突變體則均表現為黃綠葉[6,10]。擬南芥和水稻鎂螯合酶H亞基的突變體分別產生淺綠和黃綠葉表型[2,6]。
本研究中的淡綠葉突變體HM133是EMS誘變秈稻IR64獲得,其葉綠素含量明顯低于野生型IR64,灌漿期凈光合速率明顯下降,類囊體發育異常,遺傳分析表明HM133淡綠葉性狀受單隱性核基因控制。通過分子標記將pglHM133基因定位于第3染色體長臂RM143和RM3684之間,區間內包含編碼鎂螯合酶D亞基基因OsChlD,序列分析發現HM133中OsChlD基因第10外顯子上存在G突變為A的單堿基變異,與已報道了另4個OsChlD等位基因Chl1[10]、ygl98[27]、ygl3[28]和ygl7[13]突變位點均不同,是一個新的OsChlD等位基因。這5個OsChlD突變體的表型不完全相同,可能由于不同的突變蛋白引起,也可能是受到遺傳背景的影響。ygl7在日本晴背景下產生黃化葉,而在810S背景下則表現為黃綠葉[13],表明OsChlD控制葉色表型的機理非常復雜,受到遺傳背景的影響。Deng等[13]研究表明,ygl7在810S遺傳背景下,OsChlD、OsChlI和OsChlH的表達顯著上升,在日本晴背景下的ygl7-NIL則與野生型的表達基本相同。本研究發現突變體中OsChlD、OsChlI和OsChlH的表達量較野生型顯著下降,該結果與Deng等[13]的結果不同,可能與遺傳背景的差異有關,也可能是突變位點的不同引起的。突變體HM133中核基因編碼的HEMA1、YGL1、DVR、CAO1和葉綠體編碼基因PsaA、PsbA、rbcL的表達量呈顯著下降,而核基因編碼的Cab1R和Cab2R的表達呈顯著上升,表明OsChlD基因突變同樣影響了其他葉綠素合成酶基因和葉綠體發育相關基因的表達。Deng等[13]研究發現OsChlD突變蛋白ygl7,能行使部分OsChlD功能,能促進光合作用和光能的高效利用,結實率等性狀與野生型無明顯差異。而本研究中突變體的凈光合速率顯著降低,株高、結實率和每穗實粒數也顯著低于野生型,可能是由于OsChlD突變位點不同造成相應突變蛋白功能差異。另外,豌豆中ChlD亞基基因沉默植株中發現活性氧物質的積累[29],水稻中D亞基突變體是否存在同樣的現象仍有待進一步研究。
[1] 李智強, 朱丹, 王志龍, 等. 水稻黃綠葉突變體djyg的遺傳分析與基因定位. 中國水稻科學, 2015, 29: 601-609.
Li Z Q, Zhu D, Wang Z L,et al. Genetic analysis and gene mapping of a yellow-green leaf mutantdjygin rice.ChinJRiceSci, 2015, 29: 601-609. (in Chinese with English abstract)
[2] Jung K H, Hur J, Ryu C H, et al. Characterization of a rice chlorophyll-deficient mutant using the T-DNA gene-trap system.PlantCellPhysiol, 2003, 44: 463-472.
[3] 韓帥, 王立靜, 鐘世宜, 等. 一個新的玉米葉色突變體的遺傳分析及基因定位. 玉米科學, 2012, 20: 26-28.
Han S, Wang L J,Zhong S Y, et al. Genetic analysis and gene mapping of a new leaf color mutant in maize.JMaizeSci, 2012, 20: 26-28. (in Chinese with English abstract)
[4] Sawers R J, Viney J, Farmer P R, et al. The maizeOilyellow1 (Oy1) gene encodes the I subunit of magnesium chelatase.PlantMolBiol, 2006, 60: 95-106.
[5] Axelsson E, Lundqvist J, Sawicki A, et al. Recessiveness and dominance in barley mutants deficient in Mg-chelatase subunit D, an AAA protein involved in chlorophyll biosynthesis.PlantCell, 2006, 18: 3606-3616.
