RESTORATION OF FLESH FATTY ACID COMPOSITION IN DARKBARBEL CATFISH (PELTEOBAGRUS VACHELLI) USING A FINISHING FISH OIL DIET
2017-02-15 08:23:49SHAOTingQINChuanJieYUANDengYueWENZhengYongandLIHuaTao
水生生物學報 2017年1期
SHAO Ting, QIN Chuan-Jie YUAN Deng-Yue WEN Zheng-Yongand LI Hua-Tao
(1. Key Laboratory of Sichuan Province for Fishes Conservation and Utilization in the Upper Reaches of the Yangtze River, College of Life Science, Neijiang Normal University, Neijiang 641100, China; 2. College of Life Science, Sichuan Normal University, Chengdu 610101, China)
RESTORATION OF FLESH FATTY ACID COMPOSITION IN DARKBARBEL CATFISH (PELTEOBAGRUS VACHELLI) USING A FINISHING FISH OIL DIET
SHAO Ting1,2, QIN Chuan-Jie1, YUAN Deng-Yue1, WEN Zheng-Yong1and LI Hua-Tao1
(1. Key Laboratory of Sichuan Province for Fishes Conservation and Utilization in the Upper Reaches of the Yangtze River, College of Life Science, Neijiang Normal University, Neijiang 641100, China; 2. College of Life Science, Sichuan Normal University, Chengdu 610101, China)
This study aimed to evaluate the effects of 50%—100% soybean oil on growth performance and flesh fatty acid composition of darkbarbel catfish (Pelteobagrus vachelli), so as to assess the effects of refeeding fish oil (FO) on flesh fatty acid composition. Four isonitrogenous, isolipidic diets, i.e., FO, soybean oil (SO), 50% FO+50% SO (S1), and 25% FO+75% SO (S2), were fed to triplicate groups of 40 juvenile P. vachelli [(1.10±0.12) g] for 80d. At the end of the 80d period, all fish were fed with FO for 30d. The results showed that growth rates, hepatosomatic index (HSI), and proximate composition in darkbarbel catfish were not affected by SO. With increasing SO levels, the percentages of oleic acid, arachidonic acid, and monounsaturated fatty acids significantly increased (P<0.05). However, docosahexaenoic acid (DHA), eicosapentaenoic acid (EPA), highly unsaturated fatty acid (HUFA) levels and n-3/n-6 ratios significantly reduced with dietary SO (P<0.05). After 30d on FO, flesh levels of DHA, EPA, and Σ n-3 HUFA significantly increased in groups S2 and SO (P<0.05), but not to the same extent as those in the FO-containing groups except S1. The results revealed that it was possible to substitute almost 100% of FO with SO in the diets of darkbarbel catfish without affecting growth performance. A re-feeding period of 30d with 100% FO significantly increased flesh levels of Σn-3 HUFA, 20:5n-3, and 22:6n-3 in fish which were fed diets containing SO in the first stage.
Pelteobagrus vachelli; Fish oil; Vegetable oil; n-3 high unsaturated fatty acids; Restoration
In 2002, fish oil (FO) consumption in aquaculture was estimated at 81%, which increased to 88% in 2012[1]. Total FO production in the five main FO-producing countries, i.e., Peru, Chile, Iceland, Norway, and Denmark, was 530000 tons in 2009, a decline of 100000 tons compared to 2008[2]. Global FO supplies are becoming more costly and less available; therefore, aquaculture requires sustainable alternatives to FO[1,3]. Vegetable oils (VOs) are considered to be good lipid sources for fish diets. Several VOs have been used as partial or complete replacements of FO[4,5].
