基于植物區系的雷州半島天然林群落樹種特征研究
2017-03-09 08:28:37韓維棟黃劍堅
生態學報 2017年24期
關鍵詞:物種
韓維棟,黃劍堅
1 廣東海洋大學農學院, 湛江 524088 2 廣東海洋大學寸金學院, 湛江 524094
植物區系特征研究及群落優勢種重要值評價研究,是傳統群落生態學的重要研究內容。傳統涉及植物區系和重要值的一起研究的文獻,習慣于把植物區系和重要值分割開來進行研究[1-6]。因此,絕大部分的文獻一般從該區系具有多少物種、屬、科等組成開展分析[7- 12],而忽略了該區系植物物種或屬科所在群落中的地位(即重要值)。未能充分反映出該區域群落的某個區系植物物種數量占優勢,而實際上可能由于該區系植物物種的重要值偏低導致區系植物物種整體上在群落內處于次要地位的問題。鑒于此,本文擬提出SCI指數(SCI=species community index),即物種群落指數。該指數不僅可以真實客觀了解和反映一個地區植物區系的種類組成、分布區類型以及發生、發展等重要特征,并同時了解和反映該區系屬、或科或樹種在群落中的地位。因此,相比之前的研究思路和研究方法,則更有優勢,對于反映植物區系的物種參與群落構建,可為樹種的選擇以及進一步分析該區系樹種在群落中的競爭力,有著重要的意義。
我國熱帶北緣地區具有或曾經擁有發育良好的地帶性植被熱帶季雨林,是雷州半島地區植被區系構成的重要組成部分。由于長期人為開墾活動,廣東西南部雷州半島地區原生頂極植物群落即過渡性熱帶季雨林已經消失殆盡,小片次生季雨林也十分珍稀罕見[13]。本文以雷州半島尚存的次生季雨林木本植物群落為研究對象,開展物種群落指數研究,可更好認識其木本植物群落的組成、現狀和發展趨勢,開發利用好其天然林木資源,對雷州半島生態保護與恢復具有舉足輕重的意義,為投入近300億的雷州半島熱帶季雨林的生態恢復工程提供基礎參考。
1 研究地概況
雷州半島(109°30′—110°55′E, 20°12′—21°35′N)位于中國大陸的最南端,地勢比較平坦,由北向南呈北高-中低-南高的龜背地形,面積12470 km2,為我國第三大半島。雷州半島陸地為玄武巖、花崗巖或砂頁巖酸性磚紅土壤或赤紅壤、水稻土,沿海為濱海鹽土、砂土和紅樹林酸性硫酸土等;北熱帶海洋性特色明顯,季風明顯;從4月上旬至11月上旬為夏季,11月中旬至4月上旬為秋季和春季;熱量資源豐富,歷年年均氣溫22.8—23.5℃,低壓、熱帶風暴、臺風登陸影響頻繁。
2 研究方法與調查樣地
2.1 調查方法與樣地
本文選擇僅存的雷州半島天然植被最大的8個區域開展調查。鷹峰嶺位于雷州市英利鎮東北側,20°34′52.76″N,110°10′35.35″E,海撥247 m,是雷州半島第二高峰,植被類型(下同)為次生季雨林;謝鞋山位于廉江市廉城鎮東南5 km的謝鞋村,21°35′58.55″N,110°20′33.82″E,海撥42 m;湖光巖位于湛江市西南部, 21°08′24.26″N,110°17′11.18″E,海撥16 m;云腳村位于湛江市麻章區湖光鎮, 