利用卡氏白和尼羅紅染色觀察稻瘟病菌有性世代的結構
2016-12-07 07:50:56顧卓侃李玲王教瑜柴榮耀王艷麗張震毛雪琴邱海萍孫國昌
中國水稻科學 2016年6期
顧卓侃 李玲 王教瑜 柴榮耀 王艷麗 張震 毛雪琴 邱海萍 孫國昌
(1杭州師范大學 生命與環(huán)境科學學院, 杭州 310036;2浙江省農業(yè)科學院 植物保護與微生物研究所, 杭州 310021;3浙江農林大學 農業(yè)與食品科學學院, 浙江 臨安 311300; *通訊聯系人, E-mail: wangjiaoyu78@sina.com; sungc01@sina.com)
?
利用卡氏白和尼羅紅染色觀察稻瘟病菌有性世代的結構
顧卓侃1,2李玲2,3王教瑜2,*柴榮耀2王艷麗2張震2毛雪琴2邱海萍2孫國昌2,*
(1杭州師范大學 生命與環(huán)境科學學院, 杭州 310036;2浙江省農業(yè)科學院 植物保護與微生物研究所, 杭州 310021;3浙江農林大學 農業(yè)與食品科學學院, 浙江 臨安 311300;*通訊聯系人, E-mail: wangjiaoyu78@sina.com; sungc01@sina.com)
GU Zhuokan, LI Ling, WANG Jiaoyu, et al. Observation of sexual structure ofMagnaportheoryzaevia Calcofluor white and Nile red staining. Chin J Rice Sci, 2016, 30(6): 668-672.
稻瘟病菌(Magnaportheoryzae)是異宗配合的子囊菌,但至目前,關于其有性世代產生過程和結構的研究相對較少。本研究利用兩個稻瘟病菌菌株Guy-11與2539在多種培養(yǎng)基上進行雜交試驗,觀察有性世代產生情況。結果表明,兩菌株在所有參試培養(yǎng)基上雜交后均能產生子囊殼,但子囊殼的數量、產生速度和成熟度各不相同,以燕麥培養(yǎng)基為最佳。為了進一步觀察有性世代的結構,我們采用卡氏白和尼羅紅對子囊和子囊孢子進行染色和熒光觀察。熒光顯微鏡下,子囊和子囊孢子的細胞壁均能被卡氏白染成清晰的亮藍色,細胞結構清晰可辨。成熟的子囊殼內可產生大量的子囊,子囊中含有8個子囊孢子,子囊孢子通常含有4個細胞。同時,子囊孢子能夠被尼羅紅染成橘紅色,表明子囊孢子中儲藏大量的脂肪類物質。本研究提供了一種有效的觀察稻瘟病菌有性世代結構的熒光染色方法,為后續(xù)的研究奠定了基礎。
稻瘟病菌; 有性生殖; 卡氏白; 尼羅紅;
