小黑山島海域刺參、魁蚶和紫貽貝生境適宜性分析
2017-02-08 05:58:03唐柳青王其翔劉洪軍張智鵬劉騁躍
生態(tài)學報 2017年2期
唐柳青, 王其翔,劉洪軍,張智鵬,劉騁躍,周 健,*
1 中國海洋大學海洋生命學院,青島 266003 2 山東省海洋生物研究院,青島 266104
小黑山島海域刺參、魁蚶和紫貽貝生境適宜性分析
唐柳青1, 王其翔2,劉洪軍2,張智鵬1,劉騁躍1,周 健2,*
1 中國海洋大學海洋生命學院,青島 266003 2 山東省海洋生物研究院,青島 266104
以小黑山島臨近海域為研究對象,利用生境適宜性指數(shù)(habitat suitability index,HSI)模型選劃適宜刺參(Stichopusjaponicas)、魁蚶(Scapharcabroughtonii)和紫貽貝(Mytilusedulis)增殖修復的區(qū)域。分別針對每個修復物種篩選出7個生境評價因子,結合專家賦值法和層次分析法確定每個評價因子的權重,利用GIS空間分析模塊將現(xiàn)狀調查數(shù)據(jù)進行插值、重分類和柵格計算,繪制研究區(qū)域目標種群生境適宜性地圖。結果表明:對于刺參和紫貽貝,研究區(qū)域均適宜其生長繁殖,同一物種,相同季節(jié)在空間上無站位差異,但各季節(jié)的生境適宜性分區(qū)變化明顯;對于魁蚶來說,東北部海域較適宜增殖,其次為西部海域,四季均以較適宜生境為主,僅冬季出現(xiàn)基本適宜生境。水溫是造成季節(jié)差異的主要因素,底質類型則是引起生境站位差異的重要原因。可為后續(xù)的生物多樣性保育和生態(tài)修復提供基礎資料參考。
生境適宜性指數(shù); 刺參(Stichopusjaponicas); 魁蚶(Scapharcabroughtonii); 紫貽貝(Mytilusedulis); 小黑山島
受人類環(huán)境破壞行為加大的影響,許多生物賴以生存的棲息地遭到大面積破壞,導致自然生物種群出現(xiàn)不同程度的衰退,迫切需要加強對生物生境的保護。生境適宜性指數(shù)(habitat suitability index, HSI)模型最早是由美國地理調查局國家濕地研究中心魚類與野生生物署于20世紀80年代初提出[1],作為一種評價野生生物生境適宜程度的指數(shù),被廣泛應用于陸生野生生物的生境評價中。近年來,HSI模型被引入到生態(tài)需水、水產(chǎn)養(yǎng)殖、河流多樣性保護和生態(tài)修復等領域,用于預測和模擬水生生物的分布特征,評價人為活動造成的棲息環(huán)境變化對水生生物的影響,受到了生物學家和生態(tài)學家的高度重視[2]。隨著HSI模型的不斷發(fā)展,人們在傳統(tǒng)方法基礎上融入了遙感系統(tǒng)(RS)、地理信息系統(tǒng)(GIS)和全球定位系統(tǒng)(GPS)技術,尤其是依托GIS強大的空間數(shù)據(jù)收集、存儲、分析和圖形化顯示能力,生境適宜性分析的研究范圍不斷擴大,精度不斷提高,分析也更加全面[3-4]。Vincezi[3]等利用溶解氧(DO)、鹽度、水深和葉綠素a等生境因子對地中海馬尼拉蛤(Tapesphilippinarum)生境進行研究,為管理者提供了不同地點的馬尼拉蛤潛在經(jīng)濟產(chǎn)量。王家樵[5]等應用基于GIS軟件的HSI模型對印度洋大眼金槍魚的生境分布進行研究,從而探討了其適宜棲息的海域范圍。生態(tài)位理論指出物種僅能在其特定生境條件范圍內才能生存繁衍[6],而環(huán)境質量并不是一成不變的,隨著時間和地理因素的變遷也會影響物種的生存[7]。生境適宜性(HSI)評價的核心內容就是確定物種所處環(huán)境的變量對物種地理空間分布的影響。
小黑山島臨近海域水質狀況良好,各種理化因子季節(jié)變化明顯,該海域鹽度年均值為30.62,溶解氧年均值為9.10mg/L,四季海水溫度變化幅度較大。潮流運動形式以往復流為主,促進該海域海洋生物的攝食,適宜海洋生物的生長[8]。歷史資料顯示,該海域生物資源量豐富,自然生長的貝類,例如刺參、扇貝、鮑魚、海膽等海珍品享譽海內外[9]。然而,小黑山島海域近年來由于水域環(huán)境污染,該海域生態(tài)環(huán)境受損程度加劇,刺參、魁蚶和紫貽貝等種群資源量日趨減少。本研究通過構建基于GIS的HSI模型,對刺參、魁蚶和紫貽貝的生境適宜性進行分析,確定修復物種適宜增殖修復的區(qū)域,以期為該海域生境修復和資源恢復決策制定提供參考資料。
1 材料與方法
1.1 研究區(qū)域
