補體C3d-p28增強阿爾茨海默病DNA疫苗Th2型免疫反應的研究
2016-12-22 01:55:58郭婉姝邢曉娜曹云鵬
中風與神經疾病雜志 2016年11期
關鍵詞:血清
郭婉姝, 邢曉娜, 王 楠, 曹云鵬
?
補體C3d-p28增強阿爾茨海默病DNA疫苗Th2型免疫反應的研究
郭婉姝1, 邢曉娜1, 王 楠1, 曹云鵬2
目的 探討補體C3d-p28作為分子佐劑,在阿爾茨海默病DNA疫苗免疫反應中的作用。方法 分別將重組質粒p(Aβ3-10)10,p(Aβ3-10)10- C3d-p28.3和 空載體pcDNA3.1(+)用肌肉注射的方法免疫8~10周齡的雌性BALB/c鼠。質粒注射前24 h,布比卡因肌肉注射誘導輕微的肌肉變性。應用ELISA方法檢測血清抗Aβ抗體的滴度、抗體分型、體外脾細胞培養上清液中IL-4 和 IFN-γ的含量。免疫組織化學染色法檢測免疫血清與轉基因鼠腦內Aβ斑的結合能力。結果 重組質粒疫苗p (Aβ3-10)10僅誘導出低滴度的抗Aβ抗體,產生了Th1/Th2混合型的免疫反應。而重組質粒疫苗p (Aβ3-10)10- C3d-p28.3誘導出較高滴度的抗Aβ抗體,體外脾細胞培養上清液中IFN-γ低和IL-4高,即引起了Th2型細胞免疫反應,同時產生的抗Aβ抗體能夠與雙轉基因鼠APP/PS1腦中沉積的Aβ斑塊結合。結論 補體C3d-p28分子佐劑能夠增強抗Aβ抗體的產生并且誘發Th2型的免疫反應。
阿爾茨海默病; β淀粉樣蛋白; DNA疫苗; C3d-p28分子佐劑
Aβ的基因免疫治療,阻止Aβ產生、聚集和(或)增加Aβ清除被認為是治療阿爾茨海默病的有效策略。一個理想的Aβ疫苗,不僅要刺激機體產生能夠起治療作用的抗Aβ抗體,還要避免不恰當的Th1型免疫反應引起的腦膜腦炎等副作用。引入恰當的基因佐劑可以增加抗原提呈和機體對抗原的免疫應答能力,把免疫應答導向Th2 方向。補體C3d能夠促進抗體的產生及其親和力的成熟[1],誘發更強烈的體液免疫反應[2]。而且C3d分子可降調節免疫活性細胞Th1型細胞因子的表達水平,升調節Th2型細胞因子的表達[3]。補體C3d分子與CR2特異性結合的結合域位于C3d分子1209-1236aa區域(C3d-p28),p28分子與全長C3d分子有相似的佐劑性質[4],但p28分子大小僅占C3d分子全長的9%[5],這更增強了DNA疫苗的有效利用。本研究旨在探討補體C3d-p28作為分子佐劑,對阿爾茨海默病DNA疫苗免疫反應的影響。
1 材料與方法
1.1 實驗動物及材料
1.1.1 動物 18只、8~10周齡、雌性BALB/c鼠購于中國醫科大學實驗動物研究中心。
1.1.2 主要材料 重組質粒疫苗p(Aβ3-10)10和重組質粒疫苗p(Aβ3-10)10-C3d-p28.3由湖南長沙贏潤生物技術有限公司合成。抗Aβ抗體6E10 (Signet,美國);羊抗鼠IgG1-HRP (Zymed,美國);羊抗鼠IgG2a-HRP (Zymed,美國);羊抗鼠IgG2b-HRP (Zymed,美國);小鼠白介素-4 ELISA試劑盒(R&D Systems China Co.Ltd.,上海);小鼠IFN-γ ELISA試劑盒(R&D Systems China Co.Ltd.,上海)。
1.2 方法
1.2.1 動物分組 8~10周齡雌性BALB/c鼠共18只,按隨機數字表法分成3組,每組6只。第一組:重組質粒p(Aβ3-10)10組;第二組:重組質粒p(Aβ3-10)10-C3d-p28.3組;第三組:空載體pcDNA3.1(+)組。