21°07′25.51″N,110°14′23.59″E,海撥6 m,森林面積73 hm2;高橋天然林位于湛江市廉江市西部, 21°36′26.79″N,109°48′23.92″E,海撥4 m;根竹嶂位于廉江市和寮鎮西北部,21°51′50.35″N,110°07′52.95″E,海拔210 m;足榮村位于雷州市半島西南部, 20°37′10.28″N,109°56′20.27″E,海撥82 m,有保護較好的20多hm2風水林,是該雷州少有的小片原始森林;雙洋村位于雷州半島徐聞縣的下橋鎮,20°31′4.71″N,110°08′4.88″E,海撥121 m,有保護較好的13多hm2風水林,是徐聞少有的小片原始森林。
在每個區域設置10個20 m×20 m標準樣地。對樣地中胸徑大于5 cm的樹木進行每木檢測,記錄樹種的種類、數量、胸徑、樹高、冠幅等。
2.2 樣地重要值和物種多樣性指數的計算方法
喬木與灌木植物重要值合并計算,重要值求取按照每個樣地標準地分別統計后的平均。重要值、相對密度、相對蓋度、相對頻度等指標計算參考文獻[13],按各樹種重要值判斷該樹種在群落中的位置。
2.3 物種群落指數的建立
物種群落指數,用以描述區系成分參與構建森林群落的特征值,以植物群落的喬木和灌木樹種為調查群落對象,統計樹種數、重要值、區系分布。按照文獻依據吳征鎰《中國種子植物屬的分布區類型》屬的地理分布區類型及其種類組成[14],屬種的分布區類型及其變型為15個分布區類型。其它區系暫不列入該指數的統計范圍。物種群落指數實際為某個植物群落每個區系的科或屬的所有物種的重要值組成所有,各個部分之間通過解析分段函數整合在一起。物種群落指數定義域是各個區系的種屬的并集,值域也是15個區系成分樹種參與構建森林群落或依賴森林群落作為棲息地的特征值(即重要值)。因此,可得物種群落指數的數學公式為:
其中,f(x)值域,總和為1。其中,x1a為1區世界分布的某個樹種的重要值;x2b為2區泛熱帶分布的某個樹種的重要值;以此類推,X15o為15區中國、海南特有分布的某個樹種的重要值。雷州半島區域的所有天然林樹種的重要值按照大小開展排序。數據主要采用SPSS 20.0和Excel 2007開展數據處理。
3 結果與分析
3.1 雷州半島8個區域樣地物種群落指數分析
雷州半島8個不同區域樣地物種群落指數組成各不同,具有一定特征(表1)。湛江云腳村物種群落指數以2區系泛熱帶分布、4區系古熱帶分布、5區系熱帶亞洲至熱帶大洋洲分布的樹種重要值為主導。湛江湖光巖和雷州鷹峰嶺以2區系樹種重要值占絕對主導。雷州足榮村、徐聞雙洋村、廉江高橋和廉江謝鞋山物種群落指數皆以2區系、4區系、5區系和7區熱帶亞洲分布的樹種重要值為主導,但四個樣地各有所偏重,雷州足榮村偏重于5區系,徐聞雙洋村和廉江高橋偏重于4區系,廉江謝鞋山偏重于7區系和4區系。廉江根竹嶂由于地理位置較為特殊,為山脈的主峰,受冷空氣影響,以9區系東亞和北美洲間斷分布為主,為典型的過渡區系特征,同時帶有熱帶性區系特征。
3.2 雷州半島同一區系不同樣地對比分析