小黑山島(圖1)位于山東省煙臺長島縣(37°57′55″—37°58′44″N,120°38′21″—120°39′10″E)地處渤海海峽,島陸面積為1.29km2。常年平均水溫為12.3℃[10],最高水溫出現(xiàn)在8月,達23.8℃,最低水溫出現(xiàn)在1月,為1.4℃。葉綠素含量較其他海域偏高,潮間帶底質主要有巖礁、礫石和粗砂底質3種類型,海底地形自西向東、自南向北逐漸傾斜。
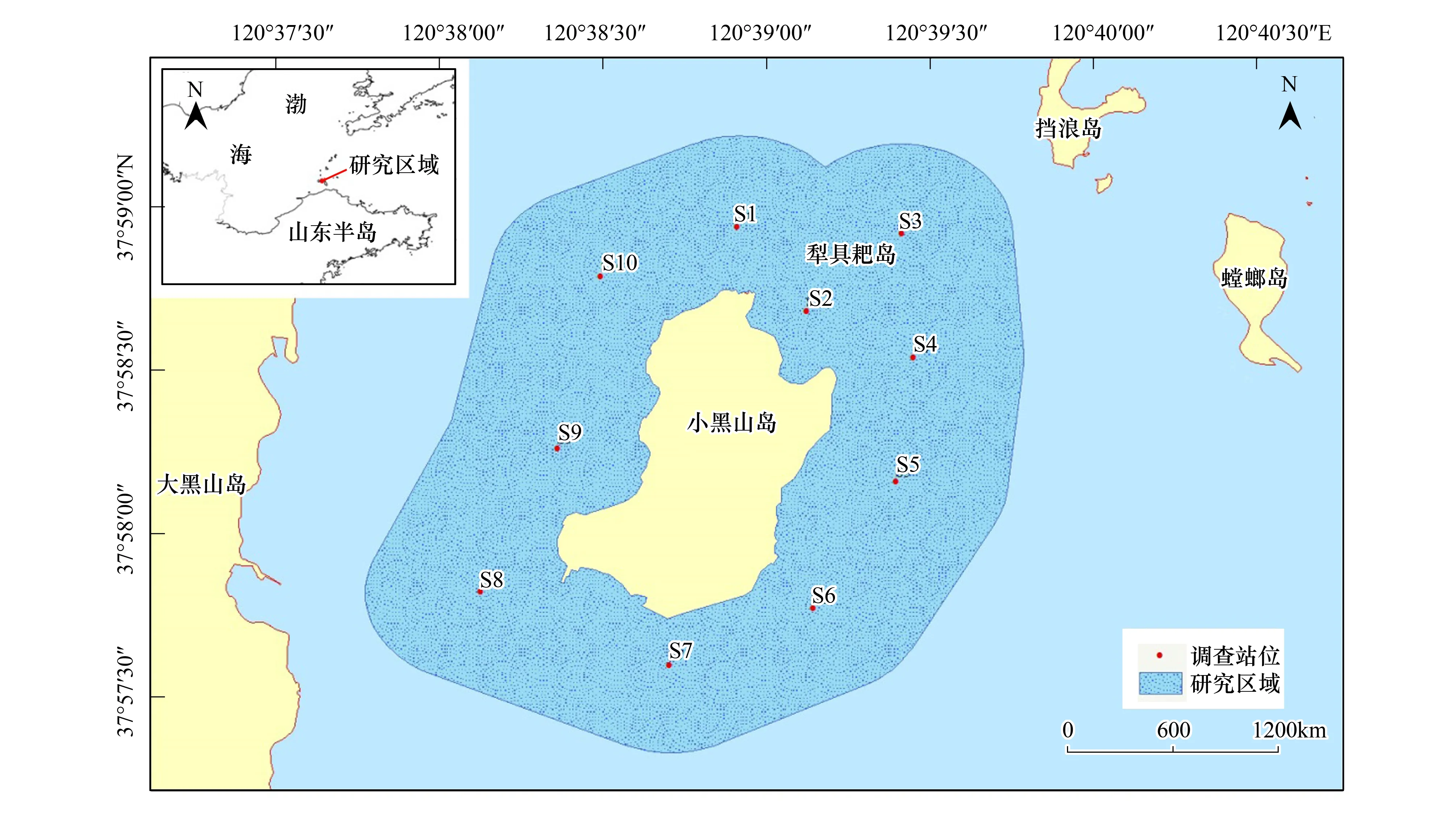
圖1 小黑山島及監(jiān)測站位Fig.1 The Xiaoheishan Island and monitoring station
本研究在環(huán)小黑山島周邊海域布設10個調查站位,并以每個站位點為基準向周邊延伸500m,劃定分析研究范圍,整個研究區(qū)域面積約為6.43km2。
1.2 數(shù)據(jù)來源
本研究所用數(shù)據(jù)來源于2013年的6月、8月、11月及2014年3月進行4個季度的航次調查。按照海洋調查規(guī)范獲得該海域水文要素(溫度、鹽度、深度、透明度等)、化學要素(pH、DO等)、沉積物(粒級含量、粒組系數(shù))等生境參數(shù)。應用反距離插值法將調查數(shù)據(jù)轉化為柵格數(shù)據(jù),并進行空間分析。
1.3 研究方法
1.3.1 生境評價因子篩選
為確保生境適宜性分析評估結果的科學有效性,查閱相關文獻[11-13]中對刺參、魁蚶和紫貽貝生活史及其生態(tài)習性的研究報道,綜合資料收集并結合咨詢專家意見的方式從影響刺參、魁蚶和紫貽貝3個物種的生存、繁殖等諸多因素中分別篩選出7個相對重要的環(huán)境因子作為HSI模型的評價因子(表1)。

表1 3個物種評價因子篩選
1.3.2 構建評價因子適宜性函數(shù)
依據(jù)生物在不同生境條件下生存繁育狀態(tài),將生境劃分為4個等級[14-15](即適宜生境、較適宜生境、基本適宜生境和不適宜生境,在HSI模型中用分段函數(shù)進行表述,并在GIS中應用重分類進行賦值。其中,適宜生境指在該條件下攝食旺盛、活動力強,生物生理狀態(tài)達到最佳,各生境要素的適宜性指數(shù)(Suitability Index,SI)賦值為4;較適宜生境即能夠滿足生物生長發(fā)育的攝食所需,維持正常生長存活,生理活動相對正常,SI賦值為3;基本適宜生境為在該條件下生物能夠耐受環(huán)境壓力,生物能夠生存,SI賦值2;不適宜生境不能提供生物生存所必需的條件,生物無法存活,SI賦值為0。對已篩選出的生境評價因子逐個構建生境適宜性函數(shù),匯總形成生境適宜性等級劃分表(表2—表4)。
刺參是典型的海洋沉積食性生物,田傳遠[16]等研究表明刺參喜棲息在含泥量不超過10%的砂泥底質,于東祥[17]等也通過對刺參自然分布調查結果顯示,底質含泥量在10%以下,有刺參分布,而含泥量高于30%的底質無刺參分布。王學廣[18]等指出刺參為狹溫性動物,最適生長水溫僅為15—18℃,當水溫<5℃,停止攝食;陳勇[19]等則通過設置6個溫度梯度研究了水溫對刺參生長的影響,發(fā)現(xiàn)當水溫>20℃時,刺參停止攝食。水深的選擇是對刺參進行增殖的重要因素,通常刺參大多生活在3—15m水域,成體有向深水區(qū)域移動的現(xiàn)象[19-20]。當鹽度為27—31.5時刺參生長速度最快,當鹽度>34或<25時,活動減弱[18,21-22]。張群樂[22]等指出刺參對海水pH適應范圍較廣,當pH值<6或>9,刺參會停止攝食,活動減弱。于東祥[24]研究指出養(yǎng)殖水體中溶解氧需≥5.0mg/L方可維持刺參的正常生長,當溶解氧≤3.6mg/L,刺參幼體出現(xiàn)死亡,當溶解氧<1.0mg/L,大批刺參出現(xiàn)死亡現(xiàn)象。此外,也有部分研究[17]指出有機碳含量對刺參的生長也有著一定的影響。
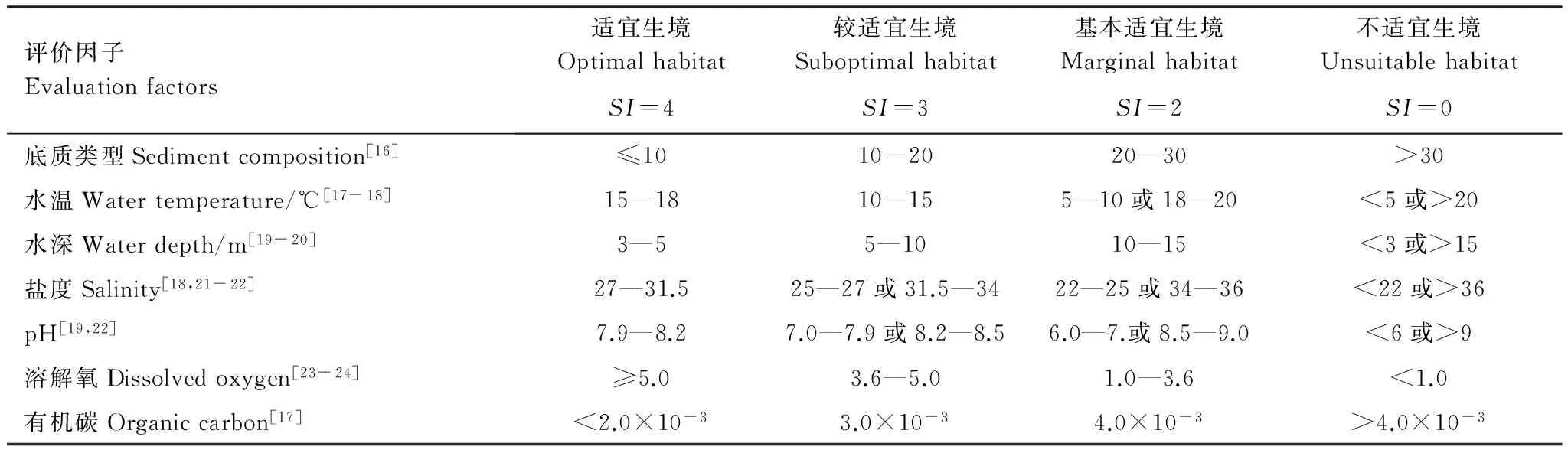
表2 刺參生境適宜性等級劃分表
粒徑>63mm含量:%
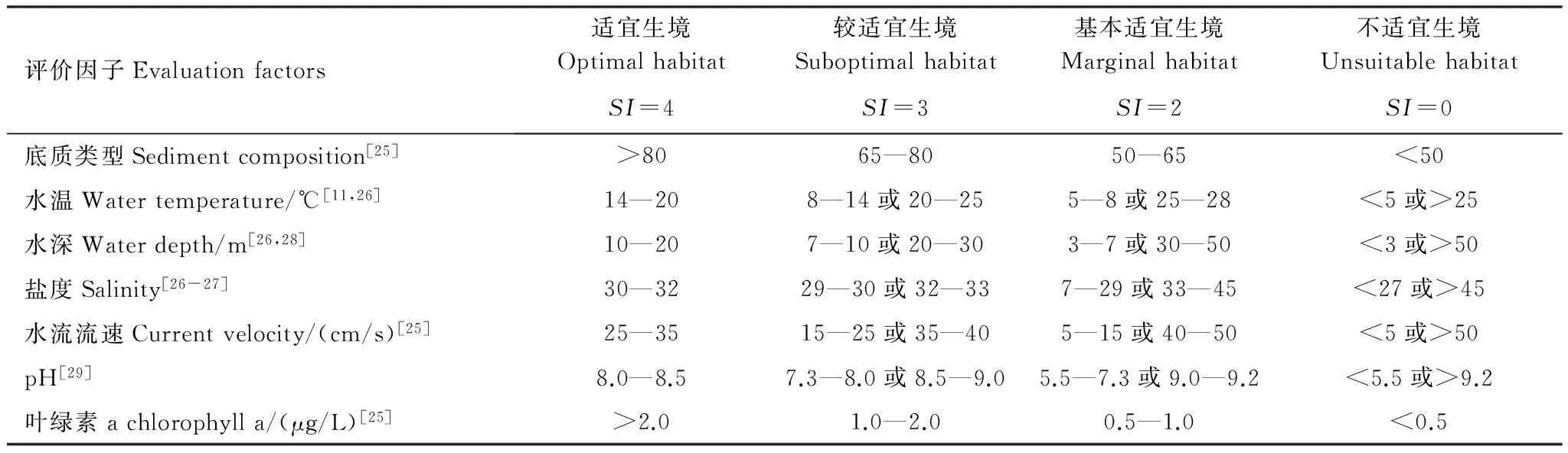
表3 魁蚶生境適宜性等級劃分表
粒徑>63mm含量:%
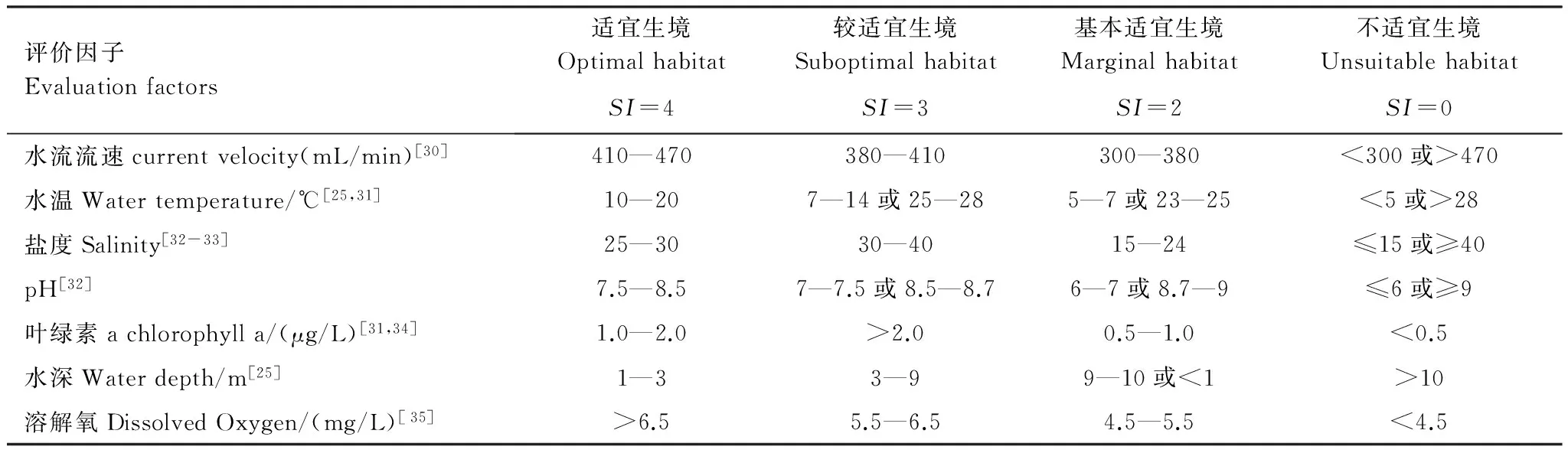
表4 紫貽貝生境適宜性等級劃分表
水流流速[30]參照櫛孔扇貝;.水深[25]參照菲律賓蛤蜊;鹽度[32-33]、pH[32]參照厚殼貽貝