VOs are rich in linoleic acid (C18:2 n-6) and oleic acids (C18:1 n-9), but devoid of n-3 highly unsaturated fatty acids (n-3 HUFAs). Linoleic and oleic acids are absent from the natural diets of most fish[6,7]. VOs contribute to imbalances in certain dietary fatty acids and affect the n-6/n-3 dietary ratio, which negatively impacts fish growth. In addition, the complete or partial replacement of FO with VOs is likely to negatively impact intestinal and hepatic cellular structure, intestinal function, immune-associated gene expression, humoral immunity, and disease resistance[8]. However, FO is rich in health-promoting n-3 long chain polyunsaturated fatty acids (n-3 LC-PUFAs), especially eicosapentaenoic acid (EPA), which are beneficial for human health[8,9]. Therefore, in aquacul-
Received date: 2016-03-22; Accepted date: 2016-07-10
Foundation item: Supported by the National Natural Science Foundation of China (No. 31402305); the Educational Commission of Sichuan Province of China (No. 14ZA0249)
Brief introduction of author: Shao Ting, E-mail: 991504974@qq.com; Tel +86-83-2341862
To avoid a reduction in flesh n-3 LC-PUFA levels, VO-fed fish have been switched to FO diets to assess whether seawater fish can recover their n-3 HUFA levels during their final growth phase[6]. These studies, which have been conducted in European sea bass (Dicentrarchus labrax L.), gilthead sea bream (Sparus aurata L.), brown trout (Salmotrutta L), and brook charr (Salvelinus fontinalis)[7,10—12]revealed that introducing FO partly restored docosahexaneoic acid (DHA), arachidonic acid (ArA), and EPA levels in fish[13,14]. In freshwater fish, VOs may induce the activation of ⊿6 and ⊿5 desaturases, which could elongate and desaturate α-linolenic acid (C18: 3n-3) into n-3 HUFA[15]. However, few studies have focused on the effects of VOs and finishing FO diets on the fatty acid composition of freshwater species.
Darkbarbel catfish, Pelteobagrus vachelli, is a valuable commercial species in China. Due to its high market value, the demand for this species has grown considerably in recent years. This study investigatedthe effects of soybean oil on darkbarbel catfish growth performance and flesh fatty acid composition, and the effects of re-feeding FO on flesh fatty acid composition.

Tab. 1 Ingredients (g/100 g) and chemical composition of the experimental diets
1 Materials and methods
1.1 Fish growth and experimental design
Juvenile darkbarbel catfish were obtained from Longfeng fish farm (Meishan, Sichuan, China). Prior to the experiment, fish were fed a commercial catfish diet for 2 weeks (Haida, Chengdu, Sichuan, China). A total of 480 fish [mean weight ± SD, (1.10±0.12) g] were randomly assigned to 12300 L fiberglass tanks (40 fish per tank). Three tanks were assigned to one of four experimental diets. The fish lived in 12h∶12h light∶dark cycles at (25±2)℃ with a mean oxygen concentration of 6.1—7.3 mg/L. Ammonia and nitrate concentrations were < 0.1 mg/L.
1.2 Experimental diets
Four isonitrogenous, isolipidic, and isoenergetic diets were formulated (Tab. 1): (1) 100% fish oil as control (FO); (2) 100% soybean oil (SO); (3) 75% SO+25% FO (S1); and (4) 50% SO+50% FO (S2). The ingredients were thoroughly mixed, extruded (2-mm diameter), and air-dried. The fatty acid compositions of the diets are shown in Tab. 2.
Following the 2-week acclimation period, fish were fed the experimental diets to apparent satiation twice daily (at 08:00 and 16:00) for 80d (growth period, GP); feed consumption was recorded weekly. Subsequently, 33 fish from each tank were fed a 100% FO-based diet for 30d (restoration period, RP).
1.3 Fish performance and sample collection
Fish were fed the experimental diets for 80d, followed by the FO-diet for 30d. At 80 and 110d, fish were fasted for 24h. Three fish per treatment were anaesthetized; tissues were sampled for the determination of live mass, liver mass, hepatosomatic index (HSI), flesh dry mass, specific growth rate (SGR), lipase activity (LPL and HL), and flesh lipid and fatty acid composition. Live mass was determined by blotting biological material onto filter paper before weighing. HSI, SGR, and feed conversion factor (FCR) were calculated by using the following equations: HSI=liver mass/live mass×100; SGR=[Ln(final weight)-Ln(initial weight)]/number of days×100; FCR=dry feed fed/wet weight gain.
1.4 Chemical analyses
Nutritional composition of the flesh and experimental diets were determined by proximate composition analyses (AOAC, 1990). Briefly, moisture content was determined by drying the samples to constant weight at 80℃; protein was determined in anautomated Kjeldahl instrument (SKD-100, Peiou, Shanghai, China) using a protein-nitrogen conversion factor of 6.25; total lipid was determined following chloroform/methanol (2:1 v:v) extraction; and ash was determined by incineration in a muffle furnace at 550℃ for 18h.