稻瘟病是世界各稻區(qū)危害最為嚴重的病害之一,其病原菌為稻瘟病菌(Magnaportheoryzae)[1]。稻瘟病菌是一種異宗配合的子囊菌,能夠寄生多種禾本科作物及雜草[2,3]。1971年,Hebert首次報道馬唐瘟菌能夠在人工培養(yǎng)基上獲得有性世代[4]。其后,國內外學者對稻瘟病菌及草瘟病菌做了大量的研究[5]。1976年,Kato等[6]報道了稻瘟病菌能在培養(yǎng)基上形成有性世代,其后各國學者也對此做了相關實驗及證實,證明不同的稻瘟病菌菌株產生有性世代的能力差異極大,不同的環(huán)境條件能顯著影響子囊殼的產生[7-10]。但至目前,在自然環(huán)境下仍未發(fā)現稻瘟病菌有性世代。因此,與稻瘟病菌無性階段和侵染機制的研究相比,關于其有性世代產生過程和結構的觀察相對較少。
利用特異的熒光染料和顯微觀察研究病菌的結構,并明確其所含成分,在稻瘟病菌侵染機制的研究中有較多的應用。卡氏白(Calcofluor White)能夠結合真菌細胞壁中的纖維素和幾丁質,在熒光顯微鏡下能夠觀察到藍色熒光[11]。尼羅紅(Nile Red)是一類苯吩啞嗪酮類化合物, 可以與脂類物質結合并發(fā)出橙紅色熒光[12,13]。尼羅紅發(fā)出的熒光能夠于水中迅速發(fā)生猝滅,而多余的染料不需要清除,這相較于蘇丹黑、尼羅藍-A等染料可以更準確地區(qū)分細胞內的脂類物質和其他貯藏物[14],因而常被用于檢測動物及微生物細胞內的油脂含量。卡氏白和尼羅紅染色在比較稻瘟病菌細胞壁強度和分生孢子細胞內脂類轉移過程中已有應用[15,16],但利用這兩種物質對有性階段進行染色觀察,尚未見報道。
本研究利用兩個稻瘟病菌菌株,觀察了在多種培養(yǎng)基上的有性世代產生情況,并利用卡氏白和尼羅紅染色對子囊和子囊孢子結構進行了熒光觀察,研究結果為進一步研究稻瘟病菌有性世代產生的條件和產生過程提供了基礎和方法。
1 材料與方法
1.1 供試菌株
稻瘟病菌菌株Guy-11、2539作為供試菌株。
1.2 培養(yǎng)基
參試培養(yǎng)基包括完全培養(yǎng)基(Complete medium,CM)[17]、燕麥培養(yǎng)基(Oatmeal agar medium,OMA)[18]、馬鈴薯葡萄糖培養(yǎng)基(Potato Dextrose Agar,PDA,馬鈴薯200 g/L,葡萄糖20 g/L)、淀粉酵母培養(yǎng)基(Starch yeast medium,Starch,酵母粉2 g/L,可溶性淀粉10 g/L,瓊脂粉15 g/L)、基本培養(yǎng)基(Minimal medium,MM)[17]、V8培養(yǎng)基(V8 medium,V8)[19],均按已報道的方法進行配制。
1.3 對峙培養(yǎng)方法
從完全培養(yǎng)基[17]上培養(yǎng)生長5d的菌株菌落邊緣切取2 mm×2 mm菌絲塊,置于5種不同培養(yǎng)基平板上對峙培養(yǎng),在24 h黑暗環(huán)境下27℃下生長7d至菌落相互接觸,然后將各處理移至22℃恒溫光照培養(yǎng)箱繼續(xù)培養(yǎng)。
1.4 調查記錄
對峙培養(yǎng)24 d后,通過奧利巴斯SZX10體式鏡(東京,日本)觀測并拍攝各種類型的培養(yǎng)基上子囊殼的形成情況和形態(tài)特征。