魁蚶營埋棲生活,底質類型對其生長非常重要,在細砂含量80%以上軟泥底質環(huán)境中分布密度較大[25]。張玉璽[11]等、謝忠明[26]等均指出魁蚶為冷水性貝類,適應的水溫范圍為5—25℃,最適宜水溫為14—20℃,而當水溫<5℃或>25℃時,魁蚶會停止生長,出現(xiàn)大量死亡。魁蚶主要棲息于深度為3—50m的海區(qū)[26,28],尤喜好10—20m水深區(qū)域,在>50m海區(qū)幾乎沒有魁蚶分布。魁蚶能耐受的鹽度范圍為27—45,最適宜鹽度為30—32[26-27]。魁蚶適宜生活在風浪不大,相對平靜穩(wěn)定水域,水流流速25—35cm/s為宜,>50cm/s則會影響魁蚶的正常生長[25]。張煥[29]等指出魁蚶稚貝最適宜pH值為8.0—8.5,當pH值>9.2或pH值<5.5時,魁蚶稚貝活動能力減弱,附著能力降低。水體中餌料生物濃度對魁蚶生長也非常重要,相關研究指出葉綠素a濃度>2.0μg/L時,適宜魁蚶的生長,當葉綠素a濃度<0.5μg/L時,則會抑制魁蚶的生長[25]。
目前,已有諸多文獻對影響貝類生長的生境因子進行了報道,但對紫貽貝這一物種的研究相對較少。本文中紫貽貝生境適宜性等級劃分表(表4)中的部分內容,參照與其生態(tài)習性相近的貝類相關資料確定。貽貝喜水流較大且通暢的海況,匡世煥等[30]通過對櫛孔扇貝研究表明,其攝食率隨著流速的加快而提高,當水流流速<300ml/min時,攝食率降低,當流速為470ml/min時,攝食率達到最大值。紫貽貝屬寒溫帶物種,對低溫適應能力較強,王如才[25],包永波[31]等指出紫貽貝適宜生長水溫5—23℃,最適水溫10—20℃,而當水溫<5℃或>28℃時,貽貝停止攝食并出現(xiàn)死亡。紫貽貝適宜生長鹽度一般是25—30,當鹽度過高或過低都會造成攝食活動下降,同時會迫使貝類體內滲透壓發(fā)生改變[32-33]。潘魯青[32]等指出貝類適宜在pH為7—9范圍生長,當pH≤7或≥8.7時,攝食明顯降低。葉綠素是浮游植物生物量的一個重要體現(xiàn),結合包永波[31]和陳清滿[34]對紫貽貝相關研究表明紫貽貝以濾食海水中顆粒物質為食,當餌料濃度為1.0—2.0μg/L,適宜紫貽貝生長,當餌料濃度<0.5μg/L, 抑制貝類的生長。王如才[25]等指出,由于海水表層浮游植物等餌料資源比較豐富,貽貝多聚集分布自潮間帶至3m水深區(qū)域。Chen[35]等研究發(fā)現(xiàn)溶解氧<4.5mg/L時扇貝存活率降低。
1.3.3 權重確定
在專家對評價因子賦值的基礎上,應用層次分析法(Analytic Hierarchy Process, AHP)[36-37]建立層次結構模型,根據(jù)各生態(tài)因子之間關系,建立遞階層次結構,隨后對同一層次的各生境因子重要性大小進行兩兩比較(采用1—9比例標度),分別構建刺參、魁蚶和紫貽貝的兩兩比較判斷矩陣,并通過一致性檢驗(即CR<0.1),檢驗結果(CR值)依次為0.0384; 0.0104; 0.0249。最終確定影響每個生境因子的權重值(表5)。
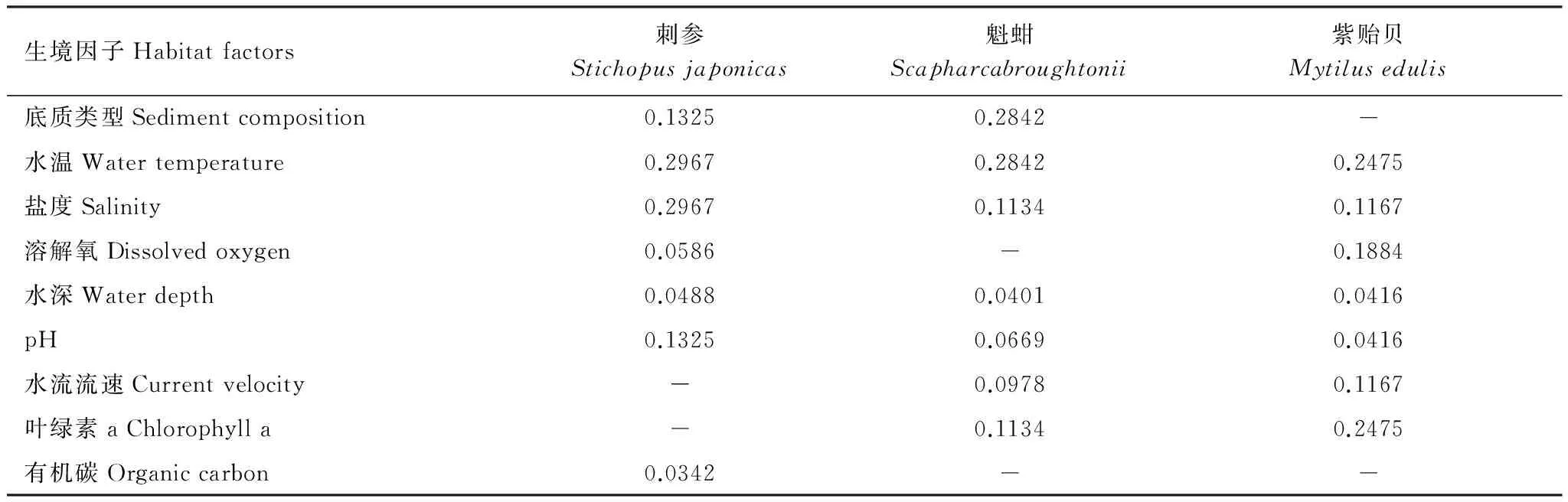
表5 刺參、魁蚶和紫貽貝評價因子權重值
1.3.4 適宜性函數(shù)分析
評價因子的適宜性等級劃分及權重值確定后,將其應用到生境適宜性模型中計算HSI值,并對整個研究區(qū)域的生境適宜性做出綜合分析。基于乘法原理的連乘法(CPM)和幾何平均法(GMM)曾是生境評價(柵格計算環(huán)節(jié))最常用的方法,但Layher[38]和龔彩霞[39]等人均指出這種算法不能很好地模擬生物體與各因子之間的綜合復雜的關系。近年來,越來越多的加權評價模型應用到適宜性分析中[40-41]。本研究中的HSI值計算公式如下:
其中,SIi代表特定因子的適宜性值;Wi代表相應特定的環(huán)境因子(i)的權重值大小;i=1…7代表的是在以上公式中所輸入的7個影響因子。
1.3.5 繪制HSI分布圖
為使分析結果更清晰直觀,依據(jù)HSI計算得分,將生境適宜性劃分為4個等級,并用相對應顏色標明,最終輸出生境適宜性分析圖。其中0—1為不適宜生境(紅色);1—2為基本適宜生境(橙色);2—3為較適宜生境(藍色);3—4為適宜生境(綠色)。
2 結果
2.1 刺參
刺參的生境適宜性在季節(jié)方面存在顯著不同,而調查站位間并未出現(xiàn)差異。該研究區(qū)域在春秋兩季全部屬于適宜生境,非常適宜刺參的生長;在夏季,研究區(qū)均為較適宜生境;冬季則屬于基本適宜生境,刺參僅能勉強存活(圖2—圖5)。
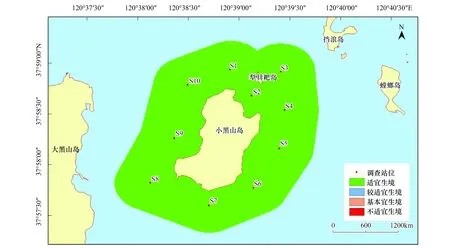
圖2 小黑山島區(qū)域刺參生境適宜性(春季)Fig.2 Map of habitat suitable area of Stichopus japonicas (spring)

圖3 小黑山島區(qū)域刺參生境適宜性(夏季)Fig.3 Map of habitat suitable area of Stichopus japonicas (summer)