1.2.2 免疫方法 采用肌肉注射法。注射部位:小鼠左后腿股四頭肌。重組質粒p(Aβ3-10)10組和重組質粒p(Aβ3-10)10-C3d-p28.3組:注射疫苗抗原濃度為1.0 μg/ μl,每只鼠注射體積為100 μl,一次注射疫苗100 μg。空載體pcDNA3.1(+)組:注射疫苗抗原濃度為1.0 μg/ μl,每只鼠注射體積為100 μl,一次注射疫苗100 μg。每次注射以上兩組重組質粒疫苗前1 d注射部位用布比卡因進行預處理:布比卡因配制濃度5 mg/ml,一只小鼠一次注射50 μl(250 μg)。三組小鼠疫苗注射前均需腹腔注射10%水合氯醛(0.03 mg/kg)進行麻醉。三組小鼠共免疫5次,每2 w免疫一次。
1.2.3 取血及血清樣本處理 疫苗接種前、從第2次注射開始每次接種后1 w取鼠眶靜脈血0.5 ml。采血后,血液置于37 ℃ 3~5 h,4 ℃下5000×g離心10 min,吸取上清,分裝-70 ℃凍存。
1.2.4 間接ELISA法檢測血清中抗Aβ抗體滴度及分型 (1)抗原包被96孔板,每孔加入100 μl 稀釋的Aβ42(5 μg/ml),4oC孵育過夜;(2)用含有0.05%TW20的1×PBS(200 μl每孔)洗板3次,下同;(3)每孔中加入200 μl封閉緩沖液,室溫下孵育1 h;(4)免疫前和免疫后的血清以1∶1000(1 μl加入1 ml封閉緩沖液)稀釋,用封閉緩沖液封閉1 h后,洗板;(5)孔中加入50 μl稀釋的血清和標準抗體,然后室溫孵育3 h,洗板;(6)加入稀釋的二抗(IgG:1:2000;IgG1:1:2000,IgG2a:1:1000,IgG2b:1∶1000),50 μl每孔;室溫下孵育1 h,洗板;(7)每孔中加入100 μl TMB,室溫下孵育15 min,或者是到顏色產生;(8)每孔中加入100 μl終止液(2N H2SO4);(9)讀取450 nm處的OD值。
1.2.5 ELISA法檢測脾細胞培養上清液中細胞因子IL-4和IFN-γ的含量 步驟按試劑盒說明書進行。
1.2.6 免疫組織化學染色方法檢測免疫血清與APP/PS1雙轉基因鼠腦內Aβ斑結合情況 (1)石蠟切片(APP/PS1雙轉基因鼠腦組織切片,4 μm),60 ℃烤箱烤片1 h,切片常規脫蠟至水,;(2)30%H2O21份加蒸餾水10份混合,室溫5~10 min以滅活內源性酶。蒸餾水洗3次。(3)熱修復抗原。將切片進入0.01 mol/L枸櫞酸鹽緩沖液(pH 6.0),微波爐加熱至沸騰。后斷電,間隔5~10 min后,反復2次。冷卻后PBS洗滌2次;(4)滴加正常山羊血清封閉液,室溫20 min;(5)滴加末次接種后的鼠血清,37 ℃ 1 h,PBS洗2 min×3次;(6)滴加生物素化山羊抗小鼠IgG,37 ℃ 20 min,PBS洗2 min×3次;(7)滴加試劑SABC,37 ℃ 20 min,PBS洗5 min×4次;(8)DAB顯色:使用DAB顯色試劑盒。取1 ml蒸餾水,加試劑盒中A,B,C試劑各1滴,混勻后加至切片。室溫顯色,鏡下控制反應時間,一般在5~30 min之間;(9)脫水、透明、封片、顯微鏡觀察。
2 結 果