1.5 形態(tài)結構觀察
將在燕麥培養(yǎng)基上生長28d后的Guy-11與2539雜交產生的子囊殼用牙簽挑至載玻片上,覆上蓋玻片,用牙簽輕輕敲擊蓋玻片,壓碎子囊殼,釋放出子囊和子囊孢子,分別使用50 μg/mL的卡氏白(Calcofluor white,Fluorescent Brightener,Sigma)和尼羅紅(Nile Red)染色,在奧林巴斯Xa21熒光顯微鏡下(東京,日本)觀察子囊殼、子囊及子囊孢子結構。
2 結果與分析
2.1 不同培養(yǎng)基上稻瘟病菌有性世代形成情況
對峙培養(yǎng)24 d后,在燕麥培養(yǎng)基和馬鈴薯培養(yǎng)基上,產生大量子囊殼(圖1-A)。成熟的子囊殼具有長長的喙(圖1-B),喙突出菌落表面,形成一個個肉眼可見的小黑點。無數的小黑點,密密麻麻,在菌落交界處形成一條子囊殼帶。同時,馬鈴薯葡萄糖培養(yǎng)基上還存在著一些正在生長或尚未成熟的子囊殼,這些子囊殼的喙非常短小,剛剛突出菌落表面或仍埋在培養(yǎng)基內。淀粉酵母培養(yǎng)基上只能形成極少量成熟的子囊殼,多數的子囊殼埋在培養(yǎng)基內,通過培養(yǎng)基背部給光能夠看到。V8培養(yǎng)基與基本培養(yǎng)基上無法形成成熟的子囊殼,但在背光照射下仍能發(fā)現培養(yǎng)基內發(fā)育不成熟的子囊殼(圖1-C)。計數統計表明,5種培養(yǎng)基中,以燕麥片培養(yǎng)基上產生的子囊殼數量最多(圖1-C),并且最早可見(14~15 d)。
2.2 子囊殼結構的卡氏白染色觀察
使用牙簽隔著蓋玻片輕輕敲打,可壓碎子囊殼。大量的子囊和部分的孢子會從破裂的子囊殼中游離出來。利用卡氏白能夠與細胞壁結合的特性,對碾碎的子囊殼進行染色,能夠清晰地辨別子囊與子囊孢子的輪廓結構(圖-2)。一個子囊通常含有8個子囊孢子,緊密排列在子囊內部。子囊孢子呈鈍菱形,通常有3個隔膜,4個細胞,與稻瘟病菌分生孢子的結構有著明顯的差異。仔細比較熒光亮度可以發(fā)現,子囊孢子細胞壁的亮度明顯高于子囊殼和子囊壁,表明子囊孢子細胞壁中的纖維素和幾丁質含量最高。
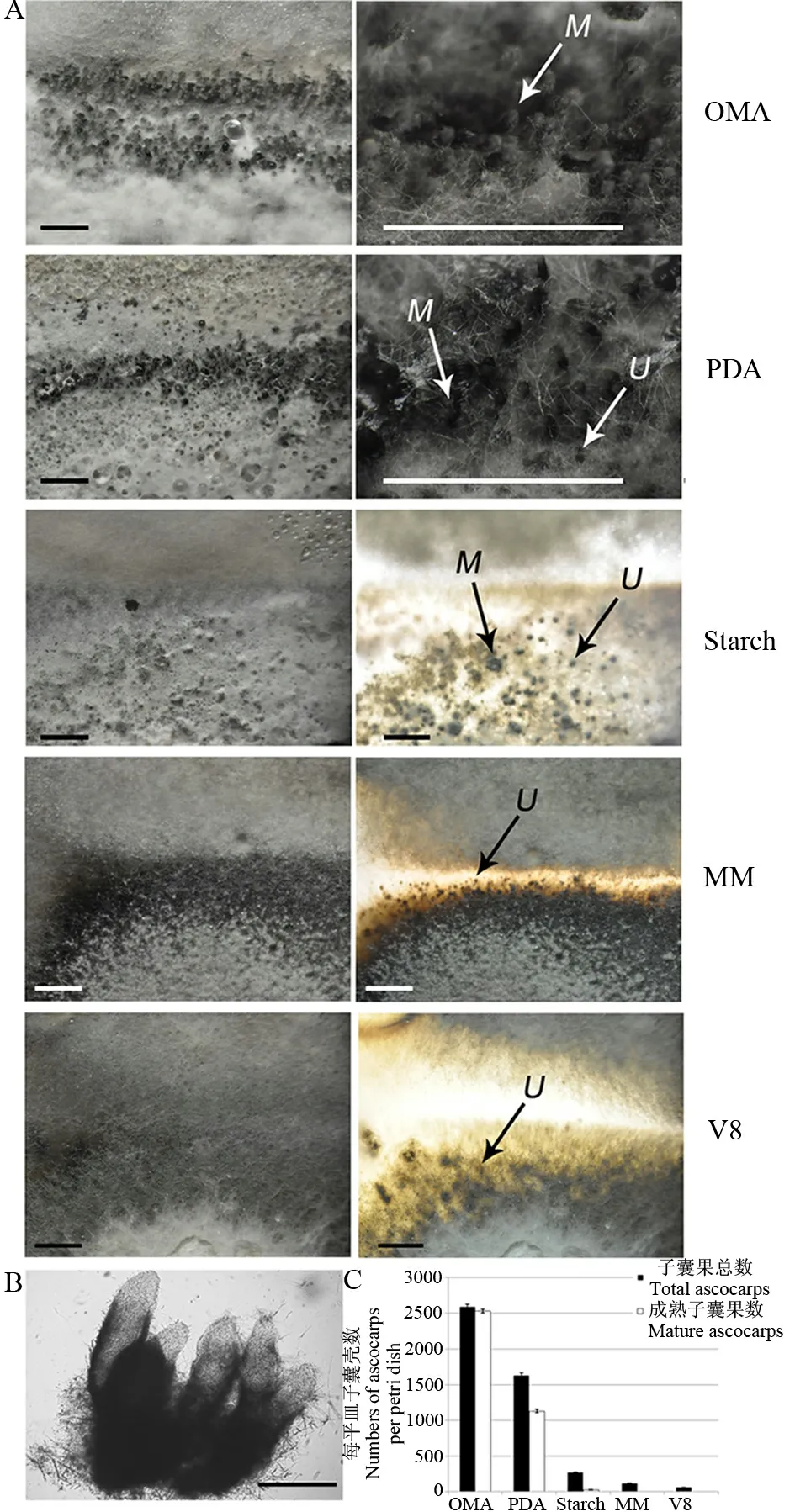
A-燕麥培養(yǎng)基(OMA)、馬鈴薯葡萄糖培養(yǎng)基(PDA)、淀粉酵母培養(yǎng)基(Starch)、基本培養(yǎng)基(MM)、V8培養(yǎng)基(V8)子囊殼形成情況;M,成熟的子囊殼;U,不成熟的子囊殼;標尺=1000 μm。B-顯微鏡下成熟子囊殼的形態(tài);標尺=100 μm。C-各培養(yǎng)基上子囊殼的形成數量統計。