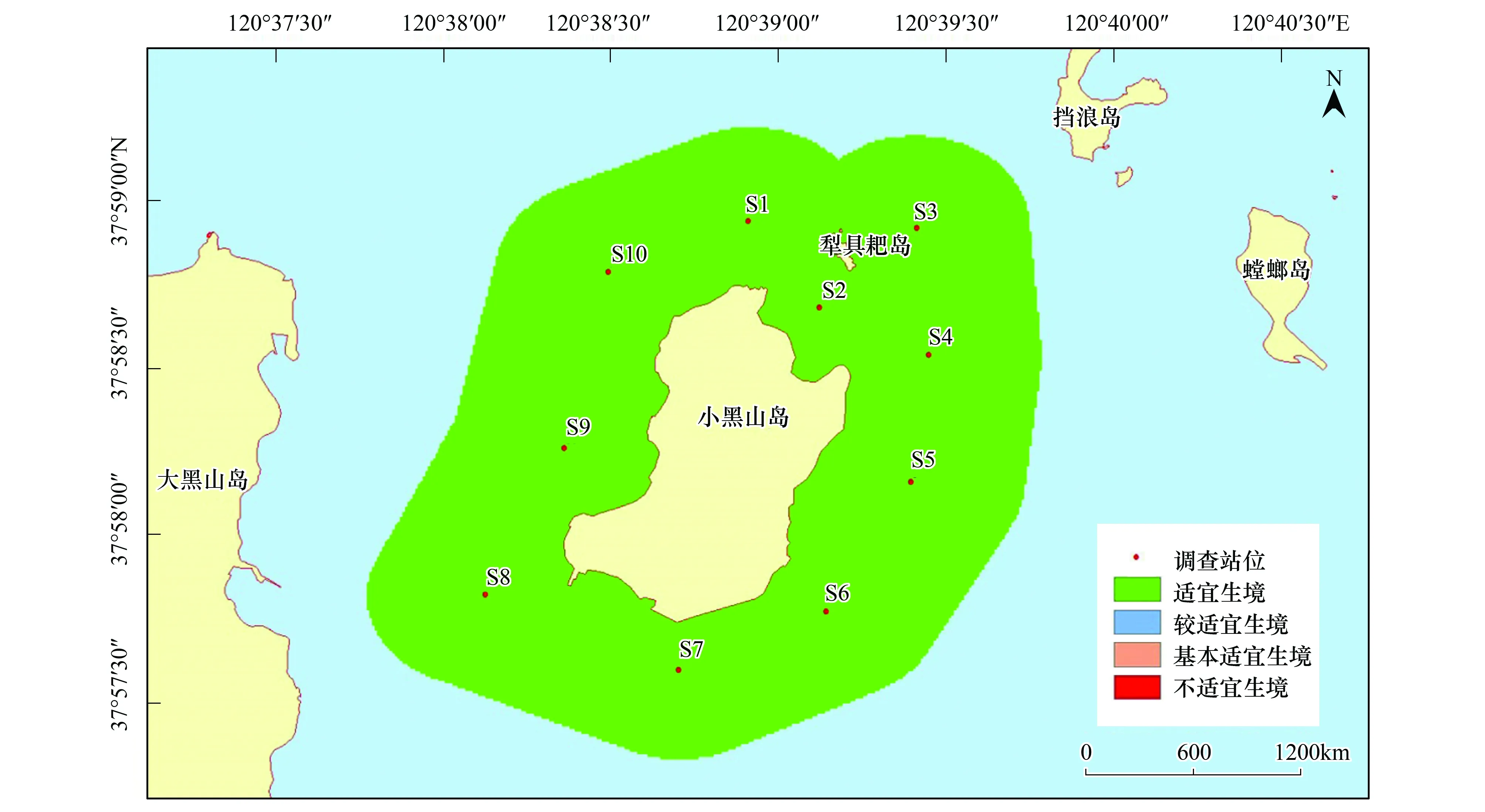
圖4 小黑山島區(qū)域刺參生境適宜性(秋季)Fig.4 Map of habitat suitable area of Stichopus japonicas (autumn)
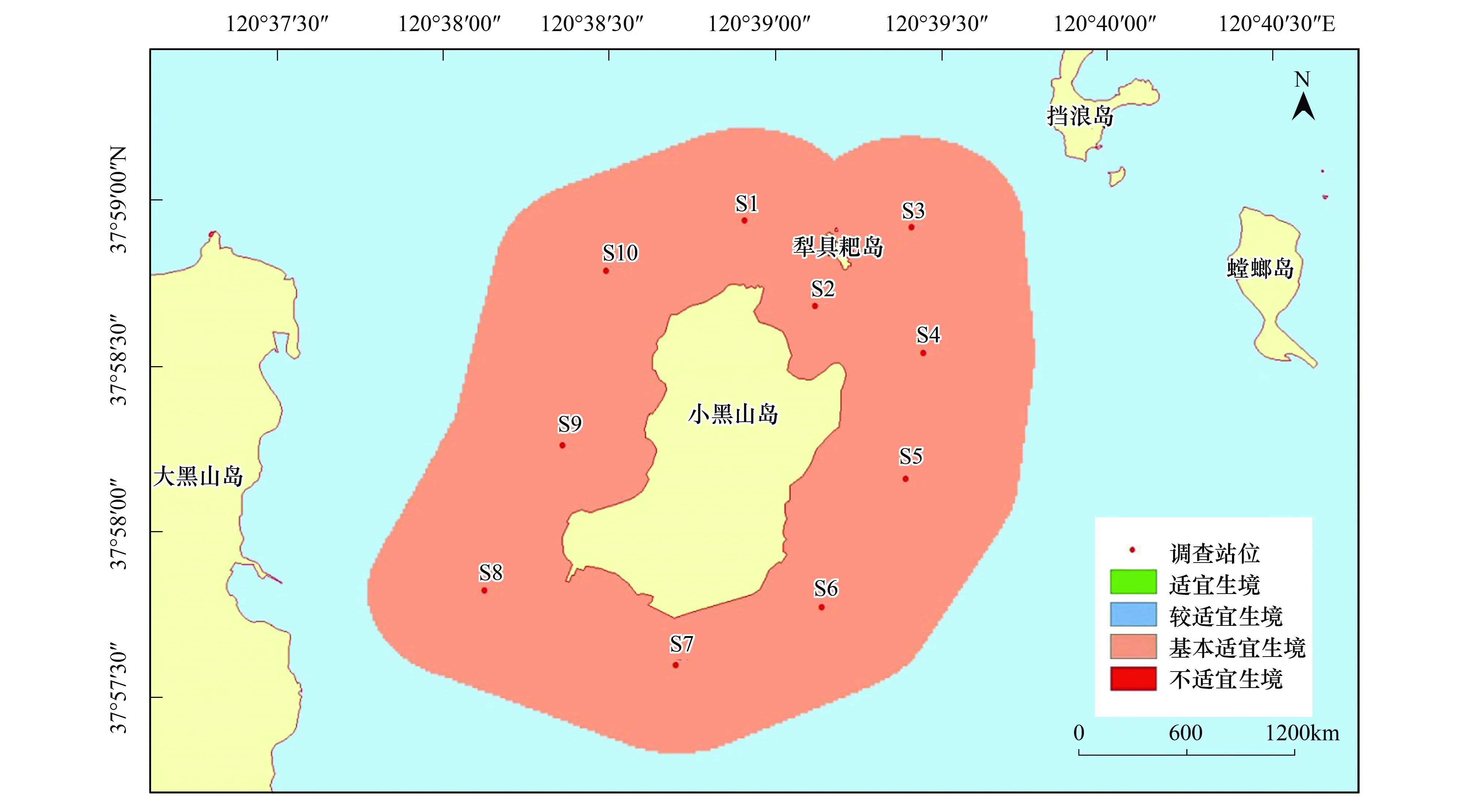
圖5 小黑山島區(qū)域刺參生境適宜性(冬季)Fig.5 Map of habitat suitable area of Stichopus japonicas (winter)
2.2 魁蚶
在本研究區(qū)域內,魁蚶四季均以較適宜生境為主,除S7站位鄰近海域在冬季出現(xiàn)基本適宜生境外,其他季節(jié)均無基本適宜生境和不適宜生境出現(xiàn)(圖6—圖9)。
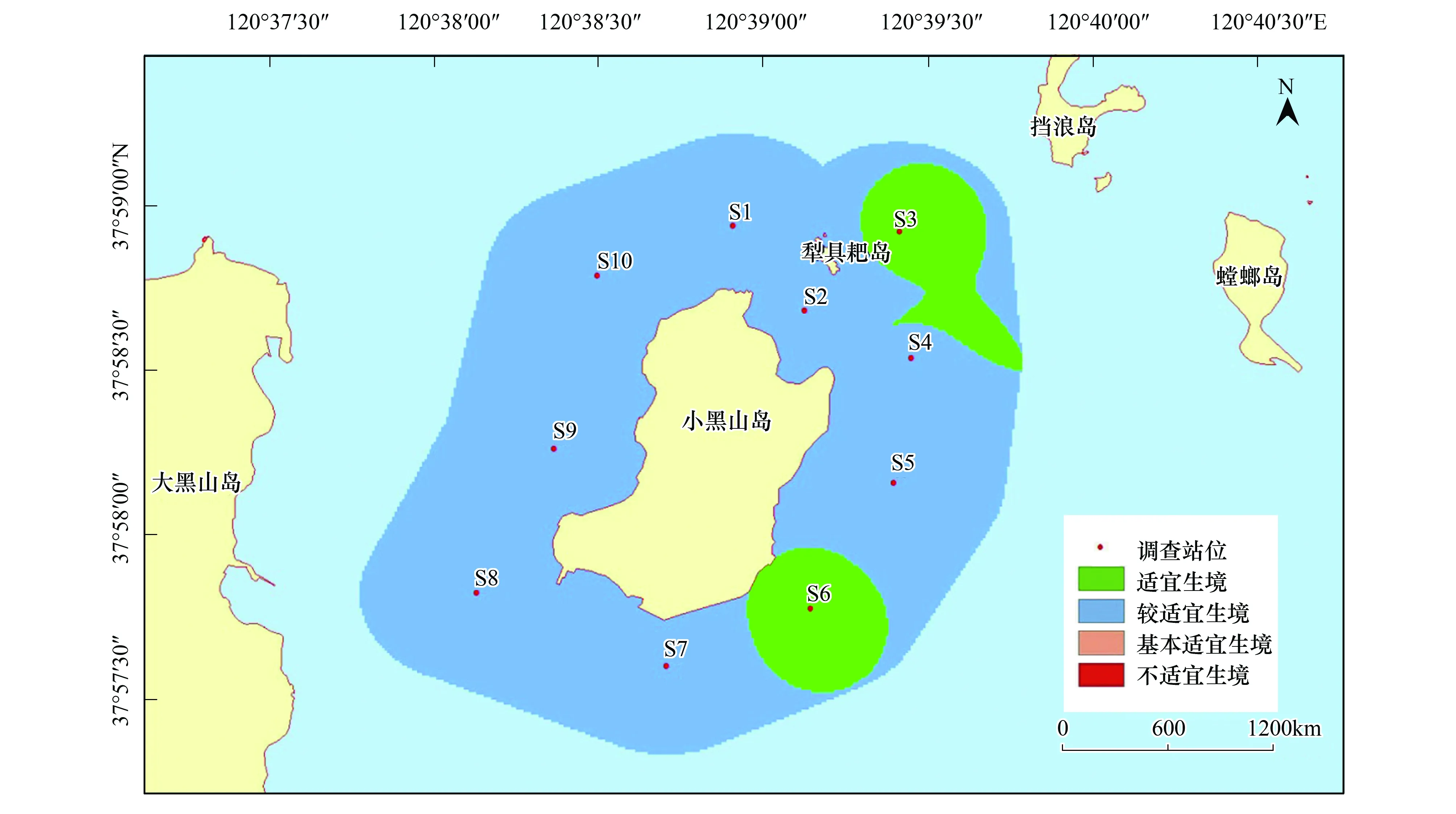
圖6 小黑山島區(qū)域魁蚶生境適宜性(春季)Fig.6 Map of habitat suitable area of Scapharca broughtonii (spring)
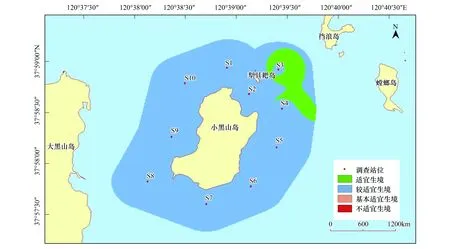
圖7 小黑山島區(qū)域魁蚶生境適宜性(夏季)Fig.7 Map of habitat suitable area of Scapharca broughtonii (summer)
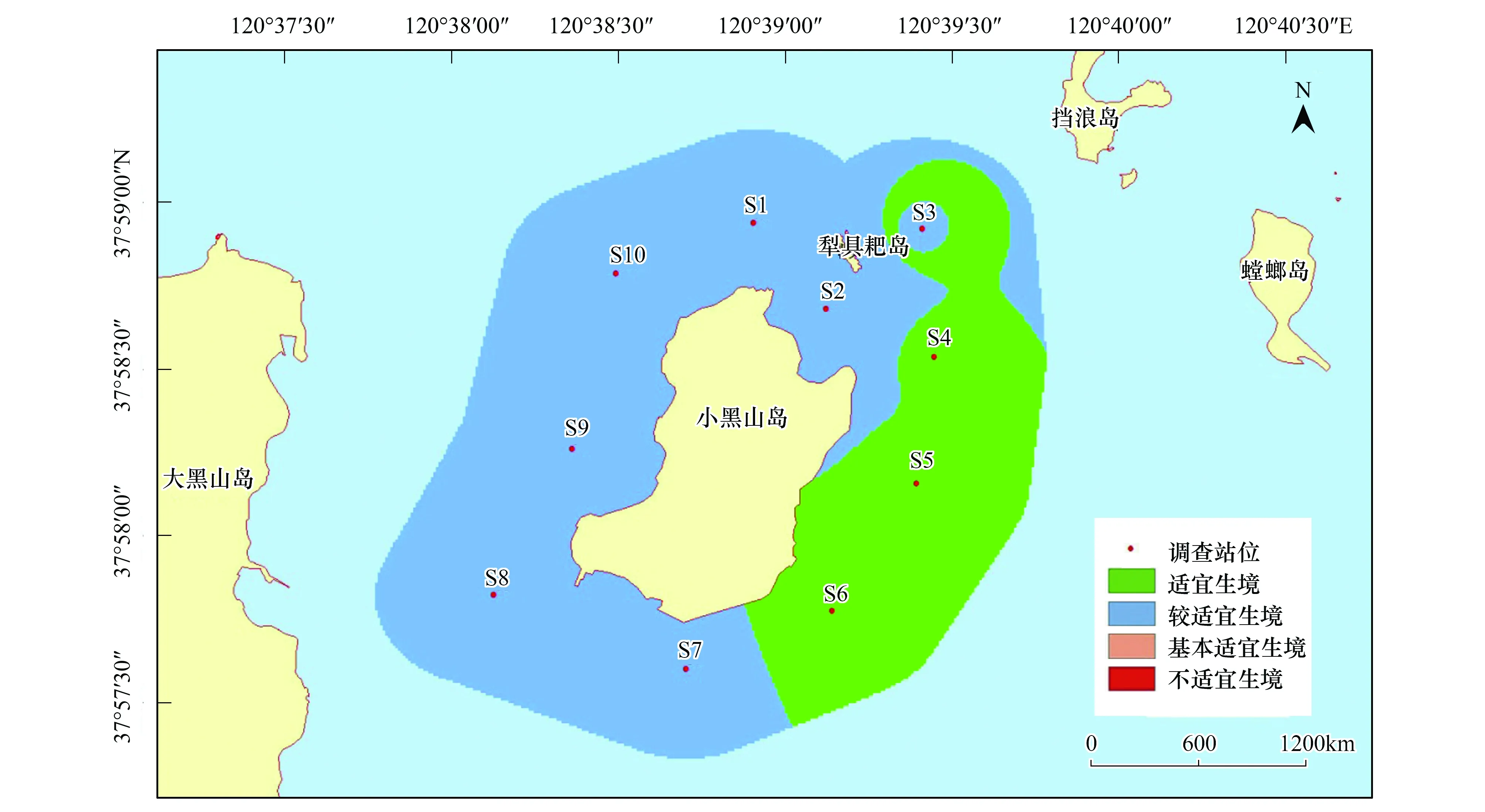
圖8 小黑山島區(qū)域魁蚶生境適宜性(秋季)Fig.8 Map of habitat suitable area of Scapharca broughtonii (autumn)
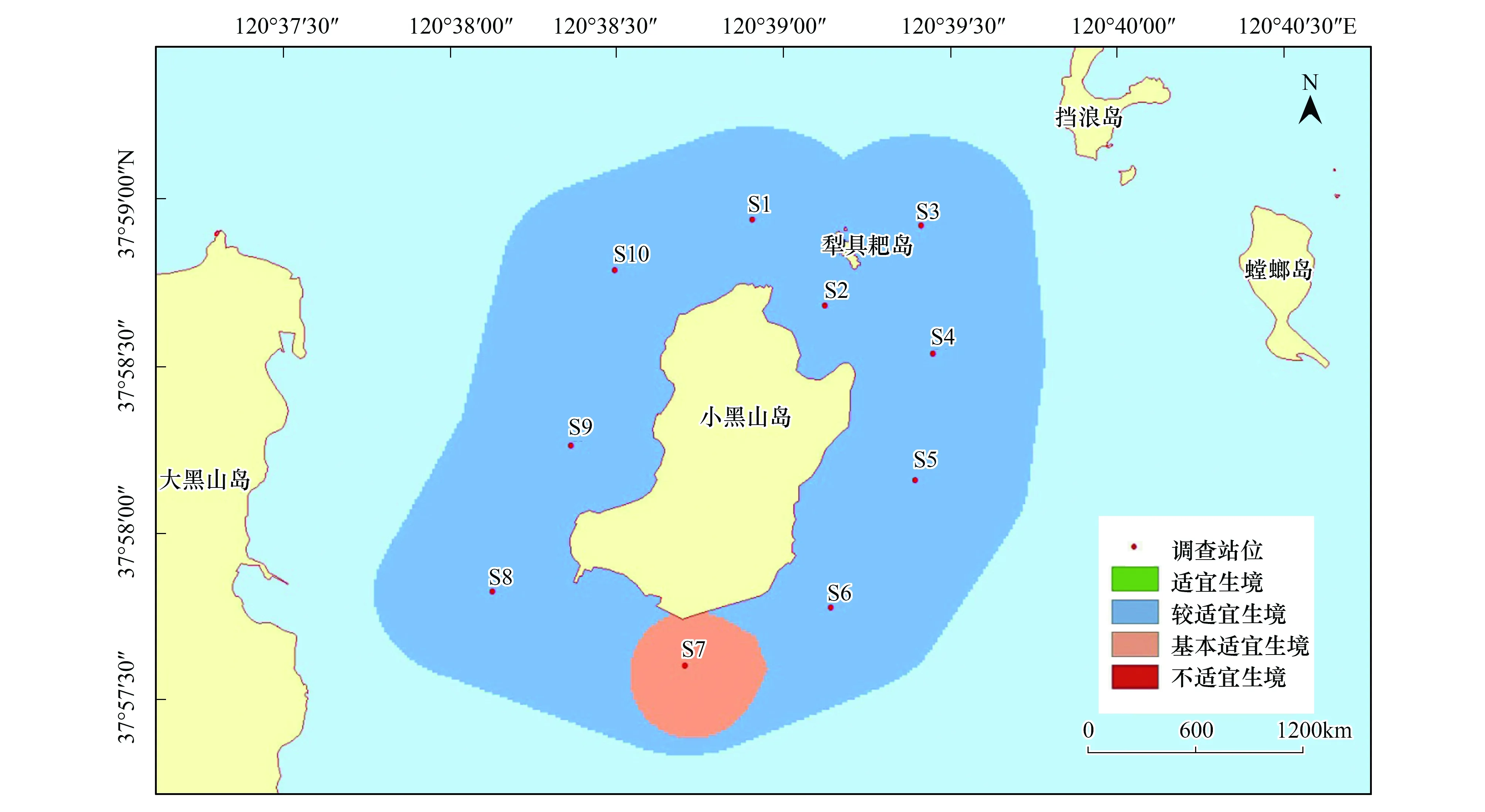
圖9 小黑山島區(qū)域魁蚶生境適宜性(冬季)Fig.9 Map of habitat suitable area of Scapharca broughtonii (winter)
大部分研究區(qū)域在春季屬于較適宜生境(約5.61km2),僅有S3、S6站位周邊區(qū)域屬適宜生境(約0.82 km2);夏季適宜生境出現(xiàn)在小黑山島東北部海域(約為0.49 km2),其他周邊海域均屬于較適宜生境(約5.94 km2);秋季適宜生境面積比其他3個季節(jié)稍大(約2.09km2),較適宜生境出現(xiàn)在S4、S5和S6站位及S3站位周邊區(qū)域;冬季除S7站位及其周邊區(qū)域屬基本適宜生境(約0.38km2)外,其他區(qū)域則屬較適宜生境范圍(約6.05 km2)。魁蚶的4個季節(jié)生境適宜性分析統(tǒng)計結果見表6。
2.3 紫貽貝
與刺參的生境適宜性分析結果類似,紫貽貝的生境適宜性除在季節(jié)上表現(xiàn)出不同外,在調查站位間并不存在差異。紫貽貝在春、夏和秋3個季節(jié),研究區(qū)域均屬于適宜生境;而冬季所研究海域則屬于較適宜生境(圖10—圖13)。
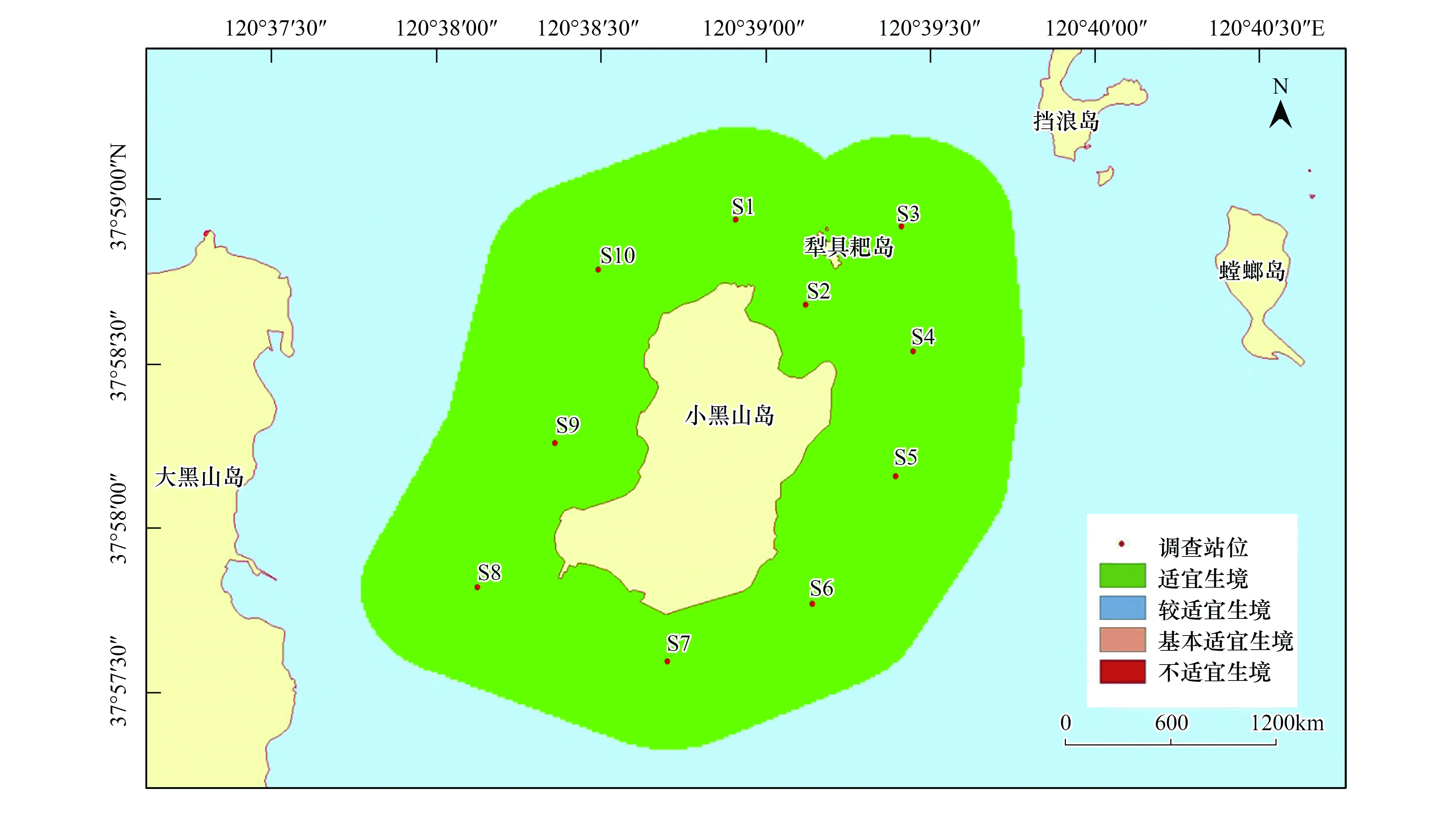
圖10 小黑山島區(qū)域紫貽貝生境適宜性(春季)Fig.10 Map of habitat suitable area of Mytilus edulis (spring)
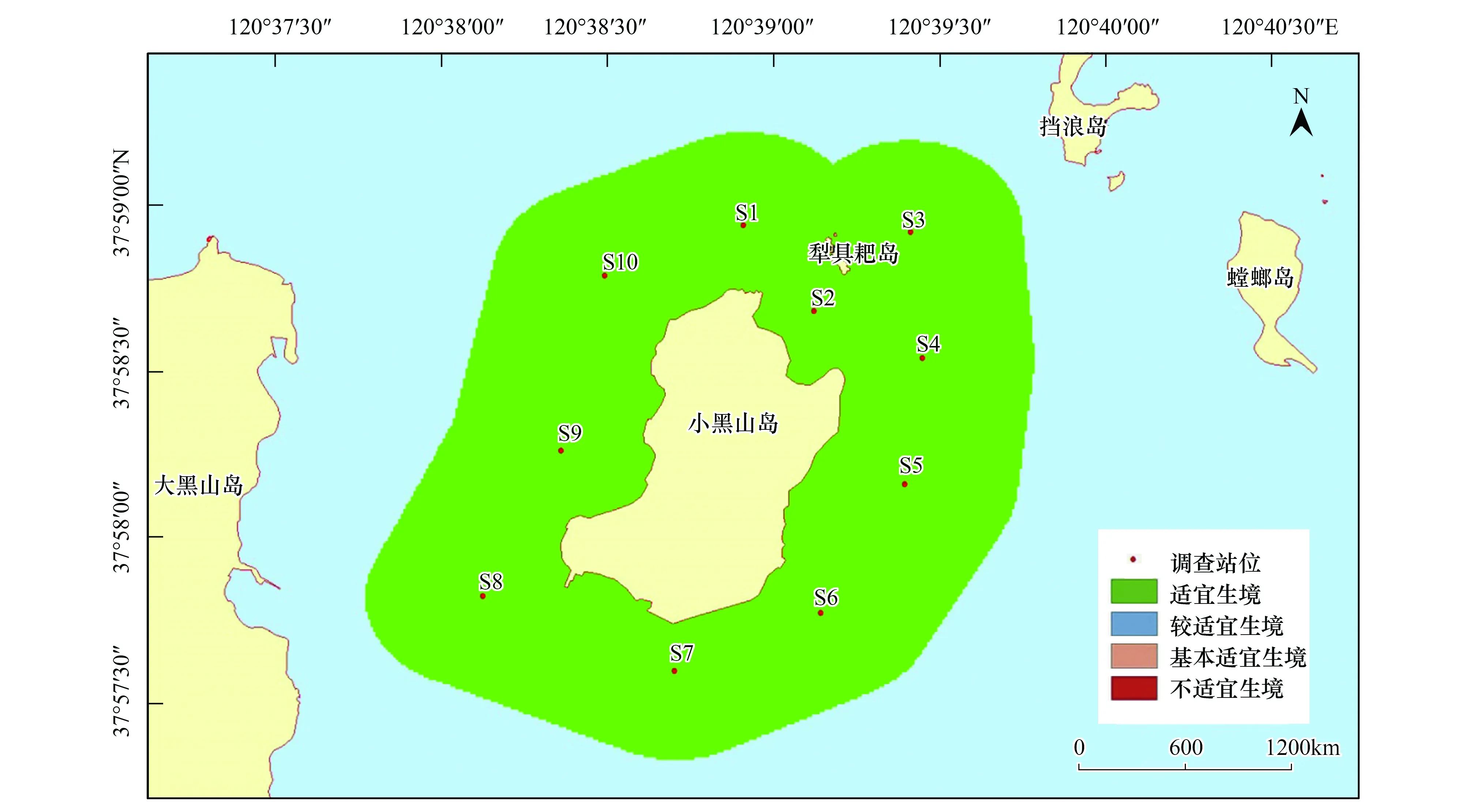
圖11 小黑山島區(qū)域紫貽貝生境適宜性(夏季)Fig.11 Map of habitat suitable area of Mytilus edulis (summer)
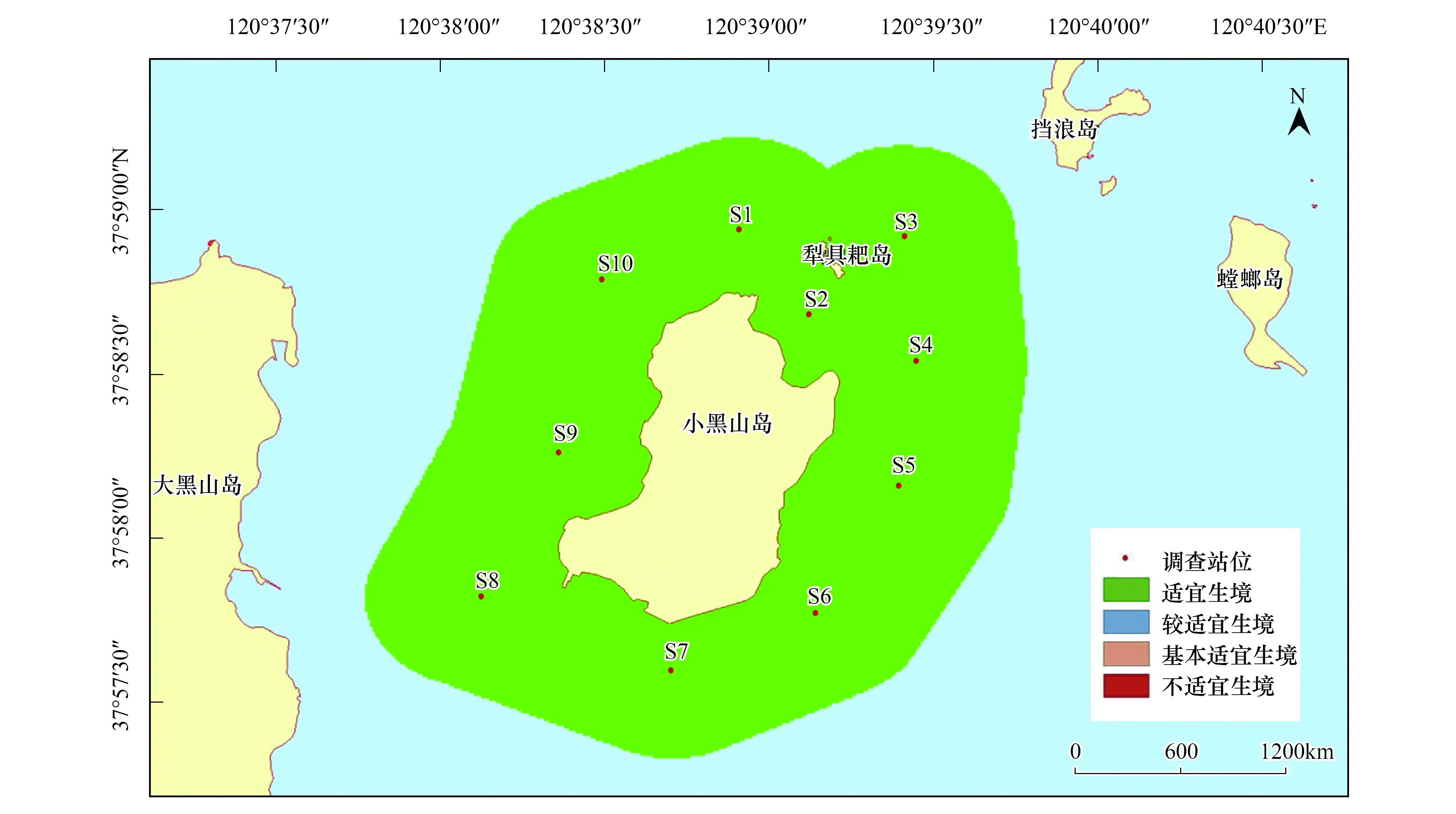
圖12 小黑山島區(qū)域紫貽貝生境適宜性(秋季)Fig.12 Map of habitat suitable area of Mytilus edulis (autumn)
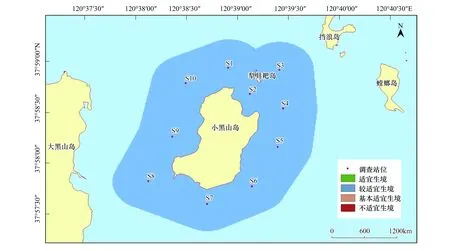
圖13 小黑山島區(qū)域紫貽貝生境適宜性(冬季)Fig.13 Map of habitat suitable area of Mytilus edulis (winter)
3 討論