3.2.1第2區系的不同樣地對比
第2區系的不同樣地物種群落指數排序可得:湛江湖光巖>雷州鷹峰嶺>廉江謝鞋山>徐聞雙洋村>湛江云腳村>雷州足榮村>廉江高橋>廉江根竹嶂(表2)。湛江湖光巖和雷州鷹峰嶺遭受過生態破壞由于歷史原因在20世紀60年代遭到大量砍伐,造成大量的天然樹種消失[15]。兩地目前以兩區系的物種為主導的生態恢復階段。其中,鵝掌柴、樸樹、紫金牛、假蘋婆、山杜英、樟葉樸、布渣葉是該區最重要的鄉土樹種。
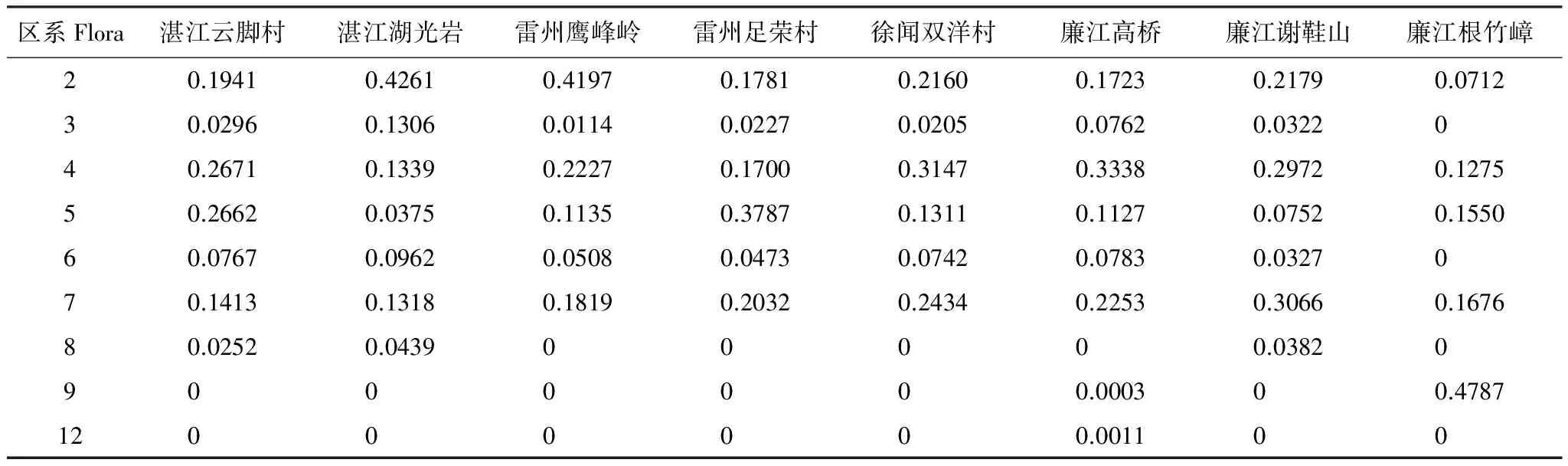
表1 不同樣地物種群落指數
3.2.2 第3區系的不同樣地對比
第3區系不同樣地物種群落指數排序可得:湛江湖光巖>廉江高橋>廉江謝鞋山>湛江云腳村>雷州足榮村>徐聞雙洋村>雷州鷹峰嶺(表3)。廉江根竹嶂樣地不存在該區系樹種。從指數分析可見該區系物種數量少,在雷州半島群落中處于劣勢地位。其中,潺槁樹是最為常見的樹種。
3.2.3 第4區系的不同樣地對比
8個樣地對比可得:廉江高橋>徐聞雙洋村>廉江謝鞋山>湛江云腳村>雷州鷹峰嶺>雷州足榮村>湛江湖光巖>廉江根竹嶂(表4)。4區系的樹種在雷州半島群落中普遍占據較為重要的地位,最高值達0.3339。可見,4區系樹種在雷州半島樣地群落群落中作為重要的組成部分。其中,竹節樹、粗糠柴、野生橄欖、楹樹等為群落優勢種;烏墨、倒吊筆、見血封喉、苦楝等喬木樹種,以及土壇樹、暗羅等灌木或小喬木在群落中處于次要地位。
3.2.4 第5區系的不同樣地對比
第5區系8個樣地對比可得:雷州足榮村>湛江云腳村>廉江根竹嶂>徐聞雙洋村>雷州鷹峰嶺>廉江高橋>廉江謝鞋山>湛江湖光巖(表5)。第5區系樹種在不同的地理環境和演替時期存在差異,雷州足榮村和湛江云腳村等保存較好的樣地群落占據主導地位,而在雷州鷹峰嶺、廉江謝鞋山、湛江湖光巖等生態恢復區處于劣勢。其中,陰香、樟樹、海紅豆等為優勢喬木樹種。
3.2.5第6區系的不同樣地對比
第6區系不同樣對比可得:湛江湖光巖>廉江高橋>湛江云腳村>徐聞雙洋村>雷州鷹峰嶺>雷州足榮村>廉江謝鞋山(表6)。該區系與第3區系的分布特點類似,樹種數量少,在雷州半島群落處于劣勢地位最高值僅為0.0962。基本上作為群落構建的次要組成。
3.2.6第7區系的不同樣地對比
第7區系不同樣對比可得:廉江謝鞋山>徐聞雙洋村>廉江高橋>雷州足榮村>雷州鷹峰嶺>廉江根竹嶂>湛江云腳村>湛江湖光巖(表7)。該區系樹種在雷州半島地區群落內分布較為均衡。其中,野生野生荔枝、銀柴、鵲腎樹、桂木、山楝、幌傘楓等為優勢種,在群落更新中占據較為重要的地位。
3.2.7其他區系的不同樣地對比
8區北溫帶分布、9區東亞和北美洲間斷分布,12區地中海區、西亞至中亞分布及其變型等3個區系在雷州半島地區的物種多樣性低,為零星分布,在雷州半島群落中普遍處于劣勢地位,廉江根竹嶂除外(表8)。主要因為廉江根竹嶂是云開大山支脈,受亞熱帶海洋季風氣候影響,為典型的熱帶亞熱帶原始森林植被,以8區北溫帶分布的米櫧樹種占據主導地位。
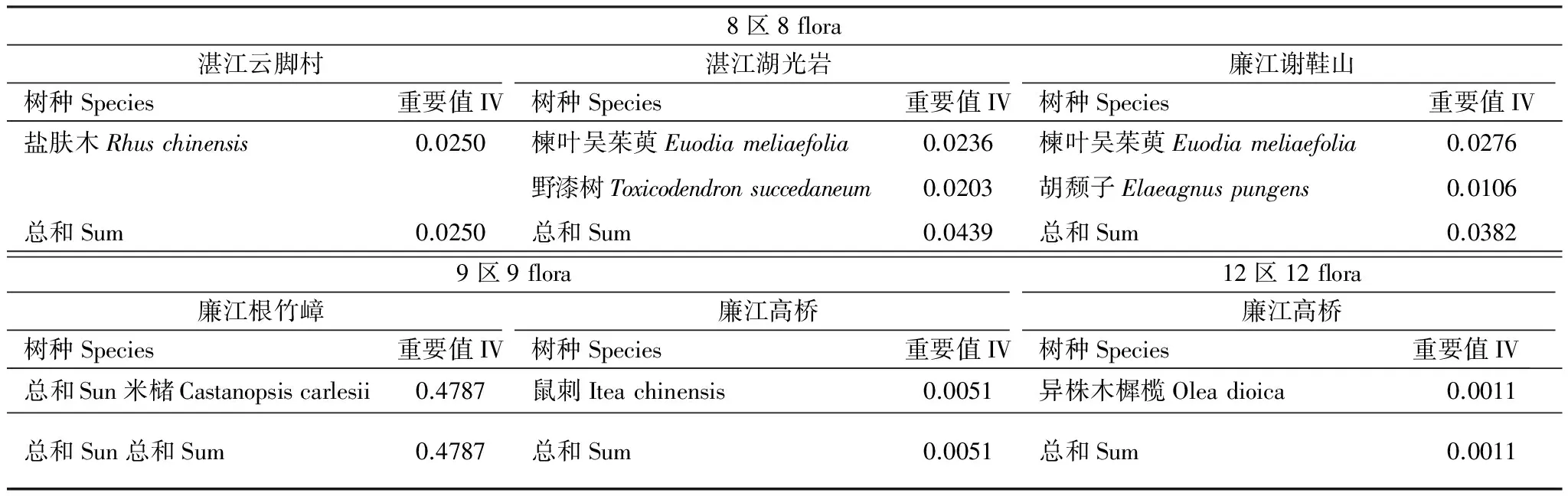