[6] Mochizuki N,Brusslan J A, Larkin R, et al.Arabidopsisgenomesuncoupled5 (GUN5) mutant reveals the involvement of Mg-chelatase H subunit in plastid-to-nucleus signal transduction.ProcNatlAcadSci, 2001, 98: 2053-2058.
[7] 肖華貴, 楊煥文, 饒勇, 等. 甘藍型油菜黃化突變體的葉綠體超微結構、氣孔特征參數及光合特性. 中國農業科學, 2013, 46: 715-727.
Xiao H G, Yang H W, Rao Y,et al. Analysis of chloroplast ultrastructure, stomatal characteristic parameters and photosynthetic characteristics of chlorophyll-reduced mutant inBrassicanapusL.SciAgricSin, 2013, 46: 715-727. (in Chinese with English abstract)
[8] Beale S I. Green genes gleaned.TrendsPlantSci, 2005, 10: 309-312.
[9] Lee S, Kim J H, Yoo E S, et al. Differential regulation of chlorophyll a oxygenase genes in rice.PlantMolBiol, 2005, 57: 805-818.
[10] Zhang H, Li J, Yoo J H, et al. RiceChlorina-1 andChlorina-9 encode ChlD and ChlI subunits of Mg-chelatase, a key enzyme for chlorophyll synthesis and chloroplast development.PlantMolBiol, 2006, 62: 325-337.
[11] Wu Z, Zhang X, He B, et al. A chlorophyll-deficient rice mutant with impaired chlorophyllide esterification in chlorophyll biosynthesis.PlantPhysiol, 2007, 145: 29-40.
[12] Wang P, Gao J, Wan C, et al. Divinyl chlorophyll(ide) a can be converted to monovinyl chlorophyll(ide) a by a divinyl reductase in rice.PlantPhysiol, 2010, 153: 994-1003.
[13] Deng X J, Zhang H Q, Wang Y, et al. Mapped clone and functional analysis of leaf-color geneYgl7 in a rice hybrid (OryzasativaL. ssp.indica).PLoSOne, 2014, 9(6): e99564.
[14]Sakuraba Y, Rahman M L, Cho S H, et al. The rice faded green leaf locus encodes protochlorophyllide oxidoreductase B and is essential for chlorophyll synthesis under high light conditions.PlantJ, 2013, 74: 122-133.
[15] Li Q Z, Zhu F Y, Gao X, et al. Young Leaf Chlorosis 2 encodes the stroma-localized heme oxygenase 2 which is required for normal tetrapyrrole biosynthesis in rice.Planta, 2014, 240: 701-712.
[16] Chen H, Cheng Z, Ma X, et al. A knockdown mutation ofYELLOW-GREENLEAF2 blocks chlorophyll biosynthesis in rice.PlantCellRep, 2013, 32: 1855-1867.
[17] Li J, Pandeya D, Nath K, et al.ZEBRA-NECROSIS, a thylakoid-bound protein, is critical for the photoprotection of developing chloroplasts during early leaf development.PlantJ, 2010, 62: 713-725.
[18] Su N, Hu M L, Wu D X, et al. Disruption of a rice pentatricopeptide repeat protein causes a seedling-specific albino phenotype and its utilization to enhance seed purity in hybrid rice production.PlantPhysiol, 2012, 159: 227-238.
[19] Lv X G, Shi Y F, Xu X, et al.Oryzasativachloroplast signal recognition particle 43 (OscpSRP43) is required for chloroplast development and photosynthesis.PLoSONE, 2015, 10(11): e143249.
[20] Zhang F, Luo X, Hu B, et al.YGL138(t), encoding a putative signal recognition particle 54 kDa protein, is involved in chloroplast development of rice.Rice, 2013, 6(1): 7.
[21]Wellburn A R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution.JPlantPhysiol, 1994, 144: 307-313.
[22]Arnon D I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase inBetavulgaris.PlantPhysiol, 1949, 24: 1-15.