隨著人類對漁業(yè)資源需求的日益增長,物種生物多樣性受到嚴重威脅,對其進行種群修復已成為全球關注的焦點。據(jù)聯(lián)合國糧食與農(nóng)業(yè)組織(FAO,2007)統(tǒng)計年鑒顯示,全球已有94個國家實施種群增殖修復項目,專家們也明確指出確定增殖物種的修復區(qū)域是決定修復成效的關鍵因素[42-43],但仍有許多海洋生物資源的增殖修復并未達到預期效果[44]。值得注意的是,在實證分析中許多HSI模型往往將年度調查數(shù)據(jù)的均值作為模型的輸入變量,忽略了生境因子季節(jié)差異性等因素的影響。然而,在同一區(qū)域內,部分生境因子(如底質類型)可以認為是恒定的,而其他生境因子(如鹽度、溶解氧和葉綠素a)則呈現(xiàn)出顯著的季節(jié)差異[3,45]。綜上,季節(jié)尺度的生境適宜性分析缺失可能是導致預期修復效果難以確保的重要因素之一。本研究以煙臺長島縣小黑山島臨近海域為研究對象,分別對刺參、魁蚶和紫貽貝的生境適宜性進行了季節(jié)性探討,通過對比分析分別選劃出3個研究物種的適宜修復區(qū)域,以期為確定更加有效的生境修復方案奠定基礎。
如圖2—圖5所示,小黑山島臨近海域常年較適宜刺參的生長,尤其是春季和秋季,而水溫是造成刺參生境適宜性差異的主要原因。刺參為狹溫性海洋底棲生物,夏眠和冬眠是刺參生活史中重要的階段。當外界溫度升高(高于20℃)或降低(低于5℃)到一定的界限時,刺參就會遷移到海水較深海域進行夏眠或冬眠[46]。根據(jù)對研究區(qū)域監(jiān)測顯示,小黑山島海域在春秋季節(jié)水溫變化范圍為12.9℃—14.3℃,鹽度30.26—30.48,pH8.14—8.23,DO高于4mg/L,有機碳0.12%—0.31%,各生境因子均適宜刺參的生長。夏季水溫在23—24.7℃之間(高于20℃,得分為0分),在此期間刺參會遷移到深海區(qū)域進行夏眠,其生長速度延緩。冬季(4.8—5℃)(水溫≤5℃,得分為0分),刺參進行冬眠,停止攝食。程濟生指出,由于黃海海域冷水團的存在,山東半島的巖礁海域為海洋生物資源天然的增殖修復場所[47]。此外,劉夢俠等[48]借助ArcGIS 9.3空間分析模塊,對山東省近海底播增殖區(qū)域和主要增養(yǎng)殖種類進行了評估,指出膠東半島煙臺、威海榮成、乳山等區(qū)域均適宜刺參的增養(yǎng)殖。小黑山島海域地處渤海海峽,位于廟島群島西南部海域,研究表明該海域均適宜刺參生長增殖,與劉夢俠等指出的刺參適宜底播增殖區(qū)域劃定結論相符。
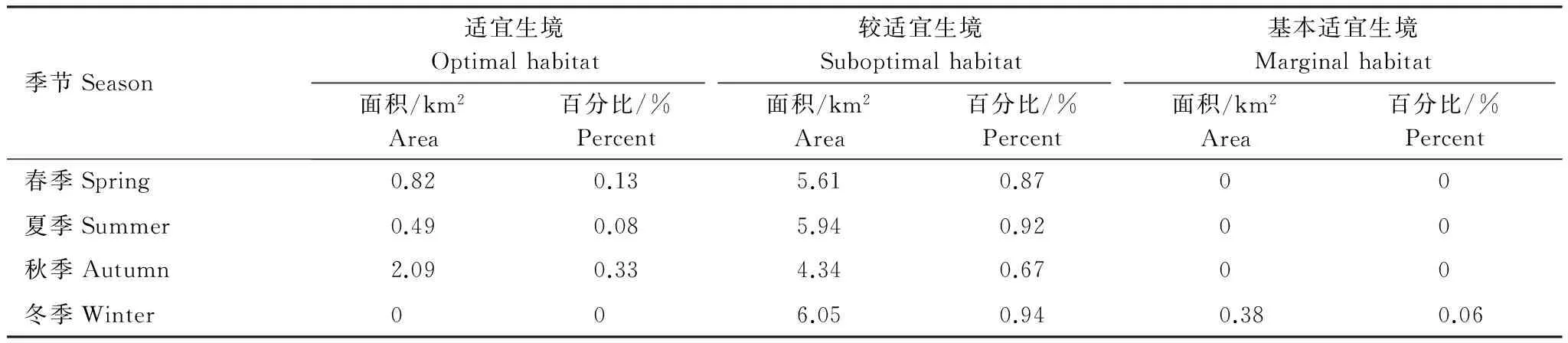
表6 魁蚶4個季節(jié)的生境適宜性分析結果
魁蚶是營底棲生活的貝類,喜泥質或泥沙質海底,其分布區(qū)主要取決于海區(qū)的底質、水溫、鹽度和水深等環(huán)境因子的影響[11]。其中底質(含砂率)類型對其潛沙、水平移動、存活和生長至關重要[49],粒徑越大,抵制流水沖刷的能力就越強。根據(jù)資料顯示,本研究海域底質類型(含砂率)穩(wěn)定,水流流速和鹽度等因子受季節(jié)影響不大,但東部海域含砂率相對較高,更適宜魁蚶的生長。周珊珊[50]通過設置6種不同底質組合實驗,觀察魁蚶稚貝分布情況,證實了魁蚶稚貝對較大粒徑中砂(0.25mm<粒徑<0.50mm)的選擇偏好性。此外,綜合葉綠素a和水溫等因素影響,研究區(qū)域在4個季節(jié)均以較適宜生境為主,春季、夏季和秋季的東部研究海域則為適宜生境。Park[51]等研究發(fā)現(xiàn)冬季或早春水溫的升高可以促進Scapharcasubcrenata(Lischke, 1869)的產(chǎn)卵和孵化,而魁蚶在水溫低于8℃時停止生長,水溫超過25℃則出現(xiàn)大量死亡[52]。小黑山島研究區(qū)域四季水溫變化明顯,受水溫和葉綠素a(>2.0μg/L)等綜合影響,春季S3和S6站位適宜性得分為4分,夏季S3站位周邊海域出現(xiàn)適宜生境;秋季綜合水深(6—9.3m)和葉綠素a影響,S4,S5,S6站位均為適宜生境。冬季S7站位臨近海域受水溫(5℃)和水深(6m)影響為基本適宜生境(0.38km2),其他海域均為較適宜生境。
紫貽貝以濾食顆粒物質為食,適宜生活在風浪小,潮流暢通的淺海區(qū)域,葉綠素a和溶解氧(DO)對紫貽貝的生存繁育有很大影響[34]。據(jù)調查資料顯示,小黑山島區(qū)域的葉綠素a含量平均為0.98mg/m3[8],餌料充足,溶氧含量變化范圍在7.71—12.26mg/L之間,基本能夠滿足紫貽貝生長所需。由圖10—圖13可知,本研究區(qū)域在春、夏和秋季均為紫貽貝生長的適宜生境,冬季則屬較適宜生境。此外,水溫對紫貽貝生存也存在一定的影響,在適宜溫度范圍內,攝食率隨溫度升高而增加,當溫度升到最大值時,攝食率不再升高反而下降[31,53],這也是造成季節(jié)性差異的主要原因。由于配子發(fā)育被視為溫度依賴性和能量需求的過程,因此配子發(fā)育和產(chǎn)卵周期都會受到生長海域中營養(yǎng)條件和溫度變化的影響[54-56]。Rinaldi等[57]基于長達1周年葉綠素a和溫度數(shù)據(jù)跟蹤監(jiān)測資料,通過能量代謝估算,指出Ganzirriz 灣適宜紫貽貝移位增殖,而Faro灣更適宜原位增殖修復。小黑山島冬季各站位水溫均小于5℃(得分為0分),不適宜紫貽貝的生存,而在其他季節(jié)水溫在7—14℃或14—23℃范圍內,得分為3分和4分,相對適宜紫貽貝生長。