表8 其它區系不同樣地區系重要值分析
4 討論
4.1 雷州半島8個區域樣地物種群落指數的討論
雷州半島不同區域樣地群落特征各不相同。本文研究結果表明雷州半島地區,除了海拔210 m立地條件較為特殊的廉江根竹嶂,其他樣地均以熱帶性區系成分為主,系具明顯熱帶性質。該結果與韓維棟等研究結果一致[16],是合理和正確的。本文所提出來的物種群落指數,有效分析與揭示雷州半島8個重要的天然植被區域樣地森林區系樹種組成的特征,何種區系樹種在群落中占據主導地位。研究一個樣地或者一個區域的物種群落指數,需要結合當地的立地條件、人為干擾、種間競爭、種內競爭以及區系植物競爭等多種內外界影響因素加以深入分析,探尋之間的關系,哪種影響因素占主導,如何影響區系樹種在群落中的分布等。
4.2 同一區系不同區域樣地對比分析結果討論
第2區系樹種在雷州半島的群落構建中存在著生態環境越好,所占地位越低的趨勢。因此,該區系樹種對生態環境恢復有著重要影響,在生態恢復的群落演替前期占據主導地位。韓維棟等[15]研究結果表明,臺風是導致湛江湖光巖地區森林群落發生演替的一個重要因素。該區系占主導地位,可能與臺風干擾有關。
第3區系樹種在所有的樣地中與其它區系樹種競爭時均不占優勢,甚至在廉江根竹嶂樣地不存在該區系樹種。那么,究竟是什么影響因素導致該區系樹種在雷州半島地區群落中作為次要的存在,甚至并不存在呢,還需要進一步深入研究和分析。第6區系樹種和第3區系分布類似,總體上在群落中不占優勢。那么究竟是立地條件,還是種群競爭等何種因素導致兩個區系的物種在雷州半島地區占次要位置,還需要進一步深入分析。
第4區系在雷州半島分布較為側重,所有樣地均出現,在雷州鷹峰嶺、徐聞雙洋村、廉江高橋等樣地以及不同演替時期可占據重要地位。由于該區以熱帶雨林、季雨林和稀樹草原為主,終年炎熱多雨,季節變化不明顯,植物全年生長,且氣候條件在很長的地質時期內無多大變化,導致植物區系種類十分豐富,達4500種以上。該區系的樹種普遍適應雷州半島地區,并呈現良好的適應性,可能與其土壤適應性強很有直接相關的關系。第7區系和第4區系分布類似,在雷州半島地區廣泛均衡分布,并在個別樣地占據主導地位。
第5區系樹種主要存在雷州半島群落演替中后期,并在群落中占據主導的地位。同時,雷州足榮村和徐聞雙洋村,兩個樣地距離50 km以內,氣候條件差異不大,而海拔相差40 m。可能立地條件是導致兩個樣地該區系樹種差別較大原因。
8區系、9區系和12區系由于不適宜當地的環境條件,在群落中處于低下的地位。那么,屬于8區北溫帶分布的米櫧樹種重要值高達0.4787,作為優勢種存在,極為罕見,但究竟是何影響因素導致,需深入分析。
5 結論
雷州半島地區樣地熱帶季雨林群落的樹種組成與構建以熱帶性區系成分為主,具明顯熱帶性質。第2區系重要值存在著生態環境越好,該區系所占群落比例越低的趨勢;第3區系樹種與其它區系樹種競爭時均不占優勢,屬于次要的存在;第4區系分布較為側重,在不同演替時期可占據重要地位;第5區系主要存在雷州半島群落演替中后期,并在群落中占據主導地位。第6區系樹種和第3區系分布類似。第7區系和第4區系分布類似。8區系、9區系和12區系群落樹種指數為次要存在。因此,物種群落指數能夠較好的體現雷州半島樣地各個區系樹種在植物群落組成的特征。