A, Perithecia formed on oatmeal agar (OMA), potato dextrose agar (PDA), starch yeast medium (Starch), minimal medium (MM) and V8 medium (V8). M, Mature perithecia; U, Unmatured perithecia. Bars=1000 μm. B, Microscopical analysis of morphology of perithecia. Bars=100 μm. C, Number of total and mature perithecia formed on different media.
圖1 各培養(yǎng)基上子囊殼的形成情況
Fig. 1. Perithecia formed on different media.
2.3 子囊與子囊孢子的尼羅紅染色
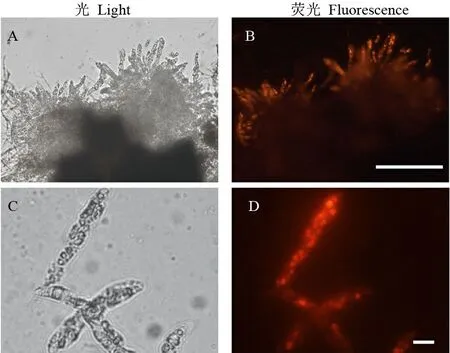
尼羅紅能特異地結合真菌細胞中的脂類物質,熒光顯微鏡下呈現橙紅色熒光[15]。利用尼羅紅溶液對稻瘟病病菌的子囊殼、子囊及子囊孢子進行染色(圖3),顯微鏡下可見,子囊孢子內部被染成明亮的橙紅色,而子囊殼、子囊、子囊與孢子的間隙以及子囊孢子的細胞壁幾乎不能被染色,表明子囊孢子的內部富含脂類物質,這點與無性孢子類似。

A~D-大量游離的子囊,標尺=100 μm; E-一個完整的子囊,標尺=10 μm; H-放大的子囊孢子,標尺=10 μm。
A-D, Asci migrated from a perithecium, Bars=100 μm; E and F, An intact ascus, bar=10 μm; G and H, Magnified image of an ascospore, bar = 10 μm.
圖2 子囊與子囊孢子的卡氏白染色觀察
Fig. 2. Structure of perithecia stained with Calcofluor white (Fluorescent Brightener) under a fluorescent microscopy.
3 小結與討論
我們的實驗結果表明,燕麥片培養(yǎng)基上更有利于稻瘟病菌有性世代的產生,此前也有相關的報道[3]。然而在其他參試培養(yǎng)基上都有初期的子囊殼產生,只是無法充分成熟或發(fā)育極慢。這表明稻瘟病菌有性世代的啟動是由菌株本身的特性(交配型)決定的,但不同的營養(yǎng)成分可以極大地干擾其有性世代的發(fā)育和是否能產生可育的后代。自然條件下,很難發(fā)現有性生殖的存在[4]。在寄主葉片和莖干等發(fā)病部位,未見發(fā)現有性世代的報道,很可能是因為這些部位的營養(yǎng)物質無法滿足有性世代形成對營養(yǎng)的需求。燕麥培養(yǎng)基上子囊殼發(fā)育良好,或許暗示著在水稻或其他禾本科植物籽粒上,有形成可育的有性后代的可能。
A~B-標尺=100 μm; C~D-標尺=10 μm。
A and B, Bar =100 μm; C and D, bar =10 μm.