2.1 血清抗Aβ抗體滴度及分型的檢測 經重組質粒p(Aβ3-10)10和重組質粒p(Aβ3-10)10-C3d-p28.3免疫的小鼠均產生了抗Aβ抗體,并且在第二次免疫后血清抗Aβ抗體的滴度平穩增長,重組質粒p(Aβ3-10)10從最初的5.09±1.42 μg/ml增長到:(26.11±8.55) μg/ml;重組質粒p(Aβ3-10)10-C3d-p28.3組從最初的(10.46±4.35) μg/ml增長到(57.12±12.08) μg/ml。重組質粒p(Aβ3-10)10-C3d-p28.3組的抗Aβ抗體的滴度高于重組質粒p(Aβ3-10)10組(P<0.05),但是pcDNA3.1(+)免疫的小鼠未檢測出抗Aβ抗體(見圖1A)。
抗IgG1 抗體檢測:重組質粒p(Aβ3-10)10- C3d-p28.3組(32.25±6.78) μg/ml較重組質粒p(Aβ3-10)10組(10.33±3.01) μg/ml明顯增高(P<0.05),但是抗IgG2a和IgG2b抗體檢測:重組質粒p(Aβ3-10)10- C3d-p28.3組(3.28 ± 1.54) μg/ml、(6.03± 2.17) μg/ml低于重組質粒p(Aβ3-10)10組(7.02 ±2.56) μg/ml和IgG2b抗體(12.62± 4.73) μg/ml(見圖1B)。
重組質粒p(Aβ3-10)10- C3d-p28.3組的IgG1/IgG2a的比值(9.83 ±1.18)明顯高于重組質粒p(Aβ3-10)10組(1.47 ±0.04)(P<0.05,圖1C)。


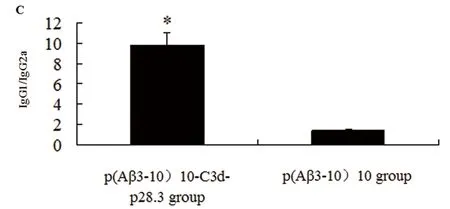
A:經重組質粒p(Aβ3-10)10、重組質粒p(Aβ3-10)10-C3d-p28.3和空載體pcDNA3.1(+)免疫后小鼠的血清抗Aβ抗體滴度。B:重組質粒p(Aβ3-10)10和重組質粒p(Aβ3-10)10-C3d-p28.3組抗Aβ抗體的分型:IgG1,IgG2a,IgG2b的含量。C:重組質粒p(Aβ3-10)10和重組質粒p(Aβ3-10)10-C3d-p28.3組的IgG1/IgG2a的比值。*P<0.05,與重組質粒p(Aβ3-10)10組相比
圖1 抗Aβ抗體的滴度和分型分析


A:在相應的自身抗原刺激下的脾細胞培養上清液中IL-4的含量測定。*P<0.05,與空載體pcDNA3.1(+)組和重組質粒p(Aβ3-10)10組相比。B:在相應的自身抗原刺激下的脾細胞培養上清液中IFN-γ的含量測定。*P<0.05,與重組質粒p(Aβ3-10)10-C3d-p28.3組和空載體pcDNA3.1(+)組相比
圖2 脾細胞培養上清液中IL-4、IFN-γ的檢測


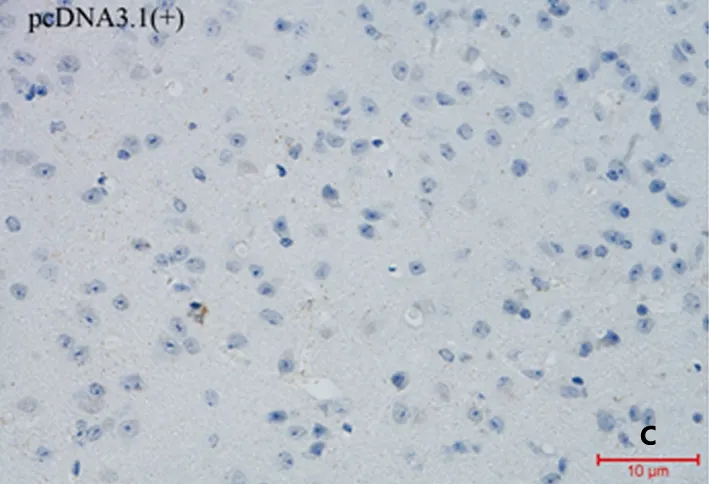
A:重組質粒p(Aβ3-10)10免疫后的血清能夠與12月齡的APP/PS1雙轉基因鼠腦內Aβ斑結合。B:重組質粒p(Aβ3-10)10-C3d-p28.3免疫后的血清能夠與Aβ斑結合。C:空載體pcDNA3.1(+)免疫后的血清不能與Aβ斑結合
圖3 免疫血清與APP/PS1雙轉基因鼠腦內Aβ斑結合能力的檢測
2.2 脾細胞培養上清液中細胞因子的檢測 脾細胞培養上清液中的IL-4含量:重組質粒p(Aβ3-10)10-C3d-p28.3組(82.26 ±9.94 pg/ml)明顯高于重組質粒p(Aβ3-10)10組(42.33 ± 6.78) pg/ml和pcDNA3.1(+)組(13.74±4.25) pg/ml(P<0.05),而重組質粒p(Aβ3-10)10組略高于pcDNA3.1(+)組(P<0.05,圖2A)。