Tab. 2 Fatty acid composition of the experimental diets (% total fatty acids; mean±SD, n=3)
1.5 Enzymatic assays
Frozen liver samples were thawed and homogenized in ice-cold 0.9% physiological saline using a Bio-gen Series homogenizer (PRO 200, Oxford, CT, USA). Hepatic homogenates were centrifuged at 15000×g for 15min at 4℃ (Centrifuge 5417R, Eppendorf, Germany). The supernatant, free from lipids, was used to determine the activities of LPL and HL and the soluble protein content. LPL and HL activities were determined using an LPL/HL commercial kit (A067; Nanjing Jiancheng Bioengineering Institute, Nanjing, China) and expressed in units/mg soluble protein. Protein concentration in the homogenates was determined by the Bradford method (1976) using a commercial protein assay kit (Nanjing Jiancheng Bioengineering Institute) with bovine serum albumin as the standard.
1.6 Fatty acid analysis
Lipids were extracted using chloroform and methanol (2∶1)[16]. Fatty acids were converted into methyl esters using an acid-catalyzed methylation method (GB/T22223-2008). The fatty acid profile was determined in a capillary gas chromatograph (GC) coupled to an HP6890 flame ionization detector and an SPTM-2380 column (30 mm×0.25 mm×0.20 mm). Separation was performed using nitrogen as the carrier gas. The column temperature was maintained at 120℃ for 5min, then it was heated to 240℃ at 4℃/min and maintained at this temperature for 20min. The detector temperature and split injector (50∶1) were maintained at 260℃. Fatty acids were identified by comparing their retention times with those of fatty acid standards (Sigma, USA). Peak areas were determined using Varian software.
1.7 Statistical analyses
Results are presented as mean ± standard deviation (mean±SD). Data were analyzed by ANOVA and Duncan's multiple range test. The growth parameters and flesh fatty acid levels in growth and restoration periods were compared using independent t-tests within the same group; P<0.05 was considered statistically significant. Statistical analyses were performed using SPSS software (SPSS 18.0).
2 Results
2.1 Growth performance and proximate composition
With dietary treatments of 80d, there were no significant differences in body weight. A-700% increase in weight was observed, with no animal deaths or differences in feed consumption (Tab. 3). Among the dietary treatments, there were no significant differences in SGR, which ranged from 1.89±0.49 to 2.28±0.39, or in HSI, which ranged from 1.59±0.34 to 1.83±0.38.
After the 30d restoration period on FO, fish grew an additional -31% (-3 g) relative to their weights at the end of the 80d growth period (Tab. 3). Final mean weights at the end of the 30d restoration period ranged from (11.42±2.71) g (FO) to (14.65±4.13) g (S1), with no significant differences among the dietary treatments. However, fish previously fed S1 and SO had higher growth rates than fish previously fed FO or S2. There were no significant differences in SGR, HSI, or FCR at the end of the restoration period. In addition, there were no significant differences in SGR, HSI, or FCR between the growth period and restoration period within the same group (Tab. 3).
Proximate composition analysis of fish fillets revealed no significant differences among the dietary treatments during the growth period or restoration period. During the two periods, fillet lipid levels ranged from (7.83±0.76)% to (8.12±0.68)% and from (7.19±0.97)% to (8.02±0.54)%, respectively (Tab. 4).

Tab. 3 Growth performance of darkbarbel catfish at the end of growth period (GP) and restoration period (RP; % total fatty acids; mean±SD; n=9)
2.2 LPL and HL activities
Tab. 5 shows the effect of SO on hepatic HL and LPL activities. The dietary treatments had no effect on hepatic HL activity, but SO significantly reduced hepatic LPL activity (P<0.05). After re-feeding with FO, LPL activities in groups S2 and SO increased; however, the LPL activity in these groups was significantly lower than those in groups FO and S1 (P<0.05).