圖3 子囊與子囊孢子的尼羅紅染色
Fig. 3. Perithecia and ascospores stained with Nile red.
子囊殼破碎后能夠分離出大量的子囊,只憑借光學顯微鏡進行觀察,很難清楚分辨其中子囊孢子的個數、排列情況和細胞結構。借助卡氏白對細胞壁的染色,通過熒光顯微鏡觀察,則有效地屏蔽了子囊和孢子內容物以及雜質的干擾,從而清楚地分辨子囊和子囊孢子的細胞結構。為進一步分析不同菌株有性世代的發(fā)育狀況和分離有性生殖相關突變體奠定了基礎。同時,研究結果也表明,子囊孢子的細胞壁成分與分生孢子類似,均含有纖維素和幾丁質等主要成分。尼羅紅是脂肪的特異性染料,子囊孢子細胞可以被染上明亮的橙紅色,表明細胞內富含脂質。這與分生孢子的情況類似,分生孢子內貯藏的大量脂肪在萌發(fā)過程中迅速降解,提供病菌致病過程的營養(yǎng)與能量[15]。稻瘟病菌的子囊孢子同樣含有大量脂類物質,應該可以滿足萌發(fā)和致病的需要,具有潛在的致病能力。而有關子囊孢子的萌發(fā)的詳細過程、能否形成典型侵染結構及其致病能力等,仍需進一步研究。
對比有利于有性世代形成的實驗室條件,我國南方稻區(qū)晚稻生育后期的溫度與光照適合稻瘟病菌有性世代的形成。也有部分研究者根據分子遺傳學上的證據推測,稻瘟病菌在東南亞等稻瘟病菌發(fā)源地,存在自然的有性生殖過程[20]。但即便如此,直接證據卻一直很難發(fā)現,至今尚未在自然條件下發(fā)現稻瘟病菌的子囊殼。因此,明確稻瘟病菌有性世代產生的條件、過程和結構特點,將有助于在自然界中探尋病菌有性過程,完善病菌生活史,進一步明確病菌進化與變異,為病害流行測報和防控策略的制定提供理論依據。
[1] Barr M E. Magnaporthe, Telimenella, and Hyponectria (Physosporellaceae).Mycologia,1997: 952-966.
[2] Zeigler R S. Recombination inMagnaporthegrisea.AnnRevPhytopathol, 1998,36: 249-275.
[3] 沈瑛, 金敏忠. 我國稻瘟病菌有性態(tài)的研究. 中國農業(yè)科學, 1994,27: 25-29.
[4] Hebert T. The perfect stage ofPyriculariagrisea.Phytopathology, 1971,61: 83-87.
[5] Yaegashi H, Yamada M. Pathogenic race and mating type ofPyriculariaoryzaefrom Soviet Union, China, Nepal, Thailand, Indonesia and Colombia.JpnJPhytopathol, 1986, 52: 225-234.
[6] Kato H, Yamaguchi T, Nishihara N. The perfect state ofPyriculariaoryzaeCav. in culture.AnnPhytopatholSocJapan, 1976, 42: 507-510.
[7] Kolmer J A, Ellingboe A H. Genetic relationships between fertility and pathogenicity and virulence to rice inMagnaporthegrisea.CanJBot, 1988,66: 891-897.
[8] Levy M, Romao J, Marchetti M, et al. DNA fingerprinting resolves pathotype diversity in a plant pathogenic fungus.PlantCell, 1991, 3: 95-102.
[9] Silue D, Notteghem J. Production of perithecia ofMagnaporthegriseaon rice plants.MycolRes,1990, 94: 1151-1152.
[10]Valent B, Crawford M S, Weaver C G, et al. Genetic studies of fertility and pathogenicity inMagnaporthegrisea(Pyriculariaoryzae).IowaStateJRes, 1986, 60: 569-594.
[11]Nagata T, Takebe I. Cell wall regeneration and cell division in isolated tobacco mesophyll protoplasts.Planta, 1970,92: 301-308.
[12]Lambert M, Moss C W. Comparison of the effects of acid and base hydrolyses on hydroxy and cyclopropane fatty acids in bacteria.JClinMicrobiol, 1983,18: 1370-1377.