對每一物種,從10個調查站位中選擇HSI值季節(jié)變化最大和最小的站位進行敏感性分析,分別對7個環(huán)境因子的實測值增減10%,計算這兩個站位各季節(jié)HSI值的變化,進而得出HSI模型敏感性指數(shù)(Coefficients of Sensitivity,CS)變動范圍。結果顯示,對于刺參來說,鹽度值發(fā)生變化后,其CS值變化幅度最大(0.812—1.571);對于魁蚶來說,底質類型變化引起的CS值變化幅度最大,并出現(xiàn)最高值2.859;對于紫貽貝來說,水溫發(fā)生變化后,其CS值變化幅度最大(0—2.589)。此外,表2—表4是本研究劃定生境適宜等級的依據(jù),在HSI模型中用分段函數(shù)進行表述,并在GIS中應用重分類進行賦值,這就經(jīng)一步降低了不同生境因子實測值變化所帶來的敏感性差異。
Pastres等[58]將GIS空間分析模塊和3維水動力模型相結合,構建了涵蓋14個狀態(tài)變量、52個環(huán)境參數(shù)的復合營養(yǎng)模型,并以此對菲律賓蛤蜊(Tapesphilippinarum)在Venice湖的生境適宜性和經(jīng)濟產(chǎn)量進行了分析估算。Longdill等[59]綜合社會和環(huán)境因素的考量,篩選出14個模型輸入變量,確定了翡翠貽貝(Pernacanaliculus)在Plenty灣的適宜增殖區(qū)域。然而,Kliskey等[60]和J?rgensen[61]均指出HSI模型輸入變量需要兼顧低成本、可操作性。本研究基于小黑山島鄰近海域的可獲得數(shù)據(jù)資料,去除了Pastres等[58]和Longdill等[59]構建模型中一些高費用及難以量化的參數(shù)(如氧化還原電位、功能區(qū)劃分等),簡化形成了包含底質類型、水溫、鹽度、溶解氧、pH、水深及葉綠a(或水流流速,或有機碳)等7個參數(shù)的HSI模型。
綜上,對于刺參和紫貽貝,本研究區(qū)域均適宜其生長繁殖,同一物種,相同季節(jié)在空間上無站位差異,但各季節(jié)的生境適宜性分區(qū)變化明顯;對于魁蚶來說,東北部海域較為適宜增殖,其次為西部海域,四季均以較適宜生境為主,除冬季出現(xiàn)基本適宜生境外,其他季節(jié)均無基本適宜生境和不適宜生境出現(xiàn)。因此,在本研究區(qū)域開展刺參、魁蚶和紫貽貝3個物種的資源修復具有一定的可行性,為后續(xù)的生物多樣性維護和生境修復決策制定提供了基礎資料參考。HSI模型在預測方面具有一定優(yōu)勢,但該模型并不完善,且有一定局限性[62]。其一,本研究中生境適宜性評價雖是分季節(jié)進行的分析,但由于現(xiàn)有資料的不足,本模型所選用的評價指標并未納入物種適宜因子在生活史差異上的考量,制約了評價結果的精確性;其二,該模型并未對疾病、捕食競爭以及其他的自然因素或人為干擾造成的影響進行檢測,一旦這些影響因素變成可利用因素,生境適宜性分析也將更加完善。可以預見的是,隨著生境數(shù)據(jù)越來越易于獲取和空間分析軟件(GIS)的日益完善,HSI模型對物種分析范圍將不斷擴大,分析精度也將進一步提高。
[1] U.S. Fish, Wildlife Service. Standards for the Development of Habitat Suitability Index Models, Ecological Services Manual 103. Washington D C: U.S. Fish and Wildlife Service, 1981: 1-81.
[2] 龔彩霞. 基于棲息地指數(shù)的西北太平洋柔魚漁獲量估算[D]. 上海: 上海海洋大學, 2012.
[3] Vincenzi S, Caramori G, Rossi R, De Leo G A. A GIS-based habitat suitability model for commercial yield estimation ofTapesphilippinarumin a Mediterranean coastal lagoon (Sacca di Goro, Italy). Ecological Modelling, 2006, 193(1/2): 90-104.