2.3 Flesh fatty acid composition during growth and restoration periods
During the 80d growth period, flesh fatty acid composition was affected by dietary treatment (Tab. 6). Total saturated fatty acids (∑SFAs) were 17.85% in SO and 28.51% in FO. SFAs were significantly reduced with dietary SO (P<0.05). However, the diet with 100% SO significantly increased (P<0.05) the levels of monounsaturated fatty acids (MUFAs) such as oleic acid (C18:1n-9). Total n-3 HUFA levels, which ranged from 3.04% in S2 to 4.39% in FO, significantly decreased with increasing SO levels (P<0.05). EPA ranged from 0.45% in SO to 0.66% inFO. DHA significantly decreased with increasing SO levels (P<0.05). However, ARA levels were significantly higher in SO (0.49%) than in FO (0.31%) (P<0.05).

Tab. 4 Proximate composition (%) of darkbarbel catfish at the end of the 80d period on the experimental diets and at the end of the 30d
The flesh fatty acid composition with the 30d restoration FO diet is shown in Tab. 6. For each group, the flesh fatty acid levels in the growth period were compared with those in the restoration period. In S1, flesh 22:6n-3 levels significantly increased (P<0.05). In S2, flesh C18:1n-9 and Σn-6 levels significantly decreased, while flesh Σn-3 HUFA, C20:5n-3, C22:5n-3, n-3/n-6, and C22:6n-3 levels significantly increased. In SO, flesh Σ n-9, Σ n-6, and 18:3n-6 levels significantly decreased, while flesh Σn-3 HUFA, C20:5n-3, C22:5n-3, and C22:6n-3 levels significantly increased.
3 Discussion
This study revealed that darkbarbel catfish fed diets containing SO for 80d had slightly higher growth rates than those fed FO (P>0.05) (Tab. 3). This result was similar to findings reported forsharpsnout sea bream (Diplodus puntazzo), brown trout (S. trutta L.), and brook char (Salvelinus fontinalis)[11,12,14]. The complete or partial replacement of FO with VO did not affect fish growth, possibly because there were no differences in FCR. Moreover, α-linolenic acid (C18:3n-3) was possibly desaturated into n-3 HUFA in the darkbarbel catfish P. vachelli, thereby meeting n-3 HUFA requirements for growth. In addition, previous studies have reported no significant effects of VO on the HSI of turbot (Psetta maxima) or rainbow trout (Oncorhynchus mykiss)[17,18]. Similarly, soybean oil did not significantly increase HSI in darkbarbel catfish P. vachelli. However, soybean oil-based diets significantly increased HSI, compared to FO-based diets in sharpsnout seabream (D. puntazzo)[14].
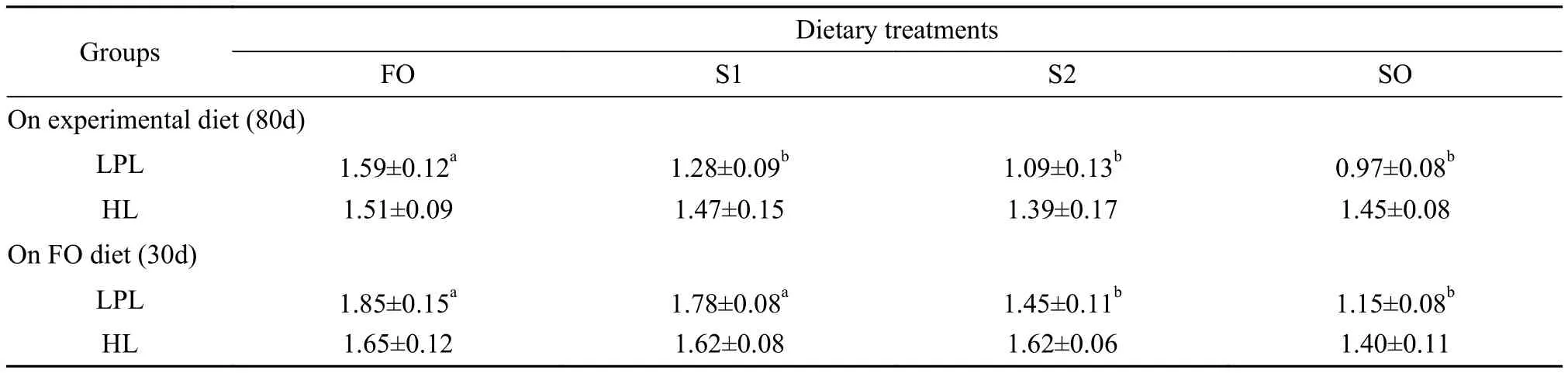
Tab. 5 LPL and HL activities at the end of the 80d period on the experimental diets and at the end of the 30d period on the fish oil diet (mean±SD; n=9)

Tab. 6 Flesh fatty acid composition at the end of growth period (GP) and restoration period (RP; % total fatty acids; mean±SD; n=9)
In this study, SO diets significantly reduced hepatic LPL activities with decreasing dietary levels of DHA, EPA, and linoleic acid (Tab. 5). Similar results have been reported by Michaud et al.[19], who observed that EPA decreased LPL mRNA levels, but linoleic acid increased LPL mRNA levels. However, Richard et al.[20]reported that the replacement of 60% FO with VO did not significantly affect hepatic lipogenesis or LPL activity in the liver and adipose tissues. Also, VO did not significantly influence hepatic HL activity. In red sea bream (P. major), HL gene expression levels were not affected during fasting or refeeding stages[21].