脾細胞培養上清液中的IFN-γ含量測定:重組質粒p(Aβ3-10)10組(51.2 ±9.67) pg/ml高于重組質粒p(Aβ3-10)10-C3d-p28.3組(29.77±7.76) pg/ml和pcDNA3.1(+)組(23.13±7.16) pg/ml(P<0.05),而后兩組并沒有差異(P>0.05,圖2B)。
2.3 免疫血清與APP/PS1雙轉基因鼠腦內Aβ斑結合能力的檢測 重組質粒p(Aβ3-10)10和重組質粒p(Aβ3-10)10-C3d-p28.3免疫后的血清均能夠與12月齡的APP/PS1雙轉基因鼠的腦內Aβ斑結合(見圖3A、B),但是重組質粒p(Aβ3-10)10結合Aβ斑的能力明顯弱于重組質粒p(Aβ3-10)10-C3d-p28.3。空載體pcDNA3.1(+)免疫后的血清(見圖3C)不能與12月齡的APP/PS1雙轉基因鼠的腦內Aβ斑結合。
3 討 論
研究發現,編碼不同抗原基因(西尼羅病毒、乙肝病毒、豬生殖和呼吸綜合征病毒、瘧原蟲環孢子蛋白)的DNA疫苗在融合了C3d-p28分子佐劑后,抗原的免疫原性大大地增強了,DNA疫苗的有效性也大大提高[5~8]。因此,補體C3d-p28作為分子佐劑應用前景廣闊。
本研究中,空載體pcDNA3.1(+)組沒有抗Aβ抗體的產生。而在第二次免疫后,重組質粒p(Aβ3-10)10-C3d-p28.3組和重組質粒p(Aβ3-10)10組都產生了抗Aβ抗體,不過重組質粒p(Aβ3-10)10-C3d-p28.3組產生的抗Aβ抗體量要明顯高于重組質粒p(Aβ3-10)10組,這說明融合了分子佐劑C3d-p28.3的Aβ疫苗免疫原性明顯強于無佐劑的Aβ疫苗。
重組質粒p(Aβ3-10)10-C3d-p28.3組產生了以IgG1為主的抗體,IgG2a和 IgG2b的值較低,相對應的重組質粒p(Aβ3-10)10組則產生了低水平的IgG1,略高水平的IgG2a和 IgG2b抗體,而且重組質粒p(Aβ3-10)10-C3d-p28.3組IgG1/IgG2a的比值明顯高于重組質粒p(Aβ3-10)10組。據研究證實疫苗免疫后誘導的免疫球蛋白IgG的亞型可以間接反應免疫反應是Th1型還是Th2型。Th1型免疫反應主要產生IgG2a抗體,通過產生IFN-γ,IL-2等細胞因子誘導細胞介導的炎癥反應;而Th2型免疫反應主要誘導IgG1抗體,通過產生IL-4,IL-5等細胞因子增強體液免疫反應[9]。IgG1/IgG2a的比值可以反映出該Aβ疫苗所誘導的主要免疫應答類型。本實驗中,融合了分子佐劑C3d-p28.3的Aβ疫苗產生了以IgG1為主的抗體,高比值的IgG1/IgG2a,這都傾向于Th2型免疫反應;而無佐劑的Aβ疫苗產生了Th1/ Th2混合型的免疫反應。
重組質粒p(Aβ3-10)10-C3d-p28.3疫苗免疫后,經自身抗原刺激,脾細胞培養上清液中IL-4高于重組質粒p(Aβ3-10)10組和空載體pcDNA3.1(+)組,脾細胞培養上清液中IFN-γ水平重組質粒p(Aβ3-10)10組高于重組質粒p(Aβ3-10)10-C3d-p28.3組和空載體pcDNA3.1(+)組,再次說明融合了分子佐劑C3d-p28.3的Aβ疫苗產生了Th2型免疫反應,所以對于AD主動免疫治療,獲得一個主要的Th2型免疫反應比單純提高抗Aβ抗體的滴度更為重要;而且,Th2型的抗體清除Aβ沉積更為有效,發生炎癥反應的可能性更低[10]。