[13]Diaz G, Melis M, Batetta B, et al. Hydrophobic characterization of intracellular lipids in situ by Nile Red red/yellow emission ratio.Micron, 2008, 39: 819-824.
[14]Kranz R G, Gabbert K K, Madigan M T. Positive selection systems for discovery of novel polyester biosynthesis genes based on fatty acid detoxification.ApplEnvironMicrobiol, 1997, 63: 3010-3013.
[15]Wang J, Zhang Z, Wang Y, et al. PTS1 peroxisomal import pathway plays shared and distinct roles to PTS2 pathway in development and pathogenicity ofMagnaportheoryzae.PloSOne, 2013, 8: e55554.
[16]Li L, Wang J, Zhang Z, et al.MoPEX19, which is essential for maintenance of peroxisomal structure and woronin bodies, is required for metabolism and development in the rice blast fungus.PloSOne, 2014, 9: e85252.
[17]Talbot N J, Ebbole D J, Hamer J E. Identification and characterization ofMPG1, a gene involved in pathogenicity from the rice blast fungusMagnaporthegrisea.PlantCell, 1993,5: 1575-1590.
[18]Yi M, Park J H, Ahn J H, et al.MoSNF1 regulates sporulation and pathogenicity in the rice blast fungusMagnaportheoryzae.FungalGenetBiol, 2008, 45: 1172-1181.
[19]Kent C R, Ortiz-Bermudez P, Giles S S, et al. Formulation of a defined V8 medium for induction of sexual development ofCryptococcusneoformans.ApplEnvironMicrobiol, 2008, 74: 6248-6253.
[20]Saleh D, Xu P, Shen Y, et al. Sex at the origin: An asian population of the rice blast fungusMagnaportheoryzaereproduces sexually.MolEcol, 2012,21: 1330-1344.
Observation of Sexual Structure ofMagnaportheoryzaevia Calcofluor White and Nile Red Staining
GU Zhuo-kan1, 2, LI Ling2, 3, WANG Jiao-yu2,*, CHAI Rong-yao2, WANG Yan-li2, ZHANG Zhen2,MAO Xue-qin2, QIU Hai-ping2, SUN Guo-chang2,*
(1School of Life and Environmental Science, Hangzhou Normal University, Hangzhou 310036, China;2Institute of Plant Protection and Microbiology, Zhejiang Academy of Agricultural Sciences, Hangzhou 310021, China;3College of Agriculture and Food Science, Zhejiang A&F University, Ling′an 311300, China;*Corresponding authors, E-mail: wangjiaoyu78@sina.com; sungc01@sina.com)
Magnaportheoryzaeis an economically important heterothallic ascomycete which causes rice blast, the most destructive rice disease worldwide. To the present, sexual generation of the fungus has been relatively less investigated. In the present work, Guy-11 and 2539, twoM.oryzaestrains in opposite mating types, were cross cultured on six deferent substrates to induce the sexual generation. The strains produced ascocarps on all of the substrates tested, however, the number, forming time and ripeness degree of the ascocarps varied largely with the producing substrates, among which oatmeal agar(OMA) gave the optimum value. In order to better detect the structures of sexual generation, the asci and ascospores were stained with Fluorescent Brightener and Nile red, and examined using a fluorescence microscopy. Under the fluorescence microscope, the cell walls were stained bright blue by Fluorescent Brightener, which greatly improved the visibility of asci and ascospores. Numbers of asci were found in mature ascocarps, with eight ascospores in each ascus, and an ascospore usually has four cells. Meanwhile, the ascospores can be stained bright orangey-red by Nile red, indicating they are rich in lipids. These findings showed that the fluorescence staining is an effective method to observe the structure of sexual generation inM.oryzae.
Magnaportheoryzae; sexual generation; Calcofluor White; Nile Red
2016-03-03; 修改稿收到日期: 2016-07-27。
國家自然科學基金資助項目(31170136, 31470249); 浙江省公益技術研究農業(yè)項目(2013C32019)。
S435.111.4+1
A
1001-7216(2016)06-0668-05
顧卓侃, 李玲, 王教瑜, 等. 利用卡氏白和尼羅紅染色觀察稻瘟病菌有性世代的結構. 中國水稻科學, 2016, 30(6): 668-672.