[4] 路春燕. 基于GIS與Fuzzy的野生植物生境適宜性評價與區(qū)劃研究-以秦嶺地區(qū)華中五味子為例[D]. 西安: 陜西師范大學, 2012.
[5] 王家樵, 朱國平, 許柳雄. 基于HSI模型的印度洋大眼金槍魚棲息地研究. 海洋環(huán)境科學, 2009, 28(6): 739-742.
[6] Chase J M, Leibold M A. Ecological Niches: Linking Classical and Contemporary Approaches. 2nd ed. Chicago, IL, USA: University of Chicago Press, 2003.
[7] Gilbert B, Lechowicz M J. Neutrality, niches, and dispersal in a temperate forest understory. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101(20): 7651-7656.
[8] 《中國海島志》編纂委員會.《中國海島志》(山東卷第一冊). 北京: 海洋出版社, 2013.
[9] 孔梅, 黃海軍, 高興國, 馬立杰, 劉艷霞. 長島縣海島開發(fā)活動的環(huán)境效應評價. 海洋科學, 2010, 34(10): 96-100.
[10] 李榮升, 趙善倫. 山東海洋資源與環(huán)境. 北京: 海洋出版社, 2002.
[11] 張玉璽, 周學家, 姜海濱, 王世信, 劉愛英, 劉信藝, 賀光明, 常建波, 曹旭清. 山東近海魁蚶生態(tài)習性的調查 // 中國貝類學會. 貝類學論文集(第五、六輯). 青島: 海洋大學出版社, 1995.
[12] 陳清滿, 章守宇, 林軍, 趙靜, 葉靈娜. 枸杞島紫貽貝生長狀況及其與環(huán)境因子關系. 上海海洋大學學報, 2012, 21(5): 809-815.
[13] 劉永宏, 李馥馨, 宋本祥, 孫慧玲, 張榭令, 顧本學. 刺參(ApostichopusjaponicusSelenKa)夏眠習性研究I-夏眠生態(tài)特點的研究. 中國水產(chǎn)科學, 1996, 3(2): 41-48.
[14] Hirzel A H, Le Lay G. Habitat suitability modelling and niche theory. Journal of Applied Ecology, 2008, 45(5): 1372-1381.
[15] Ortigosa G R, De Leo G A, Gatto M. VVF: integrating modelling and GIS in a software tool for habitat suitability assessment. Environmental Modelling & Software, 2000, 15(1): 1-12.
[16] 田傳遠, 李琪, 梁英. 刺參健康養(yǎng)殖技術. 北京: 中國海洋大學出版社, 2008.
[17] 于東祥, 孫慧玲, 陳四清, 張巖. 海參健康養(yǎng)殖技術. 北京: 海洋出版社, 2005.
[18] 王學廣. 環(huán)境因子對刺參行為及生長影響的研究[D]. 濟南: 山東大學, 2013.
[19] 陳勇, 高峰, 劉國山, 邵麗萍, 石國鋒. 溫度、鹽度和光照周期對刺參生長及行為的影響. 水產(chǎn)學報, 2007, 31(5): 687-691.
[20] 張連慶, 吳遠起. 砣磯島沿海刺參的分布和生態(tài)調查. 齊魯漁業(yè), 1990, 7(5): 34-36.
[21] 常忠岳, 衣吉龍, 幕康慶. 影響刺參生長及成活的因素. 河北漁業(yè), 2003, (2): 32-36.
[22] 張群樂, 劉永宏. 海參海膽增養(yǎng)殖技術. 青島: 青島海洋大學, 1998:1-7, 53-57, 65-66.
[23] 楊秀蘭, 王鵬飛, 焦玉龍, 周全利, 李華東, 李永昌, 劉剛, 張華革. 刺參中間培育及生長特性的研究. 齊魯漁業(yè), 2005, 22(10): 1-4.
[24] 于東祥, 燕敬平, 孫慧玲, 陳碧鵑. 刺參養(yǎng)殖水質的變化特點和調控. 齊魯漁業(yè), 2008, 25(3): 1-7.
[25] 王如才, 王昭萍, 張建中. 海水貝類養(yǎng)殖學. 青島: 青島海洋大學出版社, 1993: 297-300.
[26] 謝忠明, 燕敬平, 陳世杰. 海水經(jīng)濟貝類養(yǎng)殖技術. 北京: 中國農(nóng)業(yè)出版社, 2003.