[1] 李海靜. 江西大崗山常綠闊葉林植物區系及多樣性研究[D]. 北京: 北京林業大學, 2005.
[2] 龍海建, 洪劍明, 胡東, 徐慶安. 拒馬河北京段濕地植物群落的種類組成及區系分析. 首都師范大學學報: 自然科學版, 2011, 32(1): 49- 53.
[3] 侯昭強, 文光玉, 周丹, 李敏敏, 杜凡. 云南榧樹群落及其植物區系特征研究. 西部林業科學, 2015, 44(3): 37- 44.
[4] 王立龍, 陸林, 戴建生. 太平湖國家濕地公園生態保育區草本植物區系及其在不同干擾下的多樣性動態. 自然資源學報, 2010, 25(8): 1306- 1319.
[5] 朱華. 西雙版納龍腦香林與熱帶亞洲和中國熱帶北緣地區植物區系的關系. 云南植物研究, 1994, 16(2): 97- 106.
[6] 江明喜, 吳金清, 葛繼穩. 神農架南坡送子園珍稀植物群落的區系及生態特征研究. 武漢植物學研究, 2000, 18(5): 368- 374.
[7] 修晨, 歐陽志云, 鄭華. 北京永定河—海河干流河岸帶植物的區系分析. 生態學報, 2014, 34(6): 1535- 1547.
[8] 郝云慶, 江洪, 余樹全, 金靜, 馬元丹. 桫欏植物群落區系進化保守性. 生態學報, 2009, 29(8): 4102- 4111.
[9] 董東平, 鄭敬剛, 葉永忠. 河南嵩山國家森林公園木本植物區系. 林業科學, 2009, 45(3): 160- 166.
[10] 李家湘, 王旭, 黃世能, 周光益, 鄒濱, 羅鑫華. 南嶺中段冰災受損群落和植物區系特征及保護生物學意義. 林業科學, 2010, 46(3): 166- 172.
[11] 王雷, 張道遠, 黃振英, 田長彥. 新疆鹽生植物區系分析. 林業科學, 2008, 44(7): 36- 42.
[12] 王娟, 馬欽彥, 杜凡, 楊宇明. 云南大圍山種子植物區系海拔梯度格局分析. 植物生態學報, 2005, 29(6): 894- 900.
[13] 韓維棟, 高秀梅, 黃月瓊, 劉新田, 李林峰. 廉江次生季雨林群落類型與多樣性研究. 林業科技, 2001, 26(4): 1- 4.
[14] 吳征鎰. 中國種子植物屬的分布區類型. 云南植物研究, 1991, 13(S4): 1- 139.
[15] 韓維棟, 黃劍堅, 劉素青. 強臺風干擾后林分空間結構變化的研究. 中南林業科技大學學報, 2013, 33(7): 8- 13.
[16] 韓維棟, 陳杰, 高秀梅. 雷州半島木本植物區系及其驅動力分析. 生態科學, 2014, 33(6): 1091- 1094.
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