In this study, with increasing dietary SO, flesh Σ MUFA and C18:1n-9 levels increased (P<0.05), in accordance with the dietary fatty acid composition (Tab. 6). Similarly, the proportions of C18:1 n-9, C18:2 n-6, and C18:3 n-3 in Atlantic salmon (S. salar) flesh increased with increasing dietary rapeseed oil levels[22]. Meanwhile, C20:5n-3, C22:6n-3, Σ n-3 HUFA, and n-3/n-6 ratios in P. vachelli decreased with the addition of SO (P<0.05), correlating with the dietary levels. Similar results have been reported in marine species and fresh water species, such
[1]Pike I H. Eco-efficiency in aquaculture: global catch of wild fish used in aquaculture [J]. International Aquafeed, 2005, 8: 38—39
[2]FAO. The State of World Fisheries and Aquaculture. 2010, as turbot (P. maxima), gilthead sea bream (S. aurata), and sharpsnout seabream (D. puntazzo)[6,14,17]. In Atlantic salmon (S. salar), VO diets (33% of total oil) significantly reduced the flesh levels of C20:5n-3 and C22:6n-3 (by 70% and 75%, respectively), relative to the flesh levels in FO-fed fish[22]. Therefore, flesh fatty acid composition was representative of the dietary fatty acid profile. In addition, there was selective deposition of DHA in darkbarbel catfish P. vachelli, turbot (P. maxima), and Atlantic salmon (S. salar)[17,22]. Flesh C22:6n-3 levels were consistently higher than those in the experimental diets in P. vachelli (Table 6). Bell et al.[23]and Fr?yland et al.[24]reported that this selective deposition was attributable to the high specificity of fatty acyl-transferases for C22:6n-3 and to higher beta oxidation of C22:6n-3, compared to that of C20:5n-3. C20:5n-3 levels in European sea bass (D. labrax) flesh were lower than those in rapeseed, linseed, or soybean oil-containing diets[25]. This result was similar to that obtained in this study (Tab. 6).
After 30 d with a finishing FO diet, flesh Σn-3 HUFA, C20:5n-3, and C22:6n-3 levels in the S2 and SO groups significantly increased, but not to the extent found in fish fed FO for 110d (Tab. 6). Fountoulaki et al.[11]reported that feeding FO to gilthead sea bream (S. aurata L.) for 120d was not sufficient for the restoration of C20:5n-3 and C22:6n-3 levels. Similar conclusions were reached in European sea bass (D. labrax) fed 60% VO followed by FO for 5 months[25]. However, Izquierdo et al.[6]reported that flesh C22:6n-3 and C20:4n-6 levels in gilthead seabream (S. aurata) were restored after a 7-month feeding trial to levels of 60%—80% following a 3-month re-feeding period with FO. These results revealed that restoration of 20:5n-3 and 22:6n-3 in flesh with FO finishing diets may be different in different fish species; 30d FO finishing diets could significantly increase Σn-3 HUFA, C20:5n-3, and C22:6n-3 in P. vachelli fed with SO in the growth period.
In summary, the results of this study suggest that in the diets of darkbarbel catfish, soybean oil may substitute for almost 100% of the FO without affecting the growth rates. A re-feeding period of 30 d with 100% FO may significantly increase levels of Σn-3 HUFA, C20:5n-3 and C22:6n-3 in flesh, after feeding diets containing ≥50% SO.