腦內Aβ斑塊沉積是AD診斷的決定性指標,它的清除用來評價AD模型鼠的治療效果。一個12月齡的APP/PS1轉基因鼠腦切片用來檢測免疫血清產生的抗Aβ抗體與腦內Aβ斑塊結合的能力。重組質粒p(Aβ3-10)10-C3d-p28.3疫苗和重組質粒p(Aβ3-10)10疫苗免疫后的血清均能夠與12月齡的APP/PS1轉基因鼠腦切片的Aβ斑塊結合,而空載體pcDNA3.1(+)組的免疫血清不能夠與腦切片的Aβ斑塊結合。這說明了融合了分子佐劑C3d-p28.3的Aβ疫苗對腦內Aβ斑塊產生了特異性的免疫反應,并且這種能力強于無佐劑的Aβ疫苗。
總之,重組質粒疫苗 p(Aβ3-10)10-C3d-p28.3,正因為融合了分子佐劑C3d-p28.3,不僅能夠刺激產生具有治療水平的抗Aβ抗體,還能夠誘發Th2型的免疫反應,避免了炎癥副作用,最重要的是,產生的抗Aβ抗體能夠與APP/PS1雙轉基因鼠腦內的Aβ斑塊結合。分子佐劑C3d-p28.3的作用機制正在研究中。
[1]Yang S,Wang C,Fang X,et al. Fusion of C3d molecule with neutralization epitope(s) of hepatitis E virus enhances antibody avidity maturation and neutralizing activity following DNA immunization K[J]. Virus Res,2010,151(2):162-169.
[2]Li DJ,Wang HM,Zhao XR,et al. Gene fusion of molecular adjuvant C3d to hCGβenhances the anti-hCGβ antibody response in DNA immunization[J]. J Reproduct Immunol,2003,60:129-141.
[3]Movsesyan N,Mkrtichyan M,Petrushina I,et al. DNA epitope vaccine containing complement component C3d enhances anti-amyloid-β antibody production and polarizes the immune response towards a Th2 phenotype[J]. J Neuroimmunol,2008,205:57-63.
[4]Bower JF,Ross TM. A minimum CR2 binding domain of C3d enhances immunity following vaccination[J]. Adv Experiment Med Biol,2006,587:249-264.
[5]Dunn MD,Rossi SL,Carter DM,et al. Enhancement of anti-DIII antibodies by the C3d derivative P28 results in lower viral titers and augments protection in mice[J]. Virology J,2010,7:95.
[6]Wang LX,Xu W,Guan QD,et al. Contribution of C3d-P28 repeats to enhancement of immune responses against HBV-preS2/S induced by gene immunization[J]. World J Gastroenterol,2004,10(14):2072-2077.