[27] 孫鵬飛, 徐維柱, 王衛(wèi)民, 宋泉清, 高成美. 淺海底播養(yǎng)殖魁蚶試驗. 水產(chǎn)養(yǎng)殖, 2009, 30(11): 14-14.
[28] 劉世祿, 楊愛國. 中國主要海產(chǎn)貝類健康養(yǎng)殖技術. 北京: 海洋出版社, 2005.
[29] 張煥, 宋國斌, 齊曉陸, 田春雨. pH和氨氮對魁蚶稚貝生長與存活的影響. 中國農(nóng)業(yè)信息, 2013(13): 139-140.
[30] 匡世煥, 方建光, 孫慧玲, 李鋒. 桑溝灣櫛孔扇貝不同季節(jié)濾水率和同化率的比較. 海洋與湖沼, 1996, 27(2): 194-199.
[31] 包永波, 尤仲杰. 幾種因子對海洋濾食性貝類攝食率的影響. 水產(chǎn)養(yǎng)殖, 2004, 25(6): 29-33.
[32] 潘魯青, 范德朋, 馬甡, 董雙林. 環(huán)境因子對縊蟶濾水率的影響. 水產(chǎn)學報, 2002, 26(3): 226-230.
[33] Navarro J M. The effects of salinity on the physiological ecology ofChromytiluschorus(Molina, 1782) (Bivalvia: Mytilidae). Journal of Experimental Marine Biology and Ecology, 1988, 122(1): 19-33.
[34] 陳清滿. 枸杞島海域養(yǎng)殖紫貽貝生長與生態(tài)因子關系初步研究[D]. 上海: 上海海洋大學, 2012.
[35] Chen J H, Mai K S, Ma H M, Wang X J, Deng D, Liu X W, Xu w, Zhang W B, Tan B P, Ai Q H. Effects of dissolved oxygen on survival and immune responses of scallop (ChlamysfarreriJones et Preston). Fish & Shellfish Immunology, 2007, 22(3): 272-281.
[36] 郭金玉, 張忠彬, 孫慶云. 層次分析法的研究與應用. 中國安全科學學報, 2008, 18(5): 148-153.
[37] 鄧雪, 李家銘, 曾浩鍵, 陳俊羊, 趙俊峰. 層次分析法權重計算方法分析及其應用研究. 數(shù)學的實踐與認識, 2012, 42(7): 93-100.
[38] Layher W G, Maughan O E. Spotted bass habitat evaluation using an unweighted geometric mean to determine HSI value. Proceedings of the Oklahoma Academy of Science, 1985, 65: 11-17.
[39] 龔彩霞, 陳新軍, 高峰, 官文江, 雷林. 棲息地適宜性指數(shù)在漁業(yè)科學中的應用進展. 上海海洋大學學報, 2011, 20(2): 260-269.
[40] Acevedo P, Cassinello J. Human-induced range expansion of wild ungulates causes niche overlap between previously allopatric species: red deer and Iberian ibex in mountainous regions of southern Spain. Annales Zoologici Fennici, 2009, 46(1): 39-50.
[41] 嚴輝, 段金廒, 孫成忠, 于光, 江曙. 基于TCMGIS的明黨參產(chǎn)地適宜性研究. 南京中醫(yī)藥大學學報, 2012, 28(4): 363-366.
[42] Gomez E D, Mingoa-Licuanan S S. Achievements and lessons learned in restocking giant clams in the Philippines. Fisheries Research, 2006, 80(1): 46-52.
[43] Bell J D, Leber K M, Blankenship H L, Loneragan N R, Masudae R. A new era for restocking, stock enhancement and sea ranching of coastal fisheries resources. Reviews in Fisheries Science, 2008, 16(1/3): 1-9.
[44] 程家驊, 姜亞洲. 海洋生物資源增殖放流回顧與展望. 中國水產(chǎn)科學, 2010, 17(3): 610-617.
[45] Li F Q, Cai Q H, Fu X C, Liu J K. Construction of habitat suitability models (HSMs) for benthic macroinvertebrate and their applications to instream environmental flows: a case study in Xiangxi River of Three Gorges Reservior region, China. Progress in Natural Science, 2009, 19(3): 359-367.
[46] 趙鵬. 刺參Apostichopusjaponicus攝食選擇性的基礎研究[D]. 青島: 中國科學院研究生院(海洋研究所), 2010.
[47] 程濟生. 黃渤海近岸水域生態(tài)環(huán)境與生物群落. 青島: 中國海洋大學出版社, 2004: 363-365.
[48] 劉夢俠, 王其翔, 劉名, 周健, 高翔, 劉洪軍, 官曙光. 山東省近海底層生物增養(yǎng)殖適宜性研究. 北京: 海洋出版社, 2011.
[49] 王如才. 中國水生貝類原色圖鑒. 杭州: 浙江科學技術出版社, 1988: 142-143.
[50] 周珊珊, 張秀梅, 劉旭緒, 張沛東. 魁蚶稚貝的底質選擇性及其潛沙能力評價. 水產(chǎn)學報, 2015, 39(6): 867-875.
[51] Park H J, Lee W C, Choy E J, Choi K S, Kang C K. Reproductive cycle and gross biochemical composition of the ark shellScapharcasubcrenata(Lischke, 1869) reared on subtidal mudflats in a temperate bay of Korea. Aquaculture, 2011, 322-323: 149-157.
[52] 卓亮亮. 魁蚶養(yǎng)殖中有害生物東亞殼菜蛤(Musculistasenhousia)的污染控制及繁殖生物學研究[D]. 青島: 中國海洋大學, 2010.
[53] Bamber R N. The effects of acidic sea water on young carpet-shell clamsVenerupisdecussata(L.) (Mollusca: Veneracea). Journal of Experimental Marine Biology and Ecology, 1987, 108(3): 241-260.
[54] Kang C K, Park M S, Lee P Y, Choi W J, Lee W C. Seasonal variations in condition, reproductive activity, and biochemical composition of the Pacific oyster,Crassostreagigas(Thunberg), in suspended culture in two coastal bays of Korea. Journal of Shellfish Research, 2000, 19(2): 771-778.
[55] Navarro E, Iglesias J I P, Larraaga A. Interannual variation in the reproductive cycle and biochemical composition of the cockleCerastodermaedulefrom Mundaca Estuary (Biscay, North Spain). Marine Biology, 1989, 101(4): 503-511.
[56] Ruiz C, Martinez D, Mosquera G, Abad M, Sánchez J L. Seasonal variations in condition, reproductive activity and biochemical composition of the flat oyster,Ostreaedulis, from San Cibran (Galicia, Spain). Marine Biology, 1992:112(1): 67-74.
[57] Rinaldi A, Montalto V, Manganaro A, Mazzola A, Mirto S, Sanfilippo M, Sarà G. Predictive mechanistic bioenergetics to model habitat suitability of shellfish culture in coastal lakes. Estuarine, Coastal and Shelf Science, 2014, 144: 89-98.
[58] Pastres R, Solidoro C, Cossarini G, Canu D M, Dejak C. Managing the rearing ofTapesphilippinarumin the lagoon of Venice: a decision support system. Ecological Modelling, 2001, 138(1/3): 231-245.
[59] Longdill P C, Healy T R, Black K P. An integrated GIS approach for sustainable aquaculture management area site selection. Ocean & Coastal Management, 2008, 51(8/9): 612-624.
[60] Kliskey A D, Lofroth E C, Thompson W A, Brown S, Schreier H. Simulating and evaluating alternative resource-use strategies using GIS-based habitat suitability indices. Landscape and Urban Planning, 1999, 45(4): 163-175.
[61] J?rgensen S E. Ecosystem Ecology. Fletcher, NC, USA: Academic Press, 2009: 379-379.
[62] Barnes T K, Volety A K, Chartier K, Mazzotti F J, Pearlstine L. A habitat suitability index model for the eastern oyster (Crassostreavirginica), a tool for restoration of the Caloosahatchee Estuary, Florida. Journal of Shellfish Research, 2007, 26(4): 949-959.
Habitat suitability ofStichopusjaponicas,ScapharcabroughtoniiandMytilusedulisin the shallow waters of Xiaoheishan Isalnd
TANG Liuqing1, WANG Qixiang2, LIU Hongjun2, ZHANG Zhipeng1, LIU Chengyue1, ZHOU Jian2,*
1OceanUniversityofChinaCollegeofMarineLife,Qingdao266003,China2MarineBiologyInstituteofShandongProvince,Qingdao266104,China
Xiaoheishan Island is located in Bohai Strait and affiliated with Miaodao Archipelago, which lies in Shandong province. The paper identifies the potential area for sustainable restoration ofStichopusjaponicas,ScapharcabroughtoniiandMytilusedulisin the shallow waters of Xiaoheishan Island using a habitat suitability index model (HSI). Seven screened environmental factors were involved in the HSI model for each species respectively. In summary, the following 9 factors: water temperature, salinity, water depth, pH, sediment composition, dissolved oxygen, organic carbon, current velocity and Chlorophyll a were analyzed in this study. Specific factor piecewise functions have been used to transform environmental factor values into normalized quality indexes (0, 2, 3 and 4). The weight of the each factor was judged with the expert knowledge and the analytic hierarchy process (AHP) method. Thus, the habitat suitable analysis maps were obtained after data interpolate, reclassification and raster calculation with the aid of GIS spatial analysis module. In addition, the analysis of habitat suitability was conducted for four seasons in this paper in order to get more detailed information. First, the results suggest that the entire area was suitable for restoration of S.japonicasand M.edulis, along with no spatial differences but obvious seasonal changes for the 2 species. For S.broughtonii, it is noteworthy that the study area was defined as the suboptimal habitat by the reason of aestivation. Moreover, the northeast region was detected as the optimal site for S.broughtoniipropagation, followed by the western part. Meanwhile, the suboptimal habitat accounted for the majority of study area throughout the year with the marginal habitat only appeared in winter. Water temperature was the main factor causing seasonal variation, while sediments composition was remained as the key reason for the differences in habitat sites. Overall, the results indicated the great feasibility for the long-term survival and successful restoration ofS.japonicas, S.broughtoniiandM.edulis. This study could provide supports and suggestions for subsequent research of biological diversity protection and ecological restoration.
habitat suitability index;Stichopusjaponicas;Scapharcabroughtonii;Mytilusedulis; Xiaoheishan Island
國家自然科學基金資助項目(41206102, 41476091);國家海洋公益專項資助項目(201305009)
2015-07-29;
日期:2016-06-13
10.5846/stxb201507291588
* 通訊作者Corresponding author.E-mail: zhoujian.park@163.com
唐柳青, 王其翔,劉洪軍,張智鵬,劉騁躍,周健.小黑山島海域刺參、魁蚶和紫貽貝生境適宜性分析.生態(tài)學報,2017,37(2):668-682.
Tang L Q, Wang Q X, Liu H J, Zhang Z P, Liu C Y, Zhou J.Habitat suitability ofStichopusjaponicas,ScapharcabroughtoniiandMytilusedulisin the shallow waters of Xiaoheishan Isalnd.Acta Ecologica Sinica,2017,37(2):668-682.
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
小讀者(2021年2期)2021-03-29 05:03:48
科技傳播(2019年22期)2020-01-14 03:06:54
華人時刊(2019年13期)2019-11-17 14:59:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
文苑(2018年22期)2018-11-19 02:54:14
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
中國科技博覽(2016年2期)2016-04-25 20:32:39
小學生導刊(2016年34期)2016-04-11 00:49:44