63
[3]Tacon A G J. Use of fish meal and fish oil in aquaculture: a global perspective [J]. Aquatic Resources, Culture and Development, 2004, 1(1): 3—14
[4]Caballero M J, Obach A, Rosenlund G, et al. Impact of different dietary lipid sources on growth, lipid digestibility, tissue fatty acid composition and histology of rainbow trout, Oncorhynchus mykiss [J]. Aquaculture, 2002, 214(S1—4), 253—271
[5]Bell G, Torstensen B, Sargent J. Replacement of marine fish oils with vegetable oils in feeds for farmed salmon [J]. Lipid Technology, 2005, 17: 7—11
[6]Izquierdo M S, Montero D, Robaina L, et al. Alterations in fillet fatty acid profile and flash quality in gilthead seabream (Sparus aurata) fed vegetable oils for a long term period. Recovery of fatty acid profiles by fish oil feeding [J]. Aquaculture, 2005, 250(1—2): 431—444
[7]Mourente G, Bell J G. Partial replacement of dietary fish oil with blends of vegetable oils (rapeseed, linseed and palm oils) in diets for European sea bass (Dicentrarchus labrax L.) over a long term growth study: effects on flesh and liver fatty acid composition and effectiveness of a fish oil finishing diet [J]. Comparative Biochemistry Physiology, Part B, 2006, 145(4): 389—399
[8]Izquierdo M S, Obach A, Arantzamendi L, et al. Dietary lipid sources for seabream and seabass: growth performance, tissue composition and flesh quality [J]. Aquaculture Nutrition, 2003, 9(9): 397—407
[9]Francis D S, Turchini G M, Jones P L, et al. Growth performance, feed efficiency and fatty acid composition of juvenile Murray cod, Maccullochella peelii peelii, fed graded levels of canola and linseed oil [J]. Aquaculture Nutrition, 2007, 13(5): 335—350
[10]Fountoulaki E, Vasilaki A, Hurtado R, et al. Fish oil substitution by vegetable oils in commercial diets for gilthead sea bream (Sparus aurata L.); effects on growth performance, flesh quality and fillet fatty acid profile Recovery of fatty acid profiles by a fish oil finishing diet under fluctuating water temperatures [J]. Aquaculture, 2009, 289(S3—4): 317—326
[11]Turchini G M, Mentasti T, Froyland L, et al. Effects of alternative lipid sources on performance, tissue chemical composition, mitochondrial fatty acid oxidation capabilities and sensory characteristics in brown trout (Salmo trutta L) [J]. Aquaculture, 2003, 225(1—4): 251—267
[12]Guillou A, Soucy P, Khailil M, et al. Effects of dietary vegetable and marine lipid on the growth and organoleptic quality of flesh of brook charr (Salvelinus fontinalis) [J]. Aquaculture, 1995, 136(3): 351—362
[13]Mourente G, Good J E, Bell J G. Partial substitution of fish oil with rapeseed, linseed and olive oil for European sea bass (Dicentrarchus labrax L.): effects on flesh fatty acid composition, plasma prostaglandin E2 and F2, immune functionsand effectiveness of fish oil finishing diet [J]. Aquaculture Nutrition, 2005, 11(1): 25—40
[14]Piedecausa M A, Mazón M J, García B, et al. Effects of total replacement of fish oil by vegetable oils in the diets of sharpsnout seabream (Diplodus puntazzo) [J]. Aquaculture, 2007, 263(S1—4): 211—219
[15]Panserat S, Hortopan G A, Plagnes-Juan E, et al. Differential gene expression after total replacement of dietary fish meal and fish oil by plant products in rainbow trout (Oncorhynchus mykiss) liver [J]. Aquaculture, 2009, 294(1—2): 123—131