[7]Zhang D,Xia Q,Wu J,et al. Construction and immunogenicity of DNA vaccines encoding fusion protein of murine complement C3d-p28 and GP5 gene of porcine reproductive and respiratory syndrome virus[J]. Vaccine,2011,29:629-635.
[8]Bergmann-Leitner ES,Duncan EH,Leitner WW,et al. C3d-defined complement receptor-binding peptide p28 conjugated to circumsporozoite protein provides protection against Plasmodium berghei[J]. Vaccine,2007,25:7732-7736.
[9]Finkelman FD,Holmes J,Katona IM,et al. Lymphokine control of in vivo immunoglobulin isotype selection[J]. Annu Rev Immunol,1990,8:303-333.
[10]Chauhan NB,Siegel GJ. Efficacy of anti-Abeta antibody isotypes used for intracerebroventricular immunization in TgCRND8[J]. Neurosci Lett,2005,375:143-147.
Enhancement of Th2 Immune Response to Alzheimer’s disease DNA Vaccine by complement C3d-p28
GUO Wanshu,XING Xiaona,WANG Nan,et al.
(Department of Neurology,The People’s Hospital of Liaoning Province,Shenyang 110016,China)
Objective To discuss the effect of complement C3d-p28 in the immune response of Alzheimer DNA vaccine bstract. Methods Eight to ten week-old female BALB/c mice were immunized intramuscularly with recombinant plasmid p(Aβ3-10)10,p(Aβ3-10)10-C3d-p28.3 and pcDNA3.1(+). 24 h prior to plasmid injection,bupivacaine was injected to induce mild muscle degeneration. ELISA was used to detect the titer of serum anti-Aβ antibody,isotypes of immunoglobulin and levels of IL-4 and IFN-γ in ex vivo splenocyte culture supernatants. Immunohistochemistry was used to detect the binding capability of antisera to Aβ plaques in an APP/PS1 transgenic mouse brain. Results We have successfully constructed two plasmid DNA vaccines:p(Aβ3-10)10 expressing ten repeats of Aβ3-10 without adjuvant and p(Aβ3-10)10- C3d-p28.3 encoding ten repeats of Aβ3-10 and three copies of C3d-p28 as a molecular adjuvant. Immunization of eight to ten week-old female BALB/c mice with p(Aβ3-10)10 vaccine induced only moderate titers of anti-amyloid-β antibodies and elicited a Th1/ Th2 immune response. However,adding of the C3d-p28 molecular adjuvant to Aβ3-10 DNA vaccine could generate high levels of anti-amyloid-β antibodies (P<0.05) which bound to Aβ plaques in APP/PS1 transgenic mouse brain tissue. More importantly,the vaccine elicited a predominantly IgG1 humoral response (P<0.05),low levels of IFN-γ (P<0.05) and high levels of IL-4 in ex vivo cultured splenocytes (P<0.05),indicating a Th2-polarized cellular immune response. Conclusion Complement C3d-p28 molecular adjuvant enhances anti-amyloid-β antibody generation and induces a Th2-polarized immune response.
Alzheimer’s disease; Amyloid-β; DNA vaccine; C3d-p28 molecular adjuvant
1003-2754(2016)11-0976-04
2016-07-28;
2016-10-06
(1.遼寧省人民醫院神經內科,遼寧 沈陽 110016;2.中國醫科大學附屬第一醫院神經內科,遼寧 沈陽 110001)
曹云鵬,E-mail:ypengcao@yahoo.com
R749.1+6
A
猜你喜歡
中老年保健(2021年3期)2021-08-22 06:50:04
天津醫科大學學報(2021年2期)2021-03-29 05:31:08
昆明醫科大學學報(2021年1期)2021-02-07 01:06:36
現代臨床醫學(2021年1期)2021-01-26 00:56:02
昆明醫科大學學報(2020年12期)2021-01-26 00:44:04
中華養生保健(2020年4期)2020-11-16 01:31:40
中西醫結合肝病雜志(2020年2期)2020-10-27 02:18:50
豬業科學(2018年8期)2018-09-28 01:27:38
中成藥(2017年8期)2017-11-22 03:18:47
川北醫學院學報(2015年5期)2015-12-05 08:22:29