[23]Huang Q N, Shi Y F, Zhang X B, et al. Single base substitution inOsCDC48 is responsible for premature senescence and death phenotype in rice.JIntegrPlantBiol, 2016, 58: 12-28.
[24]盧揚江, 鄭康樂. 提取水稻DNA的一種簡易方法. 中國水稻科學, 1992, 6(1): 47-48.
Lu Y J, Zheng K L. A simple method for isolation of rice DNA.ChinJRiceSci, 1992, 6(1): 47-48.(in Chinese with English abstract)
[25]Shi Y F, Chen J, Liu W Q, et al. Genetic analysis and gene mapping of a new rolled-leaf mutant in rice (OryzasativaL.).SciChinaCLifeSci, 2009, 52: 885-890.
[26]Masuda T. Recent overview of the Mg branch of the tetrapyrrole biosynthesis leading to chlorophylls.PhotosynthRes, 2008, 96: 121-143.
[27]孫小秋, 王兵, 肖云華, 等. 水稻ygl98黃綠葉突變基因的精細定位與遺傳分析. 作物學報, 2011, 37(6): 991-997.
Sun X Q, Wang B, Xiao Y H,et al. Genetic analysis and fine-mapping ofygl98 yellow-green leaf gene in rice.ActaAgronSin, 2011, 37(6): 991-997. (in Chinese with English abstract).
[28]Tian X Q, Ling Y H, Fang L K, et al. Gene cloning and functional analysis of yellow green leaf3 (ygl3) gene during the whole-plant growth stage in rice.Genes&Genom, 2013, 35: 87-93.
[29]Luo T, Luo S, Araujo W L, et al. Virus-induced gene silencing of pea CHLI and CHLD affects tetrapyrrole biosynthesis, chloroplast development and the primary metabolic network.PlantPhysiolBiochem, 2013, 65: 17-26.
Genetic Analysis and Gene Mapping of a Pale Green Leaf MutantHM133 in Rice
SHI Yong-feng1,#, HE Yan1,#, GUO Dan1, LV Xiang-guang1,2, HUANG Qi-na1, WU Jian-li1,*
(1State Key Laboratory of Rice Biology, China National Rice Research Institute, Hangzhou 310006, China;2Institute of Crop Sciences, Chinese Academy of Agricultural Sciences, Beijing 100081, China;#These authors contributed equally to this work;*Corresponding author, E-mail: beshangd@163.com)
The pale green leaf mutantHM133 was identified from an EMS-induced IR64 mutant bank. The contents of photosynthetic pigments including chlorophyll and carotenoid ofHM133 were reduced significantly at 6 weeks and 15 weeks after sowing when compared with IR64.The net photosynthetic rate ofHM133 was considerably lower than that of the wild-type IR64 at heading stage while the stomatal conductance was apparently increased. The agronomic traits including plant height, number of filled grain per panicle and seed-setting rate decreased significantly in the mutant compared with the wild-type. In addition, the mutant exhibited a less number of grana, irregular arrangement of thylakoid layer in the chloroplast at the tillering stage. Genetic and mapping analysis showed that the pale green phenotype was controlled by a single recessive gene located in the long arm of chromosome 3 between SSR markers RM143 and RM3684. The interval contains an ORFOsChlDencoding magnesium-chelatase D subunit. Sequence analysis revealed that the mutant allele carried a nucleotide substitution from G to A in the tenth exon ofOsChlD, which led to the substitution of glutamic acid for arginine acid. Therefore, it is deduced thatOsChlDis the candidate gene controlling the pale green leaf phenotype ofHM133.
Oryzasativa; pale green leaf; gene mapping; magnesium-chelatase; photosynthesis
2015-12-29; 修改稿收到日期: 2016-03-12。
國家863計劃資助項目(2014AA10A603); 浙江省超級稻研究重點實驗室項目(2013E10021)。
Q343.5; S511.032
A
1001-7216(2016)06-0603-08
施勇烽, 賀彥, 郭丹, 等. 水稻淡綠葉突變體HM133的遺傳分析與基因定位. 中國水稻科學, 2016, 30(6): 603-610.
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