[16]Folch J M, Lees M, Sloane-Stanley G H. A simple method for the isolation and purification of total lipides from animal tissues [J]. Journal of Biological Chemistry, 1957, 226(1): 497—509
[17]Regost C, Arzel J, Robin J, et al. Total replacement of fish oil by soybean or linseed oil with a return to fish oil in turbot (Psetta maxima)-1. Growth performance, flesh fatty acid profile, and lipid metabolism [J]. Aquaculture, 2003, 217(1—4): 465—482
[18]Panserat S, Hortopan G A, Plagnes-Juan E, et al. Differential gene expression after total replacement of dietary fish meal and fish oil by plant products in rainbow trout (Oncorhynchus mykiss) liver [J]. Aquaculture, 2009, 294(S1—2): 123—131
[19]Michaud S E, Renier G. Direct regulatory effect of fatty acids on macrophage lipoprotein lipase: potential role of PPARs [J]. Diabetes, 2001, 50(3): 660—666
[20]Richard N, Mourente G, Kaushik S, et al. Replacement of a large portion of fish oil by vegetable oils does not affect lipogenesis, lipid transport and tissue lipid uptake in European seabass (Dicentrarchus labrax L.) [J]. Aquaculture, 2006, 261(3): 1077—1087
[21]Liang X F, Oku H, Ogata H Y. The effects of feeding condition and dietary lipid level on lipoprotein lipase gene expression in liver and visceral adipose tissue of red sea bream Pagrus major [J]. Comparative Biochemistry Physiology, Part A, 2002, 131(2): 335—342
[22]Bell J G, Tocher D R, Henderson R J, et al. Altered fatty acid compositions in Atlantic salmon (Salmo salar) fed diets containing linseed and rapeseed oils can be partially restored by a subsequent fish oil finishing diet [J]. The Journal of Nutrition, 2003, 133(9): 2793—2801
[23]Bell J, Mcevoy J D, Mcghee F, et al. Replacement of fish oil with rapeseed oil in diets of Atlantic salmon (Salmo salar) affects tissue lipid compositions and hepatocyte fatty acid metabolism [J]. Journal of Nutrition, 2001, 131(5): 1535—1543
[24]Fr?yland L, Madsen L, Eckhoff K M, et al. Carnitine palmitoyltransferase I, carnitine palmitoyltransferase Ⅱ, and acyl-CoA oxidase activities in Atlantic salmon (Salmo salar) [J]. Lipids, 1998, 33(9): 923—930
[25]Montero D, Robaina M J, Caballero R, et al. Growth, feed utilization and flesh quality of European sea bass (Dicentrarchus labrax) fed diets containing vegetable oils: a time-course study on the effect of a re-feeding period with a 100% fish oil diet [J]. Aquaculture, 2005, 248(S1—4): 121—134
再投喂魚油對瓦氏黃顙魚肌肉脂肪酸組成的影響
邵 婷1,2覃川杰1袁登越1文正勇1李華濤1
(1. 內江師范學院生命科學學院, 長江上游魚類資源保護與利用四川省重點實驗室, 內江 641100; 2. 四川師范大學生命科學學院, 成都 610101)
為研究植物油替代魚油對瓦氏黃顙魚(Pelteobagrus vachelli)生長及肌肉脂肪組成的影響及重投喂魚油對瓦氏黃顙魚肌肉脂肪酸組成的影響, 實驗以大豆油分別替代飼料中的0(FO)、50 (S1)、75 (S2)和100% (SO)的魚油配制等氮、等能的顆粒飼料, 每組設置3個平行, 養殖80d后, 再投喂魚油30d。結果表明, 飼料中添加豆油不會顯著影響瓦氏黃顙魚的增重率、肝體指數和體成分(P>0.05)。隨著飼料中大豆油含量的增加, S2和SO組肌肉中C18:1n-9、C18:2n-6和單不飽和脂肪酸比例顯著增加(P<0.05), 而C20:5n-3, C22:5n-3及n-3/n-6比例顯著下降(P<0.05)。再投喂魚油30d后, SO組肌肉中C18:3n-6、C20:4n-6、Σ n-9、Σ n-6和S2組中C18:1n-9、Σ n-6比例顯著下降(P<0.05), 而S2和SO組肌肉中Σn-3多不飽和脂肪酸、C20:5n-3和C22:5n-3比例顯著增加(P<0.05)。在生產中, 可采用先植物油飼料、后魚油飼料的養殖方式提高瓦氏黃顙魚肌肉品質(增加有益人類健康的多不飽和脂肪酸)。
瓦氏黃顙魚; 魚油; 豆油; n-3多不飽和脂肪酸; 脂肪酸修復
Qin Chuan-Jie, E-mail: qinchuanjie@126.comture, the substitution of FO with VOs may affect growth and disease resistance, and decrease flesh fatty acid composition.
10.7541/2017.18
